Torfmooskultivierungsflächen als Lebensraum für Vögel, Amphibien, Libellen und Tagfalter
Abstract
Auf Hochmoore spezialisierte Arten sind aufgrund der großflächigen entwässerungsbasierten Nutzung und Umwandlung ihrer Lebensräume heute häufig stark gefährdet und in ihren Beständen isoliert. Aktuelle klimapolitische Ziele führen zu neuen Ansätzen für eine nasse, nachhaltige Nutzung von Torfabbauflächen oder landwirtschaftlich genutzten Hochmoorstandorten, der sogenannten Paludikultur. Auf Hochmoorböden eignet sich die Kultivierung von Torfmoosen (Sphagnum spec.), die aufgrund der nassen Bedingungen und der Etablierung von hochmoortypischer Vegetation potenzielle Lebensräume für Tierarten der Hochmoore schafft. Im Rahmen dieser Studie wurde untersucht, inwieweit diese Potenziale von Vögeln, Amphibien, Libellen und Tagfaltern genutzt werden können. In den Jahren 2017 und 2018 erfolgten dazu auf zwei Torfmooskultivierungsflächen (Einrichtung 20152016), zwei Wiedervernässungsflächen nach Torfabbau und zwei naturnahen Hochmoorflächen im Emsland (Niedersachsen) vergleichende Erfassungen. Dabei hat sich insbesondere ein hohes Potenzial der Kultivierungsflächen für hochmoortypische und andere Offenlandarten der Brutvögel sowie für hochmoortypische Libellenarten gezeigt. Für den Moorfrosch, als einzige moortypische Amphibienart, können Torfmooskultivierungsflächen als wertvolle Biotopverbundelemente dienen. Dagegen war die Habitatqualität für Tagfalter durch die Bewirtschaftung der Flächen sowie fehlende Windschutzstrukturen gering.
Sphagnum paludiculture as a habitat for birds, amphibians, Odonata, and Lepidoptera What habitat quality do sustainably managed raised bogs provide?
Peatland species are often highly endangered and isolated in their populations due to large-scale drainage-based use and destruction of their habitats. Current climate policy goals are leading to new approaches for a wet, sustainable use of peatland sites, so-called paludiculture. On raised bog soils, the cultivation of peat mosses (Sphagnum farming) is suitable, which creates potential habitats for peatland species due to the wet conditions and the establishment of vegetation typical for raised bogs. This study investigated the extent to which this potential can be used by birds, amphibians, dragonflies, damselflies, butterflies and moths . In 2017 and 2018, comparative surveys were carried out at two Sphagnum farming sites (established from 2015 to 2016), two rewetted cut-over restoration sites, and two near-natural bogs in Lower Saxony (Germany). In particular, the Sphagnum farming sites showed high potential for characteristic peatland bird species and lowland breeders, as well as for characteristic Odonata species. For the moor frog, the only peatland amphibian species in this study, Sphagnum farming sites could serve as valuable connecting elements in habitat corridors. In contrast, the habitat quality for Lepidoptera was low due to site management and the lack of windbreak structures.
- Veröffentlicht am

Eingereicht am 22. 03. 2022, angenommen am 10. 09. 2022
1 Einleitung
Auf Hochmoore spezialisierte Tier- und Pflanzenarten haben insbesondere im 19. und 20. Jahrhundert in Mitteleuropa einen drastischen qualitativen und quantitativen Lebensraumverlust erfahren, sodass ein zunehmender Rückgang dieser Arten in ihrer Verbreitung und relativen Häufigkeit zu verzeichnen ist (Hughes et al. 2008, Röser 1990, Succow & Joosten 2001, van Grunsven et al. 2020). Insbesondere die großflächige Entwässerung und Umwandlung für die land- und forstwirtschaftliche Kultivierung und der industrielle Torfabbau haben dazu geführt, dass mehr als 95 % der Moorflächen stark degradiert oder zerstört sind (Joosten et al. 2017, Succow & Joosten 2001). Naturnahe, wachsende (Torf akkumulierende) Hochmoore mit intakten Vegetationsstrukturen, die den Lebensraumansprüchen von spezialisierten Arten entsprechen, gibt es heute nur noch in sehr geringem Flächenumfang (Joosten & Couwenberg 2001). Bei der Hochmoorfauna handelt es sich um hochspezialisierte Arten, die sich an die extremen Standortbedingungen der Hochmoore (vor allem Wasser- und Nährstoffhaushalt, Temperaturschwankungen) angepasst haben (Röser 1990, Spitzer & Danks 2006). Verändern sich diese Standortbedingungen, verlieren die spezialisierten Hochmoorarten nicht nur ihren Lebensraum, sondern auch ihren Konkurrenzvorteil gegenüber weniger spezialisierten und deshalb konkurrenzstärkeren Arten (Elo et al. 2015, Röser 1990, van Kleef et al. 2012). Die Isolation der noch intakten Hochmoor-Restflächen führt zudem dazu, dass hochmoortypische Arten in ihrer Ausbreitung eingeschränkt und deshalb die Populationen geschwächt werden (Verberk et al. 2006). Es ist davon auszugehen, dass die bestehenden Gefährdungen der Hochmoorarten durch die Folgen des Klimawandels noch zusätzlich verstärkt werden. Insbesondere fehlende Niederschläge in den Sommermonaten, aber auch ganzjährig höhere Temperaturen, führen langfristig zu Verschlechterungen der Habitatqualität etwa durch Austrocknen von Gewässerstrukturen oder Veränderungen in der Vegetationsstruktur und -zusammensetzung (Behrens et al. 2009, Ott et al. 2015, Sperle & Bruelheide 2021, Thomas et al. 2004, Weiß et al. 2011).
Durch die Wiedervernässung von Moorflächen werden seit den 1980er-Jahren neue Ersatzlebensräume für viele Arten der Hochmoore geschaffen (Baumann et al. 2021 b, Blüml & Sandkühler 2015). Dies beschränkt sich jedoch bisher vor allem auf ehemalige Torfabbauflächen (NMU 2016, Schmatzler 2015). Mehr als drei Viertel der Moorböden befinden sich weiterhin in land- und forstwirtschaftlicher Nutzung (UBA 2020). Die von der Klimarahmenkonvention der Vereinten Nationen gesetzten Klimaschutzziele, die Treibhausgasemissionen zu senken, um die globale Erwärmung auf möglichst 1,5 °C zu begrenzen (UNFCCC 2015), bringen auch im Landnutzungssektor beim Umgang mit Moorstandorten einen erheblichen Handlungsbedarf bis 2030 beziehungsweise 2050 mit sich (BMU 2016, 2021, Roe et al. 2019, Tanneberger et al. 2021). Um die hohen Treibhausgasemissionen aus der entwässerungsbasierten Nutzung von Moorböden zu reduzieren, müssen die Wasserstände auf diesen Flächen angehoben werden (Joosten et al. 2016, Tanneberger et al. 2021, Tiemeyer et al. 2020). Dabei sind ein vollständiger Torferhalt und eine Bindung von Kohlenstoff nur bei naturnahen oberflächennahen Wasserständen möglich (Wilson et al. 2016), die eine herkömmliche landwirtschaftliche Nutzung, insbesondere den Einsatz von schweren Maschinen, nicht zulassen und an nasse Standortbedingungen angepasste Nutzpflanzen erfordern. Deshalb werden Paludikulturen als nasse und somit klimaschonende Bewirtschaftungsformen von landwirtschaftlich vorgenutzten Moorstandorten oder ehemaligen Torfabbauflächen getestet (Wichtmann et al. 2016). Während auf Niedermoorstandorten verschiedene Kulturen möglich sind, eignet sich auf den nährstoffarmen Hochmoorböden die Kultivierung von Torfmoosen (Sphagnum spec.) (Gaudig et al. 2018). Pflanzenbauliche Versuche zeigen, dass die Torfmoosbiomasse als Substratausgangsstoff für den Erwerbsgartenbau geeignet ist und aufgrund ähnlicher physikalischer und chemischer Eigenschaften eine nachhaltige Alternative für die Verwendung von Weißtorf darstellt (Emmel 2008, Gaudig et al. 2018, Grantzau & Gaudig 2005). Aber auch für die nutzungsfreie Renaturierung von ehemaligen Abtorfungsflächen wird zunehmend die Bedeutung der Beimpfung mit Torfmoosen erkannt, um über die Einbringung von Torfmoosmaterial die Wiederansiedlung der typischen Hochmoorvegetation in Renaturierungsprojekten zu ermöglichen und zu beschleunigen (Hölzel et al. 2019). In den kommenden Jahren ist deshalb nicht nur im Gartenbau, sondern auch in Naturschutzprojekten mit einer erhöhten Nachfrage nach Torfmoos-Spendermaterial und einem Bedarf an Torfmoosvermehrungsflächen in Naturschutzprojekten zu rechnen.
Durch das Wassermanagement und die Etablierung von Torfmoosen werden auf Torfmooskultivierungsflächen künstliche Nutzungssysteme geschaffen, die potenzielle Ersatzhabitate für Arten der Hochmoore darstellen. Inwieweit diese Systeme tatsächlich als Lebensraum genutzt werden, ist jedoch noch weitgehend unbekannt, da es weltweit bisher nur wenige Pilotflächen zur Torfmooskultivierung gibt. Diese Pilotflächen wurden vor allem in Deutschland und Kanada angelegt (Gaudig et al. 2018, Grobe et al. 2021, Holthuis & Hofer 2021, Pouliot et al. 2015). Ein Nachweis der Nutzung durch hochmoortypische Arten erfolgte bisher nur für wenige ausgewählte Artengruppen wie Spinnen und Libellen (Muster et al. 2015, Muster et al. 2020, Zoch & Reich 2020).
Ziel dieser Studie war es deshalb, das Potenzial von Torfmooskultivierungsflächen als Lebensraum für Vögel, Amphibien, Libellen und Tagfalter abzuschätzen. Dazu wurden im Emsland (Niedersachsen) die Artenvorkommen auf zwei Torfmooskultivierungsflächen mit zwei Wiedervernässungsflächen nach Torfabbau und zwei naturnahen Hochmoorflächen verglichen.
2 Untersuchungsgebiete und Methoden
2.1 Untersuchungsgebiete
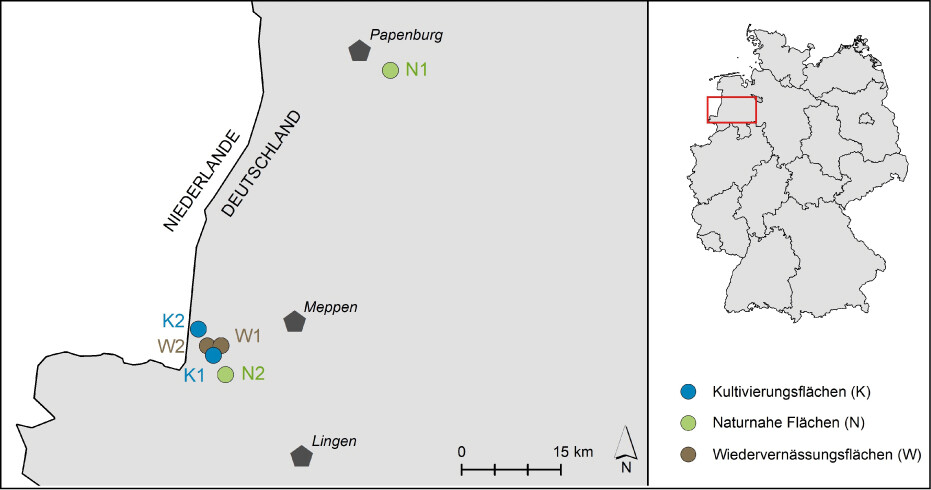
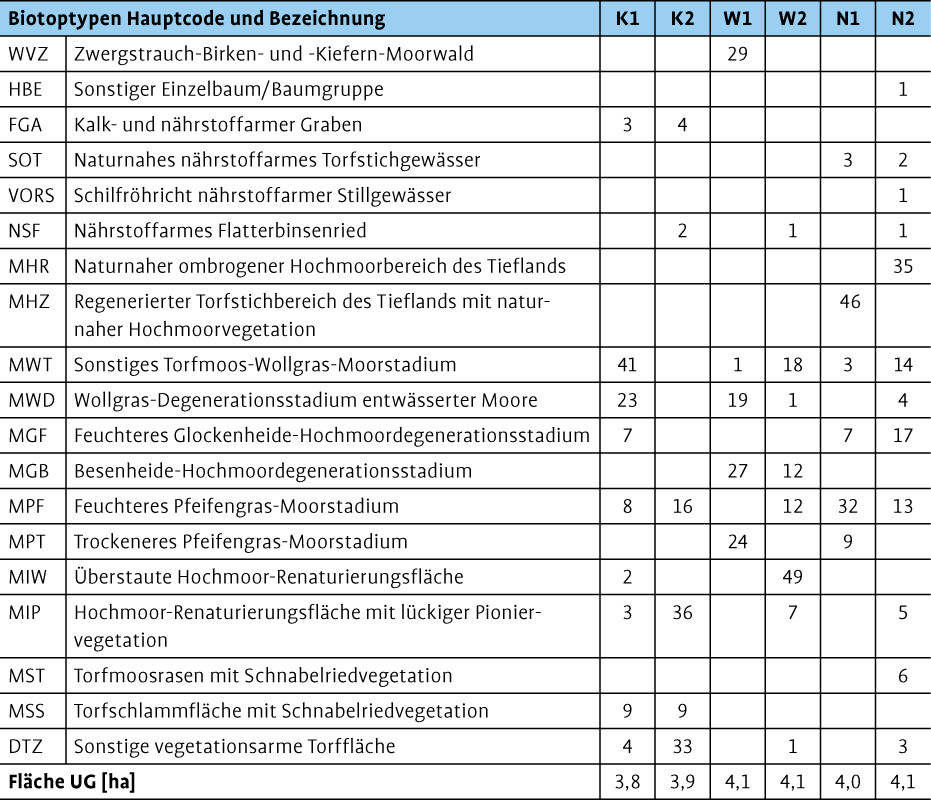
Die Untersuchungsgebiete (UG) liegen in Niedersachsen (Abb. 1) im Landkreis Emsland in der naturräumlichen Region Ostfriesisch-Oldenburgische Geest. Die Landschaft ist stark von Landwirtschaft, wenigen Waldflächen und ausgedehnten, heute überwiegend kultivierten oder in Abtorfung befindlichen Mooren geprägt (Drachenfels 2010). Es wurden zwei Torfmooskultivierungsflächen (K), zwei Wiedervernässungsflächen nach Torfabbau (W) und zwei naturnahe Hochmoorflächen (N) untersucht. In jedem UG wurde eine Fläche von rund 4 ha für die Erfassungen abgegrenzt.
Torfmooskultivierungsflächen
Die zwei Torfmooskultivierungsflächen wurden nach Beendigung des Torfabbaus (Frästorfverfahren) auf stark zersetztem Schwarztorf angelegt, für Details zur Anlage siehe auch Graf et al. (2017) und Grobe et al. (2021). Bei der Einrichtung der Flächen wurden neben den reinen Torfmoosflächen (jeweils rund 2,5 ha) auch Fahrdämme und Bewässerungsgräben gebaut, die ebenfalls bei der Untersuchung mit erfasst wurden (jeweils rund 4 ha).
Die Kultivierungsfläche K1 wurde auf einer bereits 2008 wiedervernässten Abtorfungsfläche im Naturschutzgebiet Provinzialmoor angelegt. Die Anlage der Kultivierungsfläche erfolgte in zwei Abschnitten in den Jahren 2015 und 2016. Zunächst wurde das Wasser im zuvor überstauten Polder abgelassen und die Fläche (150 × 150 m) zur Be- und Entwässerung mit Gräben ausgestattet (circa 0,3 m tief und 0,30,7 m breit, Abb. 2). Die Fläche war vollständig von überstauten Wiedervernässungspoldern umgeben, sodass zur Bewässerung manuell über Bogenrohre Wasser aus den benachbarten Poldern eingeleitet werden konnte. Aus einem Graben, der aktuelle Abtorfungsflächen entwässert, konnte zusätzlich Wasser gepumpt werden. Der pH-Wert in den Gräben auf der Fläche lag zwischen 3,6 und 5,4.
Im Gegensatz dazu wurde die Kultivierungsfläche Drenth (K2) direkt nach Fertigstellung der Abtorfung 2015 ohne vorangegangene langjährige Vernässung eingerichtet. Dabei wurden auf einem rund 1.000 m langen und 50 m breiten Streifen sieben Polder (jeweils circa 0,4 ha) angelegt, die durch Torfdämme voneinander getrennt sind. Die Polder waren zur Be- und Entwässerung entweder mit offenen Gräben (Abb. 2) oder mit unterirdisch verlegten Drainagerohren und darauf liegenden oberirdischen Tröpfchenschläuchen ausgestattet. Das dafür erforderliche Wasser stammte aus einem regengespeisten Teich, der bei Bedarf aus einem Grundwasserbrunnen nachgefüllt werden konnte, sodass auch bei wenig Niederschlägen dauerhaft Wasser vorhanden war. Die Zuführung von Grundwasser führte zeitweise zu höheren pH-Werten (4,48,6) in den Gräben. Zwischen den Poldern befanden sich offene Torfdämme, die zur Bewirtschaftung genutzt wurden. Angrenzend an K2 fanden sich weitere, noch aktive Torfabbauflächen.
Beide Kultivierungsflächen wurden hauptsächlich mit den Bult-TorfmoosenSphagnum papillosum undS. palustre von vier verschiedenen Spenderflächen beimpft (Graf et al. 2017). Zur Verbesserung des Mikroklimas in der Initialphase wurden die Moosflächen bei der Anlage mit Schattiervlies oder Stroh abgedeckt. Während das Schattiervlies nach etwa einem halben Jahr wieder entfernt wurde, hatte sich das Stroh zersetzt, wurde verweht oder von den Torfmoosen überwachsen. Die Flächen wurden bis zu zweimal im Jahr gemäht, um den Konkurrenzdruck durch sich ausbreitende Gefäßpflanzen zu verringern. Zudem wurden die Gräben in kleinen Abschnitten einmal im Jahr geräumt, um einer Verschlämmung entgegenzuwirken. Während der Untersuchungen hat auf den Kultivierungsflächen noch keine Torfmoosernte stattgefunden.
Bei den Kultivierungsflächen handelte es sich um offene Flächen ohne höhere Gehölze. Während K1 zum Großteil durch Wollgrasbestände dominiert wurde, waren auf K2 offene Torfflächen mit spärlicher Vegetation prägend (Tab. 1). Auf beiden Flächen fanden sich zudem vor allem Pfeifengrasbestände sowie kleinflächig Torfschlammflächen mit Schnabelriedvegetation. Neben Schlenken- und Bult-Torfmoosen fanden sich auch typische Gefäßpflanzenarten der Hochmoor-Bultvegetation wie Rosmarinheide (Andromeda polifolia ) oder Moosbeere (Vaccinium oxycoccos ) auf den Flächen. Gräben und überstaute Flächen machten nur 46 % der Fläche aus.
Wiedervernässungsflächen
Bei den Wiedervernässungsflächen handelte es sich um ehemalige Abtorfungsflächen im NSG Provinzialmoor, die durch die Anlage von Poldern (vergleiche Blankenburg 2004) mit dem Ziel der Hochmoorregeneration wiedervernässt wurden. Es haben hier keine Ausbringung von Torfmoosen oder andere gezielte Maßnahmen zur Steigerung der Biodiversität stattgefunden. Die Flächen dienten deshalb als Referenz für eine Wiedervernässung nach Abtorfung.
Die Fläche W1 wurde 1999 im Rahmen des internationalen BRIDGE-Projekts wiedervernässt (Blankenburg & Tonnis 2004), wies jedoch trockene Verhältnisse auf, sodass sich hier auch im Winter keine überstauten Wasserflächen fanden (Abb. 3). Das Zentrum des UG war auf 20 % der Fläche durch Wollgrasbestände mit vereinzelten Vorkommen von Schlenken-Torfmoosen geprägt (Tab. 1). Die Hälfte der Fläche wies Besenheide- und Pfeifengrasbestände mit Birkensukzession auf. In den Randbereichen wurde der Gehölzbestand dichter und hatte sich auf 29 % der Fläche bereits zum Wald entwickelt. Dauerhafte Gewässerstrukturen waren nicht vorhanden.
Bei W2 handelte es sich um einen 2008 wiedervernässten Polder, der im Winter flächig überstaut war (bis zu 0,5 m) und im Sommer in der Regel bis auf kleine Wasserstellen austrocknete (Abb. 3). Der überstaute Bereich machte fast 50 % der Fläche aus (Tab. 1). Der pH-Wert in der Wasserfläche lag dabei zwischen 4,1 und 4,2. Aufgrund eines unebenen Reliefs mit einzelnen Erhebungen war die Wasserfläche in mehrere Teilbereiche gegliedert und von Vegetation durchzogen. In den flach überstauten Bereichen fanden sich vor allem Wollgrasbestände, durchsetzt mit dem zum Teil flutenden Spieß-Torfmoos (Sphagnum cuspidatum ). Die trockeneren Randbereiche waren geprägt durch Besenheide- und Pfeifengrasbestände mit einem lockeren Gehölzbestand aus Birken.
Naturnahe Hochmoorflächen
Bei den naturnahen Hochmoorflächen handelt es sich um Restbestände mit einer gut erhaltenden Hochmoorvegetation (Abb. 4). Die Fläche N1 liegt in dem Landschutzschutzgebiet Wildes Moor bei Papenburg und wurde in geringem Umfang durch Weißtorfabtrag beeinträchtigt. Die ehemaligen Entwässerungsgräben wurden jedoch bereits 1985 wieder verschlossen, sodass sich die Vegetation regenerieren konnte. Bei der Fläche N2 im Naturschutzgebiet Meerkolk bei Geeste handelt es sich um einen ehemaligen Moorsee, der heute mit einem Schwingrasen zugewachsen ist und durch Torfdämme eingefasst wurde, um den Wasserstand hoch zu halten.
In beiden UG fanden sich auf 4046 % der Fläche durch Bult-Schlenken-Komplexe geprägte naturnahe Hochmoorbiotope (Tab. 1). Die restliche Fläche N1 (41 %) war vor allem durch größere Pfeifengrasbestände geprägt, die zudem auf rund 7 % der Fläche mit einem lockeren Gehölzbestand aus Birken bestanden war. Dagegen war N2 durch eine Vielzahl unterschiedlicher Biotope wie Wollgrasbestände, Glockenheide-Stadien, Pfeifengrasbestände, aber auch Schilf- oder Flatterbinsenbestände und Offenbodenbereiche sowie einer Baumgruppe geprägt. Gewässer waren auf jeweils rund 2 % der Flächen vorhanden. Der pH-Wert der Gewässer auf beiden Flächen lag zwischen 4,0 und 4,5.
2.2 Erfassung der Artengruppen
In den Jahren 2017 und 2018 erfolgte eine flächendeckende Erfassung von Vögeln, Amphibien, Libellen und Tagfaltern in allen UG, mit Ausnahme der Wiedervernässungsfläche W1, auf der keine Erfassung der Amphibien und Libellen erfolgte, weil es dort keine Gewässerstrukturen gab, die über einen längeren Zeitraum Wasser führten.
Es wurde eine Brutvogel-Revierkartierung nach Fischer et al. (2005) durchgeführt. Dazu erfolgten in jedem Jahr sechs Begehungen in den frühen Morgenstunden sowie drei Begehungen in der Dämmerung im Zeitraum von April bis Anfang Juli (vergleiche Schröder et al. 2005). Bei den Begehungen wurden sowohl Beobachtungen von Vögeln mit revieranzeigenden Merkmalen aufgenommen als auch weitere Beobachtungen für die Erfassung der Gesamtartenzahl registriert.
Für die Amphibien erfolgten in jedem Jahr sieben Begehungen im Zeitraum von März bis Anfang Juli, sowohl am Tage als auch bei Nacht (Bosbach & Ortmann 2005, Schlüpmann & Kupfer 2009). Die Erfassung erfolgte durch eine Kombination aus Keschern, Reusenfang, Verhören und Sichtbeobachtungen aller Entwicklungsstadien.
Die Erfassung der Libellen und Tagfalter erfolgte jeweils an acht bis neun Terminen bei möglichst günstigen Witterungsbedingungen (sonnig, Windstille) von Mai bis September (Chovanec 2019, Mühlhofer 1999). Die Imagines wurden mittels Kescherfang und Sichtbeobachtungen erfasst. Bei den Libellen wurde zusätzlich eine Exuviensuche durchgeführt (s. auch Zoch & Reich 2020).
2.3 Datenauswertung
Die weitergehende Datenauswertung erfolgte für die in den UG bodenständigen Arten. Dafür wurde eine vereinfachte Einordnung der Bodenständigkeit für alle Artengruppen in den einzelnen Erfassungsjahren durchgeführt. Als bodenständig wurden alle Arten gewertet, die den Kriterien in Tab. 2 entsprachen (angelehnt an Fischer et al. 2005, Schlumprecht 1999, Sternberg & Buchwald 1999).
Die Angaben zum Gefährdungsstatus in Niedersachsen entstammen den Roten Listen der Brutvögel (Krüger & Nipkow 2015), Amphibien und Reptilien (Podloucky & Fischer 2013), Libellen (Baumann et al. 2021 b) und Großschmetterlinge (Lobenstein 2004).
Die nachgewiesenen Arten wurden hinsichtlich ihrer ökologischen Bindung an Hochmoore in Niedersachsen klassifiziert. Als typische Hochmoorarten wurden sowohl Arten mit einer spezifischen Bindung an Hochmoore eingeordnet als auch Arten, die in Niedersachsen einen Verbreitungsschwerpunkt in Hochmooren haben, jedoch auch andere Lebensräume besiedeln. Die Einordnung als hochmoortypische Arten erfolgte für die Vögel nach Blüml & Sandkühler (2015), für die Amphibien nach Günther (1996), für die Libellen nach Baumann et al. (2021 a) und für die Tagfalter nach Ebert & Rennwald (1991).
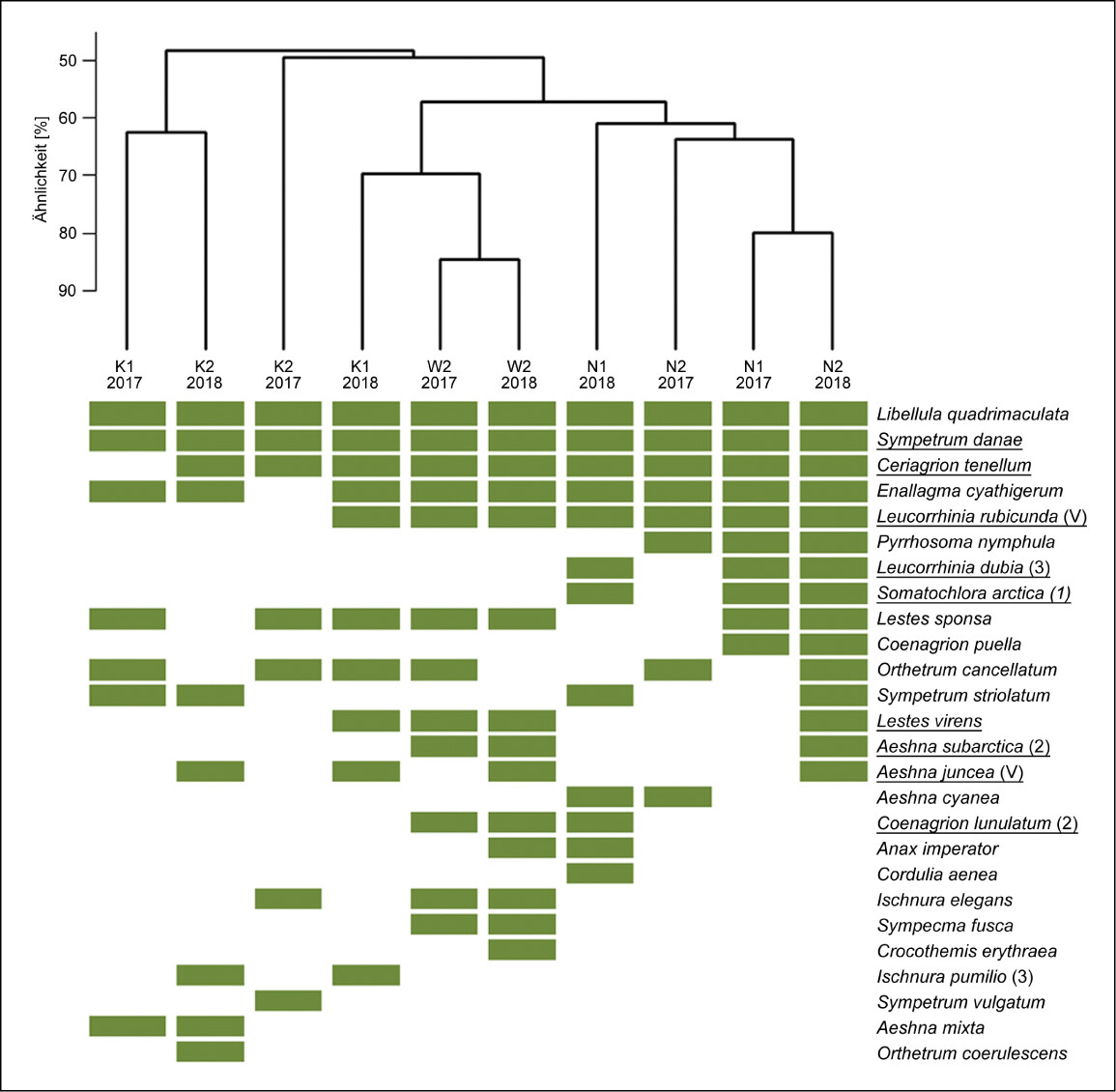
Um die Ähnlichkeit der Artenzusammensetzung der UG zu vergleichen, wurde für die Vögel und die Libellen eine hierarchische Clusteranalyse durchgeführt. Dabei wurden nur bodenständige Artvorkommen betrachtet. Für die Clusteranalyse wurde der binäre Sørensen-Ähnlichkeits-Index mit dem agglomerativen Verfahren average mit der Software R (recluster package, Dapporto et al. 2020) berechnet. Die grafische Darstellung erfolgte als Dendrogramm in Verbindung mit einer Heatmap ebenfalls mit R (gplots package, Warnes et al. 2020).
Zusätzlich erfolgte eine quantitative Auswertung, bei der für die Vögel die Anzahl der Brutpaare abgeschätzt wurde (vergleiche Südbeck et al. 2005). Bei den Amphibien wurden die Tagesmaxima an Beobachtungen von Adulten, rufenden Männchen oder Laichballen ausgewertet (Bosbach & Ortmann 2005, Schlüpmann & Kupfer 2009, Schmidt 2005). Ähnliches galt für Libellen und Tagfalter. Hier wurden jeweils die Tagesmaxima an Imagines oder im Fall von Libellen die Gesamtzahl der Exuvien gezählt.
3 Ergebnisse
Zusammengefasst wurden in allen UG 45 Vogel-, drei Amphibien-, 31 Libellen- und 22 Tagfalterarten nachgewiesen, von denen acht Vogel-, eine Amphibien-, zehn Libellen- und zwei Tagfalterarten als hochmoortypische Arten gelten.
3.1 Vögel
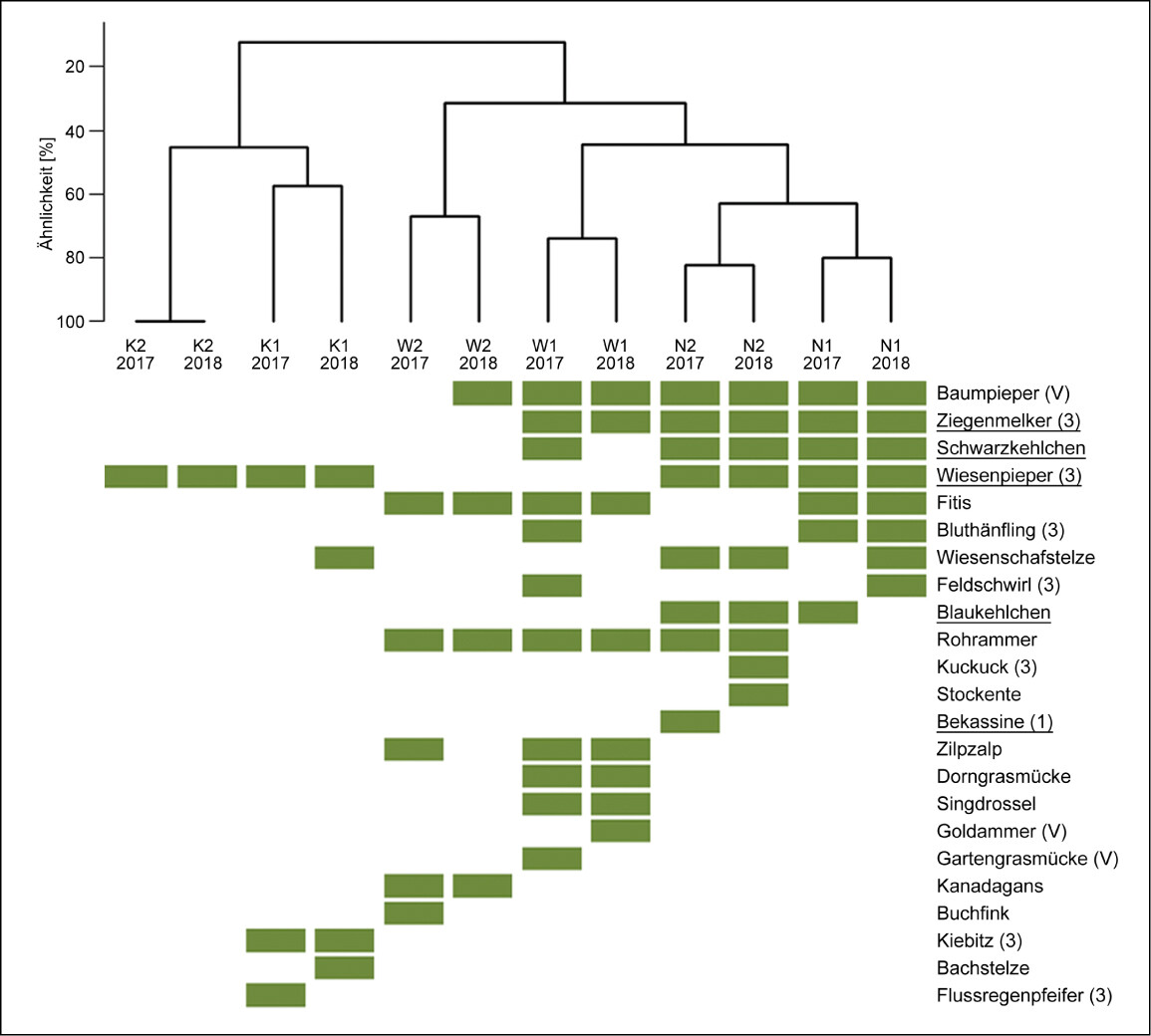
Die Brutvogelgemeinschaften (Arten mit Brutverdacht oder -nachweis) der drei Flächentypen unterschieden sich deutlich, wobei die Kultivierungsflächen die größten Unterschiede aufwiesen und die Wiedervernässungsflächen einige Übereinstimmungen mit den naturnahen Hochmoorflächen zeigten (Abb. 5). Beim Vergleich innerhalb der Flächentypen wiesen die beiden UG der naturnahen Hochmoorflächen sehr ähnliche Artengemeinschaften auf (5967 % Übereinstimmung), während es bei den beiden anderen Flächentypen deutlichere Unterschiede gab (3850 % Übereinstimmung). Auf den naturnahen Hochmoorflächen brüteten vor allem hochmoortypische Arten wie Ziegenmelker, Schwarzkehlchen, Wiesenpieper und Blaukehlchen sowie andere Arten der Halboffenlandschaft (etwa Baumpieper) oder Offenlandschaft (zum Beispiel Wiesenschafstelze, Rohrammer) (Abb. 5 sowie Tab. A1 im Online-Supplement unter Webcode NuL2231 ). Im Vergleich dazu fehlten auf den Wiedervernässungsflächen Arten wie Wiesenpieper, Blaukehlchen und Wiesenschafstelze.
Auf den Kultivierungsflächen fanden sich die wenigsten Brutvogelarten. Es wurden Wiesenpieper und Wiesenschafstelze als hochmoortypische Arten sowie Kiebitz und Flussregenpfeifer als gefährdete Arten bodenständig nachgewiesen. Weitere Beobachtungen, jedoch ohne Brutverdacht oder -nachweis wurden auf K1 von den hochmoortypischen Arten Bekassine, Lachmöwe und Rotschenkel gemacht. Auf K2 wurde zudem der in Niedersachsen vom Aussterben bedrohte Steinschmätzer mehrmals bei der Nahrungssuche beobachtet.
Die Anzahl der nachgewiesenen Brutpaare auf allen Flächen lag meist nur bei ein oder zwei Paaren je Fläche (Tab. A1 unter Webcode NuL2231 ).
3.2 Amphibien
In allen UG wurden nur Moor- und Teichfrosch bodenständig nachgewiesen (Tab. 3). Der Moorfrosch als moortypische Art hatte seinen größten Bestand auf der naturnahen Hochmoorfläche N2, wo er zudem in beiden Erfassungsjahren aufgrund des Nachweises von Laich, Larven, Juvenilen und Adulten bodenständig war. Auf N1 wurden in beiden Jahren dagegen nur juvenile und adulte Tiere nachgewiesen. Auf K1 wurden 2017 ebenfalls juvenile und adulte Tiere vom Moorfrosch beobachtet, 2018 konnte auf beiden Kultivierungsflächen zusätzlich auch Laich gefunden werden. Auf W2 wurde der Moorfrosch nur 2018 mit adulten Tieren und Laich nachgewiesen.
Die größten Bestände vom Teichfrosch wurden auf N1 und K2 erfasst. Laich wurde nur 2017 auf N2 und 2018 auf K2 gefunden. Auf den anderen Flächen oder in den anderen Jahren wurde der Teichfrosch durch die Beobachtung von juvenilen Tieren als bodenständig eingeordnet.
3.3 Libellen
Trotz einzelner gemeinsamer Arten in allen UG unterschieden sich die Artengemeinschaften der bodenständigen Libellenarten zwischen den naturnahen Hochmoorflächen und den Kultivierungsflächen deutlich (Abb. 6). Die Wiedervernässungsfläche W2 zeigte mehr Übereinstimmungen mit den naturnahen Flächen.
Zu den Arten, die ausschließlich auf den naturnahen Hochmoorflächen nachgewiesen wurden, gehörten hochmoortypische Arten: Kleine Moosjungfer (Leucorrhinia dubia ), Arktische Smaragdlibelle (Somatochlora arctica ), aber auch Generalisten wie Frühe Adonislibelle (Pyrrhosoma nymphula ), Hufeisen-Azurjungfer (Coenagrion puella ) und Blaugrüne Mosaikjungfer (Aeshna cyanea ). Die Kultivierungsflächen wiesen in beiden Jahren mit sieben bis zehn bodenständigen Arten zwar die geringsten Artenzahlen auf, darunter aber fünf hochmoortypische Arten. K1 bildete 2018 ein übergeordnetes Cluster mit W2 und den naturnahen Hochmoorflächen (Abb. 6). Ebenso wie auf den naturnahen Hochmoorflächen wurden hier die hochmoortypischen Arten Nordische Moosjungfer (Leucorrhinia rubicunda ) und Kleine Binsenjungfer (Lestes virens ) erfasst. Darüber hinaus wurde die Torf-Mosaikjungfer (Aeshna juncea ) 2018 auf N2, W2, K1 und K2 bodenständig nachgewiesen. Auf den Kultivierungsflächen fehlten jedoch weitere hochmoortypischen Arten wie Hochmoor-Mosaikjungfer (Aeshna subarctica ) und Mond-Azurjungfer (Coenagrion lunulatum ), die auf der Wiedervernässungsfläche und den naturnahen Hochmoorflächen gemeinsam vertreten waren. Vier Arten wurden ausschließlich auf einer oder beiden Kultivierungsflächen nachgewiesen, darunter Kleine Pechlibelle (Ischnura pumilio ) und Kleiner Blaupfeil (Orthetrum coerulescens ).
Bei der Abundanz der Arten zeigte sich eine zunehmende Besiedlung der Kultivierungsflächen im zweiten Untersuchungsjahr (Tab. A2 unter Webcode NuL2231 , s. auch Zoch & Reich 2020). Ab diesem Zeitpunkt ähnelten die Beobachtungszahlen der Kultivierungsflächen denen der naturnahen Hochmoorflächen. Auf der großen Wasserfläche von W2 wurden dagegen teils deutliche höhere Beobachtungszahlen erreicht.
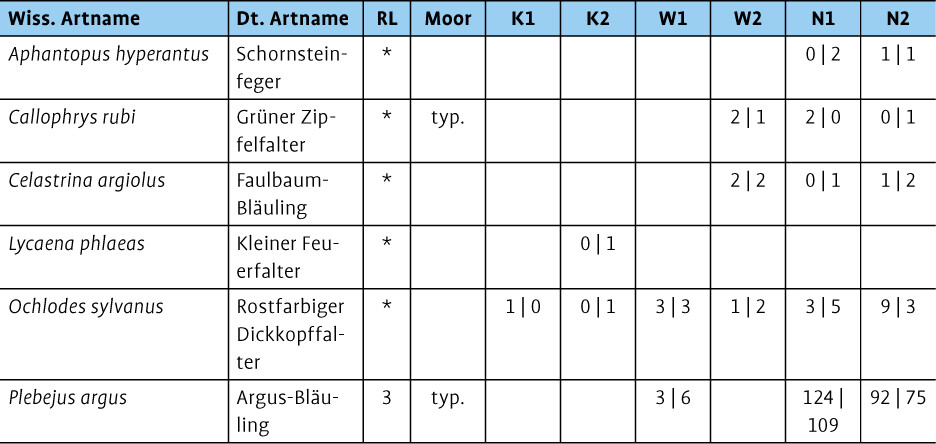
3.4 Tagfalter
Alle Untersuchungsgebiete waren mit ein bis fünf bodenständigen Tagfalterarten artenarm. Als hochmoortypische Arten wurden der Grüne Zipfelfalter (Callophrys rubi ) und der Argus-Bläuling (Plebejus argus ) festgestellt (Tab. 4). Der Argus-Bläuling hatte auf beiden naturnahen Hochmoorflächen große Bestände mit mindestens 75 Beobachtungen an einem Tag auf N2 und über 100 Beobachtungen an einem Tag auf N1. Die Art konnte ebenfalls auf W1 mit maximal sechs Beobachtungen erfasst werden. Die Nachweise vom Argus-Bläuling wurden als bodenständig eingestuft, da die Besenheide (Calluna vulgaris ) als Raupenfutterpflanze auf den Flächen große Bestände hat. Zudem wurden auf den naturnahen Hochmoorflächen Eiablagen und Kopula beobachtet.
Bei Nachweisen des Grünen Zipfelfalters handelte es sich um Einzelbeobachtungen auf W2, N1 und N2. Dennoch ist von bodenständigen Vorkommen auszugehen, da passende Futterpflanzen vorkamen (etwaCalluna vulgaris ,Frangula alnus ).
Auf den Kultivierungs- und den Wiedervernässungsflächen wurden insgesamt nur ein bis zwei Arten bodenständig nachgewiesen und meist nur mit einzelnen Individuen beobachtet. Wahrscheinlich bodenständig waren auf den Kultivierungsflächen der Kleine Feuerfalter (Lycaena phlaeas , Futterpflanze:Rumex acetosella ) und der Rostfarbige Dickkopffalter (Ochlodes sylvanus , Futterpflanzen: etwaMolinia caerulea ,Juncus effusus ).
4 Diskussion
Die Ergebnisse zeigen, dass Torfmooskultivierungsflächen bereits zwei Jahre nach der Einrichtung einigen Tierarten, darunter auch einigen hochmoortypischen Arten, einen geeigneten (Teil-)Lebensraum bieten können. Die Flächen wurden dabei sowohl als Nahrungs- wie auch als Fortpflanzungshabitat genutzt.
Vögel
Auf den Kultivierungsflächen wurden deutlich weniger Vogelarten als auf den Wiedervernässungsflächen und den naturnahen Hochmoorflächen gefunden. Ein wichtiger Einflussfaktor dafür waren wahrscheinlich die nicht vorhandenen Gehölzstrukturen auf den Kultivierungsflächen im Gegensatz zu dem lichten Gehölzbestand auf den anderen UG. So wurden auf den Kultivierungsflächen hauptsächlich typische Offenlandarten und Bodenbrüter wie etwa Kiebitz, Wiesenschafstelze und der Wiesenpieper gefunden, während Arten der Halboffenlandschaft oder der Waldränder (zum Beispiel Schwarzkehlchen, Baumpieper) fehlten. So kann auch die höhere Ähnlichkeit der Wiedervernässungsflächen mit den naturnahen Hochmoorflächen erklärt werden. Da viele Offenlandarten heute gefährdet sind (Krüger & Nipkow 2015), bieten die Kultivierungsflächen jedoch gerade für diese Arten einen wertvollen Ersatzlebensraum. Am Beispiel des Wiesenpiepers zeigt sich, dass es hochmoortypische Arten gibt, die auf Wiedervernässungsflächen unter bestimmten Bedingungen (Verbuschung oder Überstau) keine geeigneten Strukturen zum Brüten finden, während die Kultivierungsflächen mit ihren durch die Bewirtschaftung niedrig gehaltenen Vegetationsstrukturen und oberflächennahen Wasserständen offenbar gut geeignet sind. Die auf K1 bereits nachgewiesene Bekassine könnte zukünftig ebenfalls die Kultivierungsfläche zum Brüten nutzen, da die Art in Niedersachsen eine Präferenz für Wollgras-Torfmoosstadien zeigt (Blüml & Sandkühler 2015), wie sie auf K1 großflächig vorhanden sind. Die Beobachtungen von weiteren, nicht bodenständigen Arten zeigen den Einfluss der Flächenumgebung auf das Arteninventar. Die ebenfalls hochmoortypischen Arten Rotschenkel und Lachmöwe fanden vermutlich auf den die Fläche K1 umgebenden, zeitweise überstauten Wiedervernässungsflächen geeignete Brutbedingungen (vergleiche Blüml & Sandkühler 2015). Der auf K2 nachgewiesene Steinschmätzer kommt im Binnenland häufig in Torfabbaugebieten vor (Krüger et al. 2014) und nutzte die Kultivierungsfläche als Nahrungshabitat.
Amphibien
Die Eignung von Kultivierungsflächen als Habitat für (semi-)aquatische Artengruppen wie Amphibien oder Libellen hängt vor allem von der Form der Bewässerung ab. Die Exuvienfunde und Emergenzbeobachtungen der Libellen sowie Laichballenfunde der Amphibien wurden nur in den Bewässerungsgräben erbracht. Zusätzlich zu den Gräben entstanden durch Überstauungen temporäre flache Gewässer auf den Moosflächen, sowohl in den mit Gräben als auch in mit Tröpfchenschläuchen bewässerten Bereichen. Diese Wasserflächen trockneten jedoch im Verlauf des Sommers zurück, sodass eine Reproduktion hier ausgeschlossen werden kann. Bei der Bewässerung mit Gräben sollte beachtet werden, dass diese sich aufgrund der offenen Wasserflächen durch die Begünstigung von Methanbildung negativ auf die Treibhausgasbilanz der Paludikulturen auswirkt (Günther et al. 2017, Oestmann et al. 2021).
Für Amphibien können niedrige pH-Werte von sauren Hochmoorgewässern limitierende Faktoren für die Reproduktion darstellen, da sich diese negativ auf die Laich- und Larvenentwicklung auswirken (Clausnitzer 1987, Mazerolle 2005). Auch beim Moorfrosch führen pH-Werte unter 4,5 zu Ausfällen (Clausnitzer 1987). Der Bestand an Moorfröschen auf N2, die die Schlenken auf den Torfmoosschwingrasen mit pH-Werten zwischen 4,0 und 4,5 für die Reproduktion nutzten, stellt somit vermutlich eher eine Ausnahme dar. Zudem wurde auch hier eine Verpilzung der Laichballen beobachtet, sodass davon auszugehen ist, dass sich zwar ein Teil der Larven entwickeln kann, jedoch weniger als es in mesotrophen Gewässern der Fall wäre. Auch Teichfrösche erlangen außerhalb der sauren Hochmoorgewässer höhere Populationsgrößen (Mazerolle 2005). Die für das Torfmooswachstum benötigten sauren Bedingungen widersprechen somit den optimalen Voraussetzungen der Amphibien-Reproduktionsgewässer. Dennoch können Torfmooskultivierungsflächen als Alternative zur herkömmlichen landwirtschaftlichen Folgenutzung wertvolle Biotopverbundelemente und Sommerhabitate sowie in Grenzen auch Reproduktionshabitate darstellen. Insbesondere die Moorfroschbestände in Niedersachsen sind durch die räumliche Verinselung in der Agrarlandschaft gefährdet (Fischer & Podloucky 2008) und könnten durch die Anlage von Paludikulturen unterstützt werden.
Libellen
Die mit Gräben bewässerten Kultivierungsflächen eignen sich ebenfalls als Lebensraum für Libellen (s. auch Zoch & Reich 2020). Die Libellenfauna der Kultivierungsflächen war durch häufige Arten wie Vierfleck (Libellula quadrimaculata ), Becherjunger (Enallagma cyathigerum ) oder Schwarze Heidelibelle (Sympetrum danae ) geprägt, die in Norddeutschland typischerweise auch Moorgewässer besiedeln (Baumann et al. 2021 a). Auf beiden Kultivierungsflächen waren auch Hochmoore präferierende Arten wie die Scharlachlibelle oder die Torf-Mosaikjungfer (Aeshna juncea ) bodenständig. Für die Torf-Mosaikjungfer bieten die Kultivierungsflächen günstige Eiablageplätze, solange die Gräben nicht zu stark zugewachsen und deshalb offene, steile Torfkanten an den Ufern vorhanden sind (vergleiche Sternberg 1990). So wurden von allen UG die meisten Eiablagen der Torf-Mosaikjungfer auf den Kultvierungsflächen beobachtet. Erwähnenswert ist zudem der Nachweis des Kleinen Blaupfeils (Orthetrum coerulescens ), der auf beiden Kultivierungsflächen, nicht jedoch in einem der anderen UG erbracht werden konnte. Die zeitweise langsam fließenden Gräben der Torfmooskultivierungsflächen könnten den Ansprüchen der in Niedersachsen in Ausbreitung befindlichen Art entsprechen (vergleiche Baumann et al. 2021 a).
Auch wenn bei der Artenzahl keine relevanten Unterschiede zwischen den UG festgestellt werden konnten, zeigte sich bei der Artenzusammensetzung, dass insbesondere die stark spezialisierten Hochmoorarten Arktische Smaragdlibelle (Somatochlora arctica ), Kleine Moosjungfer (Leucorrhinia dubia ), Hochmoormosaikjungfer (Aeshna subarctica ) bisher auf den Kultivierungsflächen fehlten. Während Hochmoormosaikjungfer und Kleine Moosjungfer Wiedervernässungsflächen und neu angelegte Moorgewässer besiedeln und deshalb auch auf den Kultivierungsflächen zukünftig zu erwarten wären, fehlt die Arktische Smaragdlibelle in Niedersachsen auf vollständig abgetorften, wiedervernässten Hochmoorflächen (Baumann et al. 2021 a) und ist somit auf den Erhalt der naturnahen Restflächen angewiesen.
Die extreme Trockenheit im Jahr 2018 (70 % des mittleren Jahresniederschlags in Lingen DWD 2022) hat dazu geführt, dass auf K1 ab Anfang August die Bewässerungsgräben trockenfielen, da kein Bewässerungswasser mehr verfügbar war. Dagegen konnte auf K2 durch die Zuführung von Grundwasser weiter bewässert werden. Infolgedessen waren auf K1, ebenso wie auf den Vergleichsflächen ohne Wassermanagement, deutliche Bestandseinbrüche zu verzeichnen und nur noch wenige Libellenindividuen zu finden. Dagegen wurden die späten Arten wie etwa die Scharlachlibelle (Ceriagrion tenellum ) auf K2 auch im August und September noch häufig erfasst (Zoch & Reich 2020). Das zeigt die Bedeutung von Kultivierungsflächen mit künstlicher Bewässerung als Rückzugsort für (semi)aquatische Artengruppen in trockenen Jahren.
Tagfalter
Für die Tagfalter erwiesen sich die Kultivierungsflächen als wenig geeignet, obwohl die für die hochmoortypischen Arten relevanten Nektar- und Raupenfutterpflanzen (etwa Glockenheide) häufig und auch blühend zu finden waren. Andere, für Tagfalter relevante Habitatparameter wurden von den Kultivierungsflächen jedoch nicht erfüllt. Der fehlende Windschutz auf den Flächen ist vermutlich ein wichtiger Grund für die geringe Anzahl nachgewiesener Arten und Individuen. An Hochmoore angepasste Arten nutzen auch in naturnahen Hochmooren hauptsächlich die windgeschützten Randbereiche, während sie die offenen Zentren eher meiden (Burmeister 1990). Zudem waren auf den Kultivierungsflächen nur vereinzelt Ameisen vorhanden (eigene Erfassungen 20172018), die eine wichtige Rolle bei der Larvalentwicklung von verschiedenen Bläulingsarten spielen (zum Beispiel Argus-Bläuling Eeles 2019). Die Bewirtschaftungsaspekte der Kultivierungsflächen (Mahd, potenzielle Torfmoosernte) beeinträchtigen darüber hinaus die Habitatqualität für Tagfalter (Verlust von Präimaginalstadien, fehlendes Nektarangebot und Windschutz).
5 Schlussfolgerungen und Empfehlungen
Die nasse Bewirtschaftung von Hochmoorböden im Rahmen der Torfmooskultivierung bietet nicht nur aus Klimaschutzsicht großes Potenzial (Günther et al. 2017, Oestmann et al. 2021). Es hat sich gezeigt, dass die Kultivierungsflächen zum Teil den gleichen Arten einen Lebensraum bieten wie die ungenutzten Wiedervernässungsflächen, wobei auf den untersuchten Wiedervernässungsflächen größere Übereinstimmungen in der Artenzusammensetzung mit den naturnahen Flächen gefunden wurden. Dabei muss berücksichtigt werden, dass die Kultivierungsflächen nur ein bis drei Jahre nach der Etablierung erfasst wurden, sodass davon auszugehen ist, dass sich das Arteninventar über die Jahre noch ändern kann (vergleiche Muster et al. 2020). Darüber hinaus stellten die Kultivierungsflächen mit ihren oberflächennahen Wasserständen sowie den großflächigen Torfmoosrasen Strukturen bereit, die auf den Wiedervernässungsflächen nicht zu finden waren. Deshalb könnte die Anlage weiterer Torfmooskultivierungsflächen die Vielfalt an hochmoortypischen Lebensraumstrukturen neben Wiedervernässungsflächen bereichern und insbesondere Offenlandarten und ausgewählte hochmoortypische Arten in ihren Beständern fördern (vergleiche Muster et al. 2015). Dennoch sollten die künstlichen Systeme der Torfmooskultivierungsflächen nicht in Flächenkonkurrenz mit ungenutzten Wiedervernässungsflächen treten. Sie sollten vielmehr herkömmliche landwirtschaftliche Nutzungen auf Hochmoorböden ersetzen, da diese entwässerungsbasierten Nutzungsformen keinen Lebensraum für spezialisierte Hochmoorarten bieten.
Trotz der nachgewiesenen Eignung als Nahrungs- und Fortpflanzungshabitate für verschiedene Artengruppen handelt es sich bei den Kultivierungsflächen primär um bewirtschaftete Flächen mit den damit verbundenen Eingriffen. Eine Mahd der Gefäßpflanzen oder die Ernte der Torfmoose können sich letal auf einige Tierarten und ihre Entwicklungsstadien (etwa Gelege von Bodenbrütern, Eier, Puppen oder Raupen von Tagfaltern) auswirken und auf diese Weise die Populationen schwächen (vergleiche Humbert et al. 2009, Muster et al. 2020). Durch die Mahd werden außerdem die vertikalen Vegetationsstrukturen zeitweise zerstört, sodass weniger Strukturvielfalt als auf naturnahen oder nicht genutzten Wiedervernässungsflächen zu finden ist. Auch das regelmäßige Räumen der Gräben (Gaudig et al. 2018) kann in Abhängigkeit von der Ausführung Auswirkungen auf das Makrozoobenthos haben. Für eine Verbesserung der Lebensraumbedingungen auf den Kultivierungsflächen sollten Pflegeeingriffe wie die Mahd oder die Räumung von Gräben möglichst gering gehalten und abschnittsweise oder mosaikartig durchgeführt werden (Muster et al. 2015). Eine weitere Möglichkeit wäre die Förderung von zusätzlichen Habitatelementen. Beispielsweise könnten kleine Teile der Torfmoosflächen dauerhaft nicht beerntet werden, um hier die Ausbildung eines Bult-Schlenken-Systems zu ermöglichen. Durch solche Rückzugsbereiche könnte eine schnelle Wiederbesiedlung von geernteten Flächen ermöglicht werden, die nur in Zeiträumen von drei bis fünf Jahren beerntet werden (Gaudig et al. 2014) und dazwischen als ungestörter Lebensraum zur Verfügung stehen. Für Tagfalter und Libellen können Windschutzstrukturen förderlich sein. Während Baumgruppen zu Konflikten mit Offenlandarten der Vögel führen können (eingeschränkte Sicht, Sitzwarten für Prädatoren), sind für die Insekten auch höhere Gräser als Windschutz bereits interessant (Dennis 2004), weshalb nicht gemähte Streifen auch hier eine mögliche Aufwertung darstellen würden.
Neben dem hohen Potenzial für Vögel und Libellen bieten Torfmooskultivierungsflächen auch für epigäische Wirbellose wie Spinnen (vergleiche Muster et al. 2015, Muster et al. 2020), Kurzflügler oder Laufkäfer (eigene Erfassungen, Mainda 2021)) ein großes Potenzial. Da Torfmooskultivierungsflächen in Deutschland erst seit wenigen Jahren erprobt werden, ist es wichtig, auch die längerfristige Entwicklung dieser Flächen zu beobachten, nicht nur um Bewirtschaftungs- und Pflegemaßnahmen zu bewerten, sondern auch um weitere Hinweise für eine Optimierung der nutzungsfreien Wiedervernässung von Mooren zu gewinnen.
Kontakt

Lotta Zoch, M.Sc., studierte von 2010 bis 2016 Umweltplanung in Hannover. Seit 2016 wissenschaftliche Mitarbeiterin am Institut für Umweltplanung (Arbeitsgruppe Naturschutz und Landschaftsökologie) an der Leibniz Universität Hannover. Ihr Schwerpunkt ist die Untersuchung der Auswirkungen von nachhaltigen Nutzungsformen wie der Paludikultur und von Renaturierungsprojekten auf die Hochmoorfauna.
> zoch@umwelt.uni-hannover.de

Prof. Dr. Michael Reich leitet seit 1999 die Arbeitsgruppe Naturschutz und Landschaftsökologie am Institut für Umweltplanung der Leibniz Universität Hannover. Er beschäftigt sich in mehreren Forschungsvorhaben mit dem Schutz der faunistischen Biodiversität durch eine klimagerechte Regeneration von Mooren.
> reich@umwelt.uni-hannover.de
Von Lotta Zoch und Michael ReichEingereicht am 22.03.2022, angenommen am 10.09.2022
kompletter Artikel im PDF
-
Abb. 2: Die Kultivierungsflächen K1 im Provinzialmoor (oben) und K2 in Drenth (unten) mit Gräben und in ungemähtem ZustandL. Zoch -
Abb. 1: Lage der sechs Untersuchungsgebiete im westlichen NiedersachsenGrafik: Zoch 2022 -
Abb. 3: Die Wiedervernässungsflächen W1 (oben) und W2 (unten) im ProvinzialmoorL. Zoch -
Abb. 4: Die naturnahen Hochmoorflächen N1 im Landschaftsschutzgebiet Wildes Moor (oben) und N2 im Naturschutzgebiet Meerkolk (unten)L. Zoch -
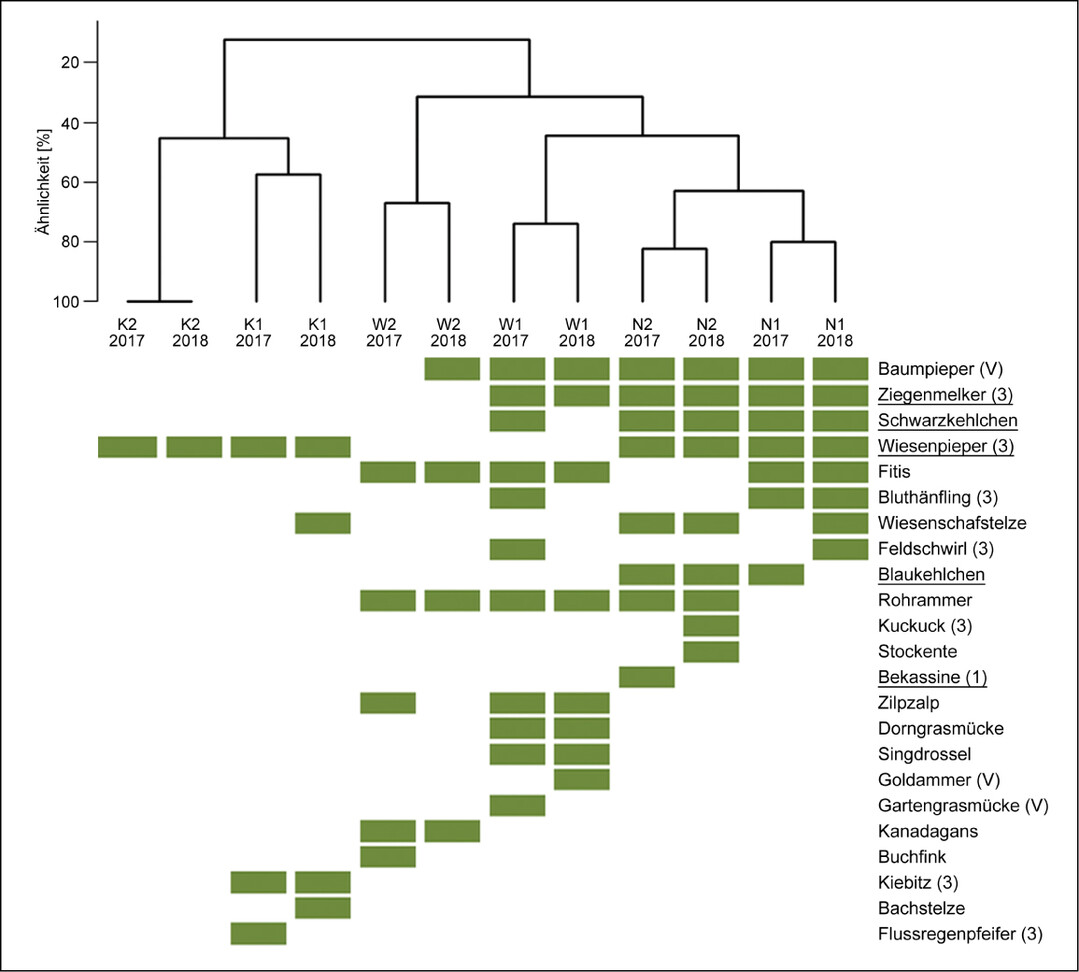
Abb. 5: Ähnlichkeit der Brutvogelgemeinschaften (Grün = Vorkommen mit Brutverdacht oder -nachweis) zwischen den Untersuchungsgebieten (K = Kultivierung, W = Wiedervernässung, N = Naturnah) und Jahren anhand des Sørensen-Index. Hochmoortypische Arten sind unterstrichen, der Gefährdungsstatus (V = Vorwarnliste, 3 = gefährdet, 2 = stark gefährdet, 1 = vom Aussterben bedroht) gemäß der Roten Liste von Niedersachsen und Bremen (Krüger & Nipkow 2015) wird in Klammern hinter der Art aufgeführt.Zoch 2022 -
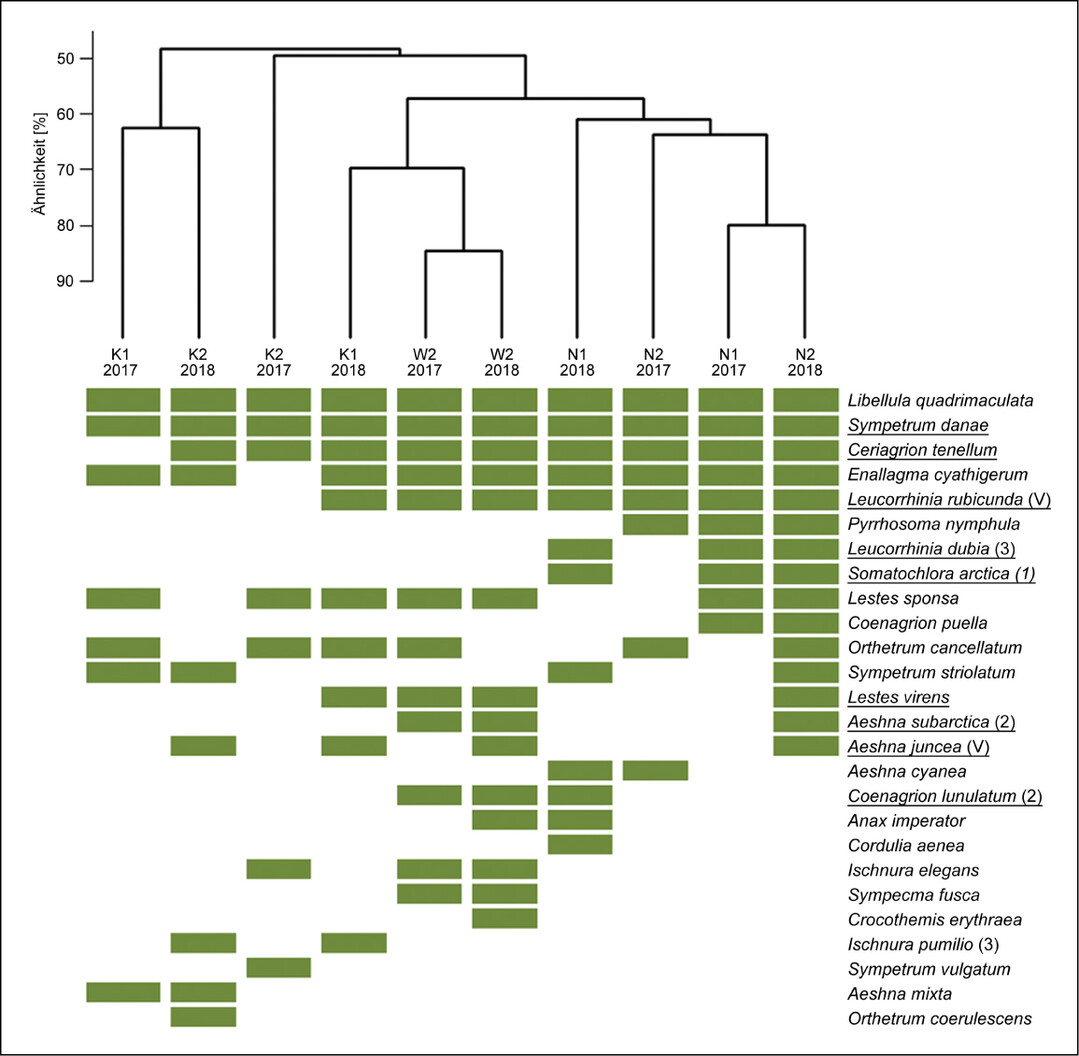
Abb. 6: Ähnlichkeit der Libellen-Zönosen (Grün = Vorkommen mit Bodenständigkeitshinweis) zwischen den Untersuchungsgebieten (K = Kultivierung, W = Wiedervernässung, N = Naturnah) und Jahren anhand des Sørensen-Index. Hochmoortypische Arten sind unterstrichen, der Gefährdungsstatus (V = Vorwarnliste, 3 = gefährdet, 2 = stark gefährdet, 1 = vom Aussterben bedroht) gemäß der Roten Liste von Niedersachsen und Bremen (Baumann et al. 2021 b) wird in Klammern hinter der Art aufgeführt.Zoch 2022 -
-
-
-
-
-













Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.