Räumliche Habitatanalyse für die Grüne Mosaikjungfer und die Helm-Azurjungfer
Abstracts
Nutzungswandel und -intensivierung der Landschaft bewirken einen Bestandsrückgang von Populationen bis hin zu deren lokalem oder regionalem Aussterben. Die Entwicklung von Habitatverbundsystemen ist eine Möglichkeit, dem Artenrückgang entgegenzuwirken. Für eine Etablierung von Habitatverbundsystemen sind u. a. Kenntnisse zu den Populationen der Art sowie zu deren Ausbreitungsverhalten unabdingbar.
In Anlehnung an den GIS-Algorithmus HABITAT-Net vonHänel(2007) wurde für die beiden FFH-Libellenarten Grüne Mosaikjungfer ( Aeshna viridis ) und Helm-Azurjungfer ( Coenagrion mercuriale ) in Nordwest-Deutschland eine Analyse der Habitatvernetzung durchgeführt, als Grundlage für die Planung von Habitatverbundsystemen. Die Ergebnisse zeigen, dass ab einer angenommenen Wanderungsdistanz von 9 km eine Vernetzung der einzelnen A. viridis -Lebensräume zwischen Oldenburg (Niedersachsen) und Bremen weitestgehend gegeben ist. Das Gebiet Hollerland in Bremen wird bei der höchsten Distanzklasse von 14 km durch Barrieren von den restlichen Gebieten getrennt. Die einzelnen C. mercuriale -Habitate sind im betrachteten Grenzraum zwischen Nordrhein-Westfalen und Niedersachsen bei einer maximalen Wanderungsdistanz von 6,6 km teilweise miteinander verbunden. Offen bleibt, inwieweit die vorhandenen Fließgewässer und Gräben als Vernetzungskorridore für die beiden Arten und die verschiedenen Landschaftselemente als Barrieren wirken. Für den dauerhaften Schutz der beiden Arten sollte in Zukunft mittels Maßnahmen zur Erhaltung, Optimierung und Vernetzung der Lebensräume ein regionales Habitatverbundsystem entwickelt werden.
Basics for the planning of habitat connectivity systems Spatial habitat analysis of the occurrence of the green hawker and the southern damselfly in northwest Germany
The intensification of land use and the degradation of semi-natural landscapes has resulted in a decline in populations and to local or regional extinction. One possibility to counteract this decline is the development of habitat connectivity. For successful development of habitat connectivity, a knowledge about populations and their dispersal behaviour is important.
Based on the GIS algorithm HABITAT-Net ofHänel(2007), for the two Habitats Directive dragonfly species Green Hawker (Aeshna viridis ) and Southern Damselfly (Coenagrion mercuriale ) in Northwest Germany, an analysis of habitat networks was carried out as the basis for the planning of habitat connectivity. The results show that, up to a supposed dispersal distance of 9 km, a largely connected system of the A viridis habitats may exist between Oldenburg (Lower Saxony) and Bremen. The Hollerland area in Bremen is separated from the remaining areas by barriers at the highest distance class of 14 km. TheC. mercuriale habitats in the border region between North Rhine-Westphalia and Lower Saxony are only partially connected with each other at a maximum migration distance of 6.6 km. It remains unclear how the existing streams and ditches are used as networking corridors by the two species and how the various landscape elements act as barriers. For the permanent conservation of the two species in the future, a regional habitat network should be constructed using measures for the conservation, improvement, and connectivity of the habitats.
- Veröffentlicht am

1 Einleitung
Nutzungswandel und -intensivierung der Landschaft führten und führen zu Degradierung, Verlust und Fragmentierung von Lebensräumen. Daraus resultieren ein Bestandsrückgang von Arten bis hin zum lokalen oder regionalen Aussterben von Populationen sowie ein Verlust von Lebensräumen und Landschaften. Aquatische Ökosysteme sind neben anderen Lebensräumen besonders stark von anthropogenen Einflüssen betroffen (Dudgeonet al. 2006) und gehören in Deutschland zu den am stärksten bedrohten Lebensräumen (Ellwangeret al. 2012). Die Beeinträchtigung von Gewässern führt somit zu einem Rückgang und einer Gefährdung von Süßwasserorganismen, u. a. von Libellen (Clausnitzeret al. 2009).
Viele Arten bilden Metapopulationen, indem ihre Teilpopulationen durch einen Austausch von Individuen miteinander in Verbindung stehen (Hanski1999). Dabei spielt die umgebende Landschaft eine entscheidende Rolle für den Individuenaustausch (u. a.Poniatowskiet al. 2016,Ricketts2001). Für den Naturschutz ist das Metapopulationskonzept besonders dann relevant, wenn die Metapopulation im Vergleich zur lokalen Teilpopulation eine höhere Überlebenschance hat. Dabei bildet es die theoretische Grundlage von Biotop-/Habitatverbundsystemen zum Individuenaustausch zwischen Teilpopulationen oder zu deren Neugründung (Reich&Grimm1996,Setteleet al. 1996). Für die erfolgreiche Etablierung von Habitatverbundsystemen mit Wanderkorridoren und Trittsteinen sind Kenntnisse zu aktuellen (Teil-)Populationen und potenziellen Habitaten der Art, zum Ausbreitungsverhalten sowie zu den positiven oder negativen Einflüssen von Landschaftselementen auf den Austausch von Individuen unabdingbar. Habitatverbundsysteme können daher immer nur auf Zielarten bezogen geplant und entwickelt werden (Reich&Grimm1996).
Im Rahmen der Fauna-Flora-Habitat-Richtlinie zur Erhaltung der natürlichen Lebensräume sowie der wildlebenden Tiere und Pflanzen soll ein kohärentes europäisches ökologisches Netz besonderer Schutzgebiete (Natura 2000) entstehen. Im Bundesnaturschutzgesetz ist die Schaffung eines länderübergreifenden Biotopverbunds auf mindestens 10 % der Landesfläche festgeschrieben (§§ 20 und 21 BNatSchG). Dies bedeutet nicht nur die Schaffung eines Biotopverbunds von Flächen nationaler oder internationaler Bedeutung, sondern auch die Kontinuität eines kleinräumigeren, regional-überregionalen Biotopverbundes (Burkhardtet al. 2003). Neben allgemeinen Kriterien werden Zielarten, die in der Regel Metapopulationen ausbilden, zur Auswahl von Flächen für den Biotopverbund aufgeführt. Als zwei der 187 Zielarten werden die Libellenarten Grüne Mosaikjungfer ( Aeshna viridis ) und Helm-Azurjungfer ( Coenagrion mercuriale ) genannt (Burkhardtet al. 2010).
Aufbauend auf den Kenntnissen zu ökologischen Ansprüchen, Ausbreitungsverhalten und den aktuellen Vorkommen von A. viridis in der Hunte-Weser-Niederung zwischen Oldenburg (Niedersachsen) und Bremen sowie von C. mercuriale im Grenzraum zwischen Nordrhein-Westfalen und Niedersachsen fand eine GIS-basierte Analyse der Habitatvernetzung statt, die als Grundlage für die zukünftige Planung von Habitatverbundsystemen dienen soll.
2 Zielarten
Aeshna viridis
In Europa ist A. viridis (Abb. 1) eng an die Krebsschere ( Stratiotes aloides ) als Eiablagesubstrat und Larvalhabitat gebunden und kann als Habitatspezialist und Kennart von Krebsscheren - Gewässern bezeichnet werden (Wildermuth&Martens2014). Die Art besiedelt, sofern dichte/ausgedehnte S. aloides -Bestände vorhanden sind, Fließgewässer mit geringer Strömung sowie Stillgewässer. Das Verbreitungsgebiet von A. viridis reicht von den Niederlanden über Norddeutschland, Skandinavien und Osteuropa bis nach Westsibirien (Wildermuth&Martens2014). In Deutschland (Rote Liste 2, Anhang IV FFH-Richtlinie) ist die Art auf das Norddeutsche Tiefland begrenzt (Klugkistet al. 2015) und kommt in Niedersachsen (Rote Liste 1) an Aller und Elbe, in der Hunte- und der Weserniederung bei Bremen sowie in der Emsniederung bei Leer vor.
Zum Ausbreitungsverhalten von A. viridis existieren keine Untersuchungen. In der Literatur finden sich nur wenige Angaben, so gilt die Art als ortstreu, wobei sie sich bei der Reifung mehrere Kilometer weit vom Gewässer entfernt (Sternberg2000).Mauersbergeret al. (2005) schildern einen Fund 4 km vom Fortpflanzungsgewässer entfernt. Im Rahmen eines Wiederansiedlungsprojekts von A. viridis konnten Individuen 2 km entfernt vom Ansiedlungsgewässer festgestellt werden (Körner&Marxmeier2018). Grundsätzlich kann bei Großlibellen von einem Tagesaktionsradius von bis zu 10 km ausgegangen werden (Sternberg&Buchwald1999; s. auch Tab. 1). Es ist, gerade wenn man die Maximaldistanzen berücksichtigt, die für die Kleinlibelle C. mercuriale ermittelt werden konnten (s. u.), sehr wahrscheinlich, dass 10 km eher im unteren Bereich möglicher Ausbreitungsdistanzen von Aeshniden liegen.
Coenagrion mercuriale
In Mitteleuropa besiedelt C. mercuriale (Abb. 2) überwiegend Wiesenbäche und -gräben, seltener Abflüsse von Quellmooren und -sümpfen oder Quellgewässer in Auen (Buchwald1989,Burbachet al. 2015). Charakteristisch für die Fließgewässer sind eine Quellnähe und/oder Grundwasserbeeinflussung, damit verbunden ein seltenes oder fehlendes Austrocknen, eine geringe bis mittlere Fließgeschwindigkeit, fehlendes Durchfrieren im Winter sowie geringe bis mittlere Nährstoff- und hohe Sauerstoffgehalte. Darüber hinaus spielt eine gut ausgeprägte submerse Vegetation für das Vorkommen der Art eine große Rolle (Buchwald1989, 1994). Der Verbreitungsschwerpunkt der Art liegt in Frankreich, Spanien, Portugal und Italien. Die nördlichsten Vorkommen befinden sich in England und in Nordwest-Deutschland (Wildermuth&Martens2014). In Deutschland (Rote Liste 2, Anhang II FFH-Richtlinie) kommt C. mercuriale in Süddeutschland sowie in Teilen West-, Mittel-, Nordwest- und Südost-Deutschlands vor (Burbachet al. 2015). In Nordrhein-Westfalen (Rote Liste 2) wurde die Art in den Einzugsgebieten von Lippe, Ems, Hunte und Weser nachgewiesen, in Niedersachsen (Rote Liste 1) u. a. im Einzugsgebiet von Hase, Hunte und Weser.
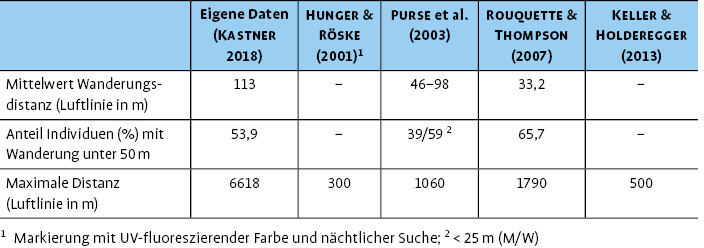
Das Ausbreitungsverhalten von C. mercuriale ist in verschiedenen Mark-recapture-Studien untersucht worden (Tab. 2). Insgesamt ist das Ausbreitungspotenzial dieser Art außerordentlich gering; über die Hälfte der Individuen bleiben vor Ort oder legen Distanzen von unter 50 m zurück, und die mittlere Wanderungsdistanz liegt zwischen 33 m und 113 m (Tab. 2). Die größte maximale Wanderungsdistanz (Luftlinie), die für C. mercuriale nachgewiesen wurde, beträgt 6,6 km (Kastner2018).
3 Methode
Die Analyse der Habitatvernetzung wurde in Anlehnung an den GIS-Algorithmus HABITAT-Net vonHänel(2006, 2007) durchgeführt. Dabei sind Funktionsräume (= miteinander verbundene Lebensräume) verschiedener Ausbreitungsdistanzklassen mittels Verwendung von GIS-Anwendungen (Hänel2007) erstellt worden. In einem ersten Schritt werden dabei alle potenziellen Habitate der Art selektiert. Im zweiten Schritt werden benachbarte Habitate auf Basis der niedrigsten Distanzklasse zu Funktionsräumen der Stufe 1 mit den ermittelten Verbindungsräumen (= Korridore) der Stufe 1 zusammengefasst. Dort, wo flächige Barrieren vorliegen, sind die Funktionsräume unterbrochen und kommen nicht zustande. Basierend auf der nächsthöheren Distanzklasse werden dann die nächsthöheren Verbindungsräume der Stufe 2 erzeugt, welche die zuerst gebildeten Funktionsräume der Stufe 1 zu Funktionsräumen der Stufe 2 zusammenfassen (Hänel2007).
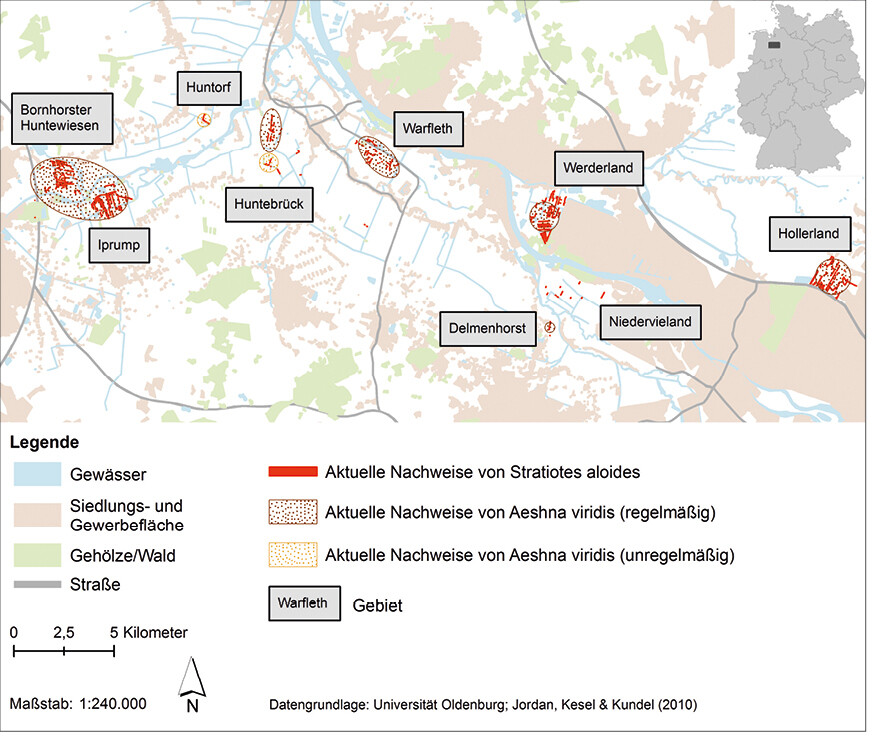
Grundlage der Verbindungs- und Funktionsräume für die Vernetzungsanalyse der A. viridis- Populationen waren die aktuellen Vorkommen von S. aloides als Lebensraum der Art in der Hunte-Weser-Niederung zwischen Oldenburg und Bremen (Abb. 3; Datengrundlage: eigene Daten,Jordanet al. 2010). Als potenzielle Ausbreitungsdistanzen wurde eine Spanne von 500 m (Nahausbreitung) bis 14 km (Fernausbreitung) verwendet. Berücksichtigt werden muss jedoch, dass es sich wohl eher um Werte im unteren Bereich möglicher Entfernungen von A. viridis handelt. Als Ausbreitungsbarrieren wurden Siedlungs- und Gewerbeflächen sowie ausgedehnte Gehölzbestände und Waldgebiete in der Analyse berücksichtigt.
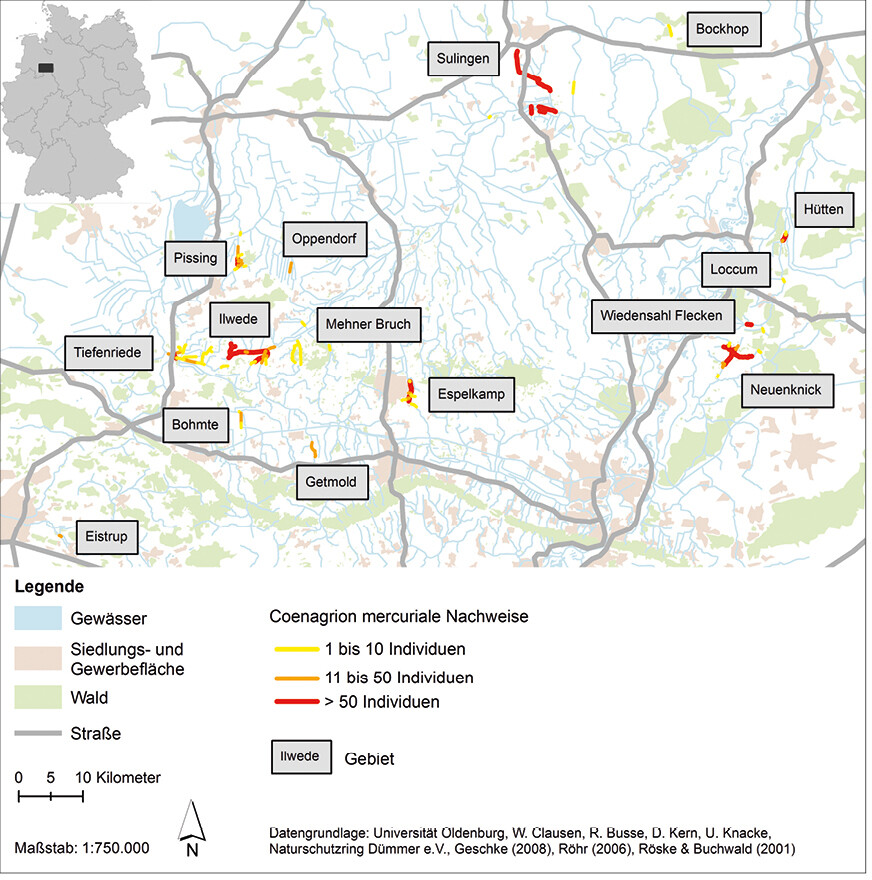
Als Grundlage für die Erstellung der Verbindungs- und Funktionsräume der Habitatvernetzungsanalyse von C. mercuriale dienten die aktuellen Vorkommen der Art im Grenzraum zwischen Nordrhein-Westfalen und Niedersachsen (Abb. 4; Datengrundlage: eigene Daten, W. Clausen, R. Busse, D. Kern, U. Knacke, Naturschutzring Dümmer e.V.,Geschke2008,Röhr2006,Röske&Buchwald2001). Die verwendeten Ausbreitungsdistanzen reichten von 25 m (Nahausbreitung; Aktionsradius eines Großteiles der Population) bis 6618 m (Fernausbreitung; maximal nachgewiesene Distanz der Art in Luftlinie). Als Ausbreitungsbarrieren sind Siedlungs- und Gewerbeflächen sowie ausgedehnte Gehölzbestände und Waldgebiete in die Analyse mit eingeflossen.
Angemerkt werden muss, dass der HABITAT-Net Algorithmus für terrestrische Lebensräume entwickelt wurde und nicht für aquatische Systeme (Fließgewässer) oder hochmobile Arten (Hänel2007). Libellen sind flugfähige/mobile Lebewesen, die in der Lage sind, Barrieren auszuweichen und größere Strecken direkt zurückzulegen, und die durch Wind verdriftet werden können. Daher ist die Ermittlung der Verbindungs- und Funktionsräume nur eine Annäherung an mögliche Wanderungswege und -mechanismen sowie potenzielle Habitate.
4 Ergebnisse
Aeshna viridis
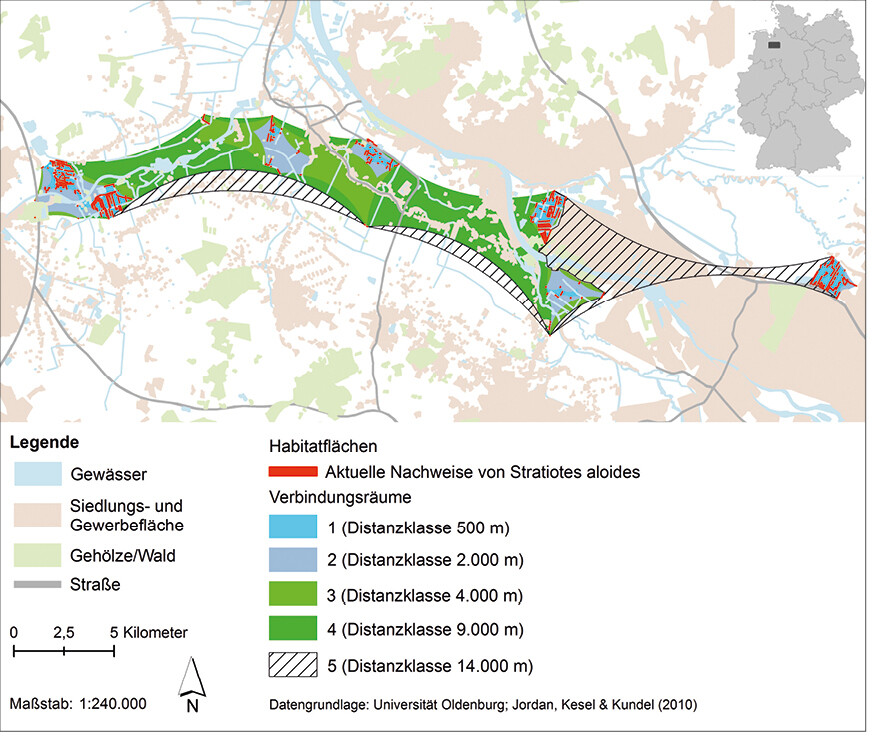
Die Ergebnisse der Vernetzungsanalyse (Abb. 5) zeigen, dass bei der Distanzklasse 500 m ein Austausch der Individuen innerhalb der einzelnen Gebiete mit Krebsscheren-Beständen stattfindet und ein Austausch von Individuen zwischen den Gebieten Bornhorster Huntewiesen und Iprump möglich ist. Alle anderen Gebiete sind bei dieser Distanzklasse voneinander isoliert, ebenso wie einzelne Habitate innerhalb des Gebiets Huntebrück. Auch bei der Distanzklasse 2 km bleiben die meisten Gebiete voneinander getrennt. Jedoch ist das isolierte Habitat im Süden von Huntebrück nun mit den Habitaten im Norden verbunden, und das Vorkommen in Delmenhorst steht im Austausch mit dem Gebiet Niedervieland. Bei der Distanzklasse 4 km stehen auch die Gebiete Huntorf, Huntebrück und Warfleth miteinander im Austausch. Das Gebiet Werderland wird weiterhin von den Gebieten Niedervieland und Delmenhorst durch größere Gehölzbestände, ein schmales Siedlungsband sowie kleinere Gewerbeflächen getrennt. Ab der Distanzklasse 9 km sind alle Gebiete zwischen Oldenburg und Bremen, bis auf das Gebiet Hollerland, miteinander verbunden. Bei der Distanzklasse 14 km würde das Gebiet Hollerland mit den restlichen Gebieten verbunden sein, jedoch wird der Verbindungsraum durch Siedlungs-/Gewerbeflächen unterbrochen (Abb. 5). Auch bei noch größeren Distanzklassen als 14 km wird eine vollständige Vernetzung aller Gebiete durch Siedlungs-/Gewerbeflächen verhindert.
Bekannt ist, dass die umgebende Landschaft bei Wanderungsbewegungen eine entscheidende Rolle spielt. Für A. viridis kann jedoch aktuell nicht beurteilt werden, welche Barrierewirkung verschiedenen Landschaftselemente (u. a. Größe und Art des Elements) ausüben. Offen bleibt darüber hinaus auch, inwieweit Fließgewässer und Gräben als Vernetzungskorridore in den Funktionsräumen und zwischen diesen wirken. Wie oben erwähnt, liegen zum Ausbreitungsverhalten von A. viridis nur einzelne Zufallsbeobachtungen vor. Somit sind die gewählten Distanzklassen vermutlich zu niedrig angesetzt, denn grundsätzlich kann bei Aeshniden von einer größeren Flugdistanz (4 km und mehr) ausgegangen werden.
Neben der dauerhaften Erhaltung und Optimierung aller S. aloides -Bestände durch eine ökologische Grabenunterhaltung nach dem Vorbild Bremens (Kunzeet al. 2012,Nagler&Müller2012) ist eine weitere Vernetzung der einzelnen Gebiete durch die Entwicklung neuer A. viridis -Habitate ( S. aloides -Bestände) wünschenswert. Eine wirksame Maßnahme stellt hierbei die Wiederansiedlung von S. aloides dar (Kastneret al. 2016).
Coenagrion mercuriale
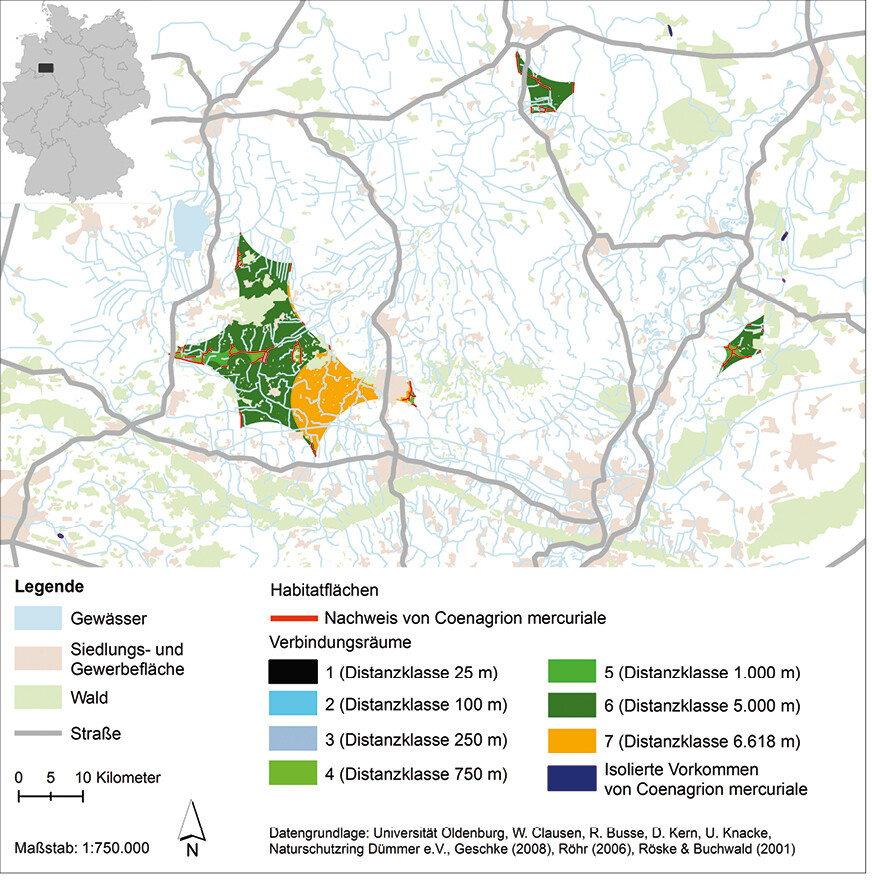
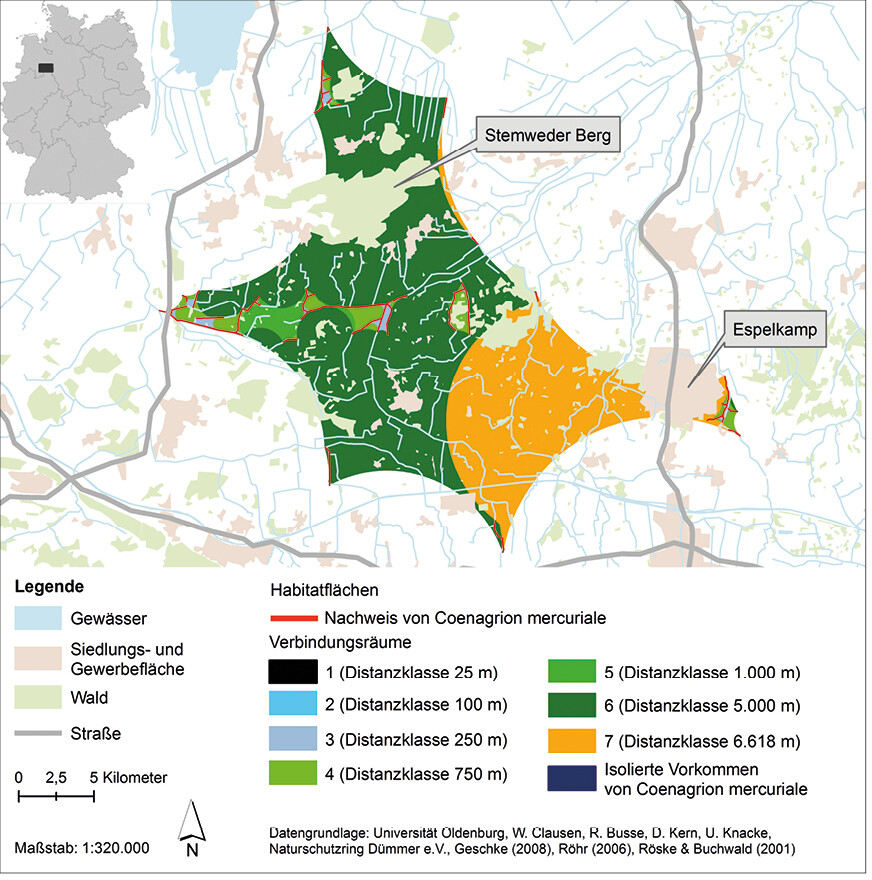
Die räumliche Habitatanalyse (Abb. 6) zeigt, dass bei der Distanzklasse 25 m nur zwischen den direkt aneinander angrenzenden Gewässerabschnitten ein Austausch von Individuen stattfindet. Auch bei der Distanzklasse 100 m findet ein Austausch nur zwischen direkt benachbarten Habitaten statt. Bei der Distanzklasse 250 m sind die Gewässerabschnitte der einzelnen Gebiete z. T. miteinander vernetzt, jedoch gilt dies nicht für alle Gewässer eines Gebiets. Dies ist bei der Distanzklasse 750 m erreicht; alle Gewässerabschnitte eines Gebiets sind miteinander vernetzt, mit Ausnahme der Abschnitte im Gebiet Sulingen (Abb. 6). Jedoch befinden sich z. B. im Gebiet Mehner Bruch Waldbereiche, die den Vernetzungsraum zerschneiden. Die Gebiete Tiefenriede und Ilwede mit Diekfluss sind ab der Distanzklasse 1 km miteinander verbunden. Alle anderen Gebiete sind weiterhin voneinander isoliert. Bei der Distanzklasse 5 km sind die Gebiete Tiefenriede, Ilwede mit Diekfluss, Mehner Bruch, Bohmte, Getmold, Pissing, Oppendorf und Oppenwehe miteinander vernetzt. Jedoch beinhaltet dieser Funktionsraum größere Gehölzbestände, Wald- und Siedlungs-/Gewerbeflächen, die als Barrieren dienen könnten, sowie den 181 m hohen bewaldeten Stemweder Berg, der den Funktionsraum im Norden durchteilt. Im Gebiet Sulingen stehen bei dieser Distanzklasse alle Gewässerabschnitte miteinander im Austausch, ebenso wie die Gewässerabschnitte in Neuenknick und Wiedensahl Flecken (Abb. 6 und 7). Bei der maximalen Wanderungsentfernung von über 6 km würde auch das Gebiet Espelkamp mit den vorher genannten Gebieten (Tiefenriede, Ilwede mit Diekfluss, Pissing usw.) in Verbindung stehen. Jedoch wird dieser Funktionsraum neben dem Stemweder Berg auch durch die Stadt Espelkamp zerschnitten (Abb. 7). Weiterhin isoliert sind die Vorkommen in Eistrup, Hütten, Loccum und Bockhop (Abb. 6).
Offen bleibt auch bei dieser Analyse, inwieweit die vorhandenen Fließgewässer und Gräben als Vernetzungskorridore in den Funktionsräumen und zwischen diesen wirken und der Stemweder Berg und die Stadt Espelkamp sowie die weiteren Wald- und Siedlungs-/Gewerbeflächen eine Barrierewirkung haben. Im Rahmen verschiedener Mark-recapture-Studien zum Ausbreitungsverhalten von C. mercuriale konnte nachgewiesen werden, dass die Art entlang von Gräben und Bächen sowie auch über Land (also über nicht-aquatische Landschaftsteile) wandert. Dabei stellen u. a. kleine gewässerbeschattende Gehölzbestände (bis ca. 50 m) keine unüberwindbaren Barrieren für einzelne Individuen dar.
Die Vernetzungsanalyse zeigt, dass im betrachteten Grenzraum zwischen Nordrhein-Westfalen und Niedersachsen bei einer maximalen Wanderungsdistanz von 6,6 km die Habitate teilweise miteinander verbunden sind und in diesen vermutlich eine Metapopulation (Tiefenriede, Ilwede mit Diekfluss, Pissing, Espelkamp) sowie mehrere einzelne isolierte Populationen (u. a. Sulingen oder Neuenknick) der Art existieren.
5 Schlussfolgerung
Aufbauend auf den Kenntnissen zum Vorkommen von A. viridis und S. aloides in der Hunte-Weser-Niederung und den Kenntnissen zum Vorkommen und zur Mobilität von C. mercuriale im Grenzraum zwischen Nordrhein-Westfalen und Niedersachsen konnte eine GIS-basierte Analyse der Habitatvernetzung für die beiden Arten durchgeführt werden. Diese zeigt, dass die einzelnen Habitate von A. viridis miteinander verbunden sind, jedoch für deren Vernetzung bei geringen Distanzklassen die Habitate durch Trittsteine (Krebsscheren-Bestände) ergänzt werden sollten. Eine Grundvoraussetzung für den Aufbau eines Habitatverbundsystems ist darüber hinaus die Erhaltung und Optimierung aller A. viridis- Habitate (Krebsscheren-Bestände) durch eine ökologische Grabenunterhaltung (Nagler&Müller2012).
Die einzelnen Habitate von C. mercuriale sind nur z. T. bei der größten Distanzklasse miteinander verbunden; für ein mögliches Habitatverbundsystem müssen v. a. zwischen den aktuell isolierten Populationen Trittsteine in Form von geeigneten Fließgewässerabschnitten geschaffen werden. Auch hier spielen für das Gelingen eines Habitatverbundsystems die Erhaltung und Optimierung der bestehenden Lebensräume durch eine an die Ansprüche der Art angepasste Gewässerunterhaltung (Serflinget al. 2004) eine zentrale Rolle. Für die beiden Habitatvernetzungsanalysen bleibt die Rolle von Fließgewässern und Gräben als Vernetzungskorridore offen, genauso wie die Barrierewirkung der verschiedenen Landschaftselemente (charakterisiert u. a. durch Größe und Art des Elements).
Aufbauend auf den Ergebnissen der hier vorgestellten räumlichen Habitatanalyse von A. viridis- und C. mercuriale- Vorkommen in Nordwest-Deutschland sollten regionale Habitatverbundsysteme mit Maßnahmen zur Erhaltung, Optimierung und Vernetzung der Lebensräume geplant und umgesetzt werden.
Dank
Die vorliegende Studie wurde im Projekt Artenhilfsprogramme für die FFH-Libellenarten Aeshna viridis, Coenagrion mercuriale und Coenagrion ornatum in NW-Deutschland wissenschaftliche Grundlagen und Maßnahmen zur Verbesserung der Habitatqualität und des Habitatverbundes, erstellt, gefördert durch die Deutsche Bundesstiftung Umwelt (DBU) (Az: 29355-33/2). Wir danken W. Clausen, R. Busse, D. Kern, U. Knacke und dem Naturschutzring Dümmer e.V. für die Bereitstellung von Funddaten.
Literatur
Aus Umfangsgründen steht das ausführliche Literaturverzeichnis unter www.nul-online.de (WebcodeNuL2231 ) zur Verfügung.
Fazit für die Praxis
Ein Habitatverbundsystem fürA. viridis undC. mercuriale sollte folgenden Maßnahmen beinhalten:
- Erhaltung und Optimierung der Lebensräume durch eine angepasste ökologische Gewässerunterhaltung,
- Erhaltung und Optimierung der Lebensräume durch eine extensive Bewirtschaftung der umgebenden landwirtschaftlichen Nutzflächen zur Verringerung von Nährstoffeinträgen,
- Vernetzung der Habitate vonA. viridis innerhalb eines Gewässersystems und zwischen zwei und mehreren Gewässersystemen durch Wiederansiedlung vonS. aloides in geeignete Gewässer,
- Erhaltung und Optimierung der Lebensräume (Gewässersystem) vonC. mercuriale durch Beseitigung beschattender Gehölze und Maßnahmen zur Sicherung der permanenten Wasserstandsführung,
- Vernetzung der Habitate vonC. mercuriale innerhalb eines Gewässersystems und zwischen zwei und mehreren Gewässersystemen durch Beseitigung von Barrieren (z. B. langen Gehölzbeständen) und durch die Schaffung geeigneter Fließgewässerabschnitte (z. B. Maßnahmen zur Wasserstandsführung, Erhöhung der Fließgeschwindigkeit).
Kontakt
Friederike Kastner arbeitet seit 2010 als wissenschaftliche Mitarbeiterin in Forschungsprojekten zu Ökologie und Schutz von Libellen in der Arbeitsgruppe Vegetationskunde und Naturschutz an der Carl von Ossietzky Universität Oldenburg. Studium der Diplom-Landschaftsökologie mit Schwerpunkt Ökologie und Naturschutz in Oldenburg. Forschungsschwerpunkte sind Ökologie von Libellen und Tagfaltern, Biotopverbund und Naturschutz.> friederike.kastner@uni-oldenburg.de
Prof. Dr. Rainer Buchwald hat seit 2005 die Professur für Vegetationskunde und Naturschutz an der Carl von Ossietzky Universität Oldenburg inne, zuvor seit 1997 die Professur für Terrestrische Ökologie und Naturschutz an der Hochschule Vechta. Studium, Promotion und Habilitation an der Albert-Ludwigs-Universität Freiburg. Forschungsschwerpunkte sind Grünland, Bioenergie (Hydrothermale Carbonisierung), Libellenökologie, Restitutionsökologie, Biotopverbund, Landwirtschaft & Naturschutz.> rainer.buchwald@uni-oldenburg.de
-
Abb. 1: Frisch geschlüpftes Individuum vonAeshna viridis (rechts) und besiedelterStratiotes aloides -Graben in der Hunte-Weser-Niederung (links).F. Kastner -
Abb. 2: Tandem vonCoenagrion mercuriale (rechts) und besiedeltes Gewässer in Nordrhein-Westfalen (links).F. Kastner -
Abb. 3: Vorkommen vonStratiotes aloides undAeshna viridis in der Hunte-Weser-Niederung zwischen Oldenburg (Niedersachsen) und Bremen.Von Friederike Kastner und Rainer Buchwald Eingereicht am 23. 07. 2018, angenommen am 31. 01. 2019 -
Abb. 4: Vorkommen vonCoenagrion mercuriale im Grenzraum zwischen Nordrhein-Westfalen und Niedersachsen.Von Friederike Kastner und Rainer Buchwald Eingereicht am 23. 07. 2018, angenommen am 31. 01. 2019 -
Abb. 5: Ergebnis der Vernetzungsanalyse zur Entwicklung eines Habitatverbundsystems fürAeshna viridis in der Hunte-Weser-Niederung zwischen Oldenburg (Niedersachsen) und Bremen. Grundlage für die Erstellung der Verbindungsräume sind die aktuellen Vorkommen vonStratiotes aloides im Projektgebiet. Verwendete Ausbreitungsdistanzen: 500 m, 2000 m, 4000 m, 9000 m und 14 000 m.Von Friederike Kastner und Rainer Buchwald Eingereicht am 23. 07. 2018, angenommen am 31. 01. 2019 -
Abb. 6: Ergebnis der Vernetzungsanalyse zur Entwicklung eines Habitatverbundsystems fürCoenagrion mercuriale im Grenzraum zwischen Nordrhein-Westfalen und Niedersachsen. Grundlage für die Erstellung der Verbindungsräume sind die Vorkommen vonCoenagrion mercuriale im Grenzraum zwischen Nordrhein-Westfalen und Niedersachsen sowie die gewonnenen Kenntnisse zum Ausbreitungsverhalten der Art. Verwendete Ausbreitungsdistanzen: 25 m, 100 m, 250 m, 750 m, 1000 m, 5000 m und 6618 m.Von Friederike Kastner und Rainer Buchwald Eingereicht am 23. 07. 2018, angenommen am 31. 01. 2019 -
Abb. 7: Ausschnitt der Vernetzungsanalyse zur Entwicklung eines Habitatverbundsystems fürCoenagrion mercuriale im Grenzraum zwischen Nordrhein-Westfalen und Niedersachsen. Grundlage für die Erstellung der Verbindungsräume sind die Vorkommen vonCoenagrion mercuriale im Grenzraum zwischen Nordrhein-Westfalen und Niedersachsen sowie die gewonnenen Kenntnisse zum Ausbreitungsverhalten der Art. Verwendete Ausbreitungsdistanzen: 25 m, 100 m, 250 m, 750 m, 1000 m, 5000 m und 6618 m.Von Friederike Kastner und Rainer Buchwald Eingereicht am 23. 07. 2018, angenommen am 31. 01. 2019 -
-
-
Friederike KastnerPrivat -
Prof. Dr. Rainer BuchwaldPrivat












Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.