Vom Auenwald zum Rot-Buchenwald?
Abstracts
Im NWR Riedried erfolgte 1986 eine Erst- und 2018 eine Wiederholungserfassung der Vegetation. Drei Waldgesellschaften wurden unterschieden und deren Veränderungen über den Untersuchungszeitraum von 32 Jahren beschrieben. In der im Gebiet vor allem randlich vorkommenden Allium-Fagus -Gesellschaft kam es als Folge des Eschentriebsterbens zur Abnahme der Baumschichtdeckung. Die Häufigkeit der Rot-Buche nahm sowohl in der Baum- als auch in der Strauchschicht deutlich zu. Auch in der Convallaria-Quercus -Gesellschaft ist die Baumschicht deutlich lückiger geworden, was hier vor allem auf den Rückgang der stark von Windwurf und Windbruch betroffenen Moor-Birken zurückzuführen ist. Lücken werden offenbar rasch von Berg-Ahorn und Rot-Buche geschlossen. Verjüngung der Stiel-Eiche ist trotz zahlreicher Altbäume nicht zu verzeichnen. Relativ geringe Veränderungen zeigen dagegen die Bestände der Solanum-Alnus -Gesellschaft, die vor allem auf ehemaligen Abtorfungsflächen stocken. Zwar wurden 2018 geringere Kronendeckungen als 1986 ermittelt, doch scheinen diese ein witterungs- oder krankheitsbedingtes Phänomen zu sein, da die Deckungen in Strauch- und Krautschicht eher rückläufig sind und vor allem einige lichtliebende Röhrichtarten, die 1986 noch häufig auftraten, 2018 nicht wiedergefunden wurden. Außerhalb der nassen Schwarz-Erlenstandorte deutet sich im Riedried insgesamt die Etablierung buchenreicher Bestände an. Mit einem Erhalt der noch vielfach als natürlich angesehenen Eichen-Hainbuchenwälder in diesem Bereich ist bei weiterer ungestörter Entwicklung, das heißt ohne aktive Förderung der Stiel-Eiche, jedoch nicht zu rechnen.
From floodplain forest to European beech forest? Development of vegetation in Riedried strict forest reserve and Nature Conservation Area (Palatinate Rheinaue) from 1986 to 2018
In Riedried strict forest nature reserve, a vegetation survey was conducted in 1986 and repeated in 2018. Three forest communities could be distinguished and their changes over the period of 32 years described. In the more marginal appearing Allium-Fagus-community, the tree layer coverage decreased as a consequence of ash dieback. The abundance of European beech increased distinctively in the tree layer as well as in the shrub layer. The tree layer of the Convallaria-Quercus-community also showed a considerable increase in gaps, which can be explained by the loss of downy birch, mainly due to windthrow. Gaps show clear rapid closing by sycamore and European beech. Despite a high number of older trees in the overstory, natural regeneration of pendunculate oak is missing. In contrast, the stands of the Solanum-Alnus-community, growing mainly on former peat diggings, showed relatively minor changes. However, a lower canopy closing was recorded in 2018 compared to 1986. But this seems to be more caused by the weather conditions or diseases, since the coverage of shrub and herb layers were declining and, most of all, light demanding species typical for reedbed communities, which occurred with high abundance in 1986, were not recorded in 2018. Except for the wet stands dominated by alder, the results indicate a development towards European beech-dominated stands in Riedried. The conservation of oak-hornbeam-forests is not expected; these are still considered in many cases as the natural vegetation on such sites undisturbed by development, i.e. without active facilitation of pendunculate oak.
- Veröffentlicht am

1 Einleitung
Die Auenlandschaften entlang der großen Flüsse wie des Rheins gehören in Mitteleuropa seit Langem zu den vom Menschen stark veränderten Lebensräumen (Ellenberg & Leuschner2010,Volk2006). Die azonalen Auenwälder haben durch Gewässerausbau, Eindeichung oder Stauregulierungen ihre ursprünglichen Standortbedingungen weitgehend verloren, wenn sie nicht sogar aufgrund der hohen Fruchtbarkeit ihrer Böden entwaldet und landwirtschaftlich – vor allem als Grünland – genutzt wurden. Dies gilt auch für das in einer Randsenke der pfälzischen Rheinaue gelegene „Riedried“, dessen Feuchtwälder in der Vergangenheit durch Entwässerungsmaßnahmen, Abtorfung und Kultivierung mit Phasen intensiver Niederwald- oder Wiesennutzung geprägt waren (Bartschet al. 1990,Wevell von Krüger1988). Bei der Ausweisung als Naturwaldreservat (NWR) 1979 und als Naturschutzgebiet (NSG) 1984 bildeten im Riedried Niedermoorreste mit Moorwäldern, aber auch eichenreiche Wälder sowie Wiesen und Röhrichte einen kleinflächig strukturierten Vegetationskomplex.
Die Aufgabe der früheren Nutzungen selbst sowie die durch die Nutzungsaufgabe einsetzenden Standortveränderungen führen in Wäldern in der Regel zu einer ausgeprägten Vegetationsdynamik (Ellenberg & Leuschner2010, Schmidt & Heinrichs2016). Nicht selten steht diese in Widerspruch zu statischen Zielen im Naturschutz, der oft bestimmte Lebensräume oder Artengruppen langfristig zu erhalten oder zu fördern versucht (Natura 2000, BNatSchG). Dementsprechend sind umfangreiche Kenntnisse über die dann einsetzenden dynamischen Prozesse in Ökosystemen notwendig, um sie aus der Sicht des Waldbaus und Naturschutzes angemessen bewerten zu können. Sie liefern damit auch eine wichtige Grundlage für Pflege- und Entwicklungspläne in Naturschutzgebieten. Naturwaldreservate bieten hier die Möglichkeit, die Sukzession nach Wegfall der Bewirtschaftung von Wäldern oder nach gravierenden Standortveränderungen zu untersuchen.
Hinsichtlich der derzeit im NWR Riedried vorkommenden Waldgesellschaften mit ihrer aktuell noch stark durch den Menschen beeinflussten Baumartenzusammensetzung gilt es abzuschätzen, inwiefern diese bereits der potenziell natürlichen Vegetation nahekommt oder ob langfristig ein Wechsel zu erwarten ist. Entscheidend dürfte dabei auch sein, inwieweit die in der Vergangenheit stattgefundenen Eingriffe wie Abtorfung und Entwässerung langfristig reversibel sind und damit eine gewisse Resilienz der ursprünglich vorhandenen naturnahen Vegetation angenommen werden darf. Zu diesem vielfältigen Prozess der Waldentwicklung gehört auch das Eschentriebsterben (Pautassoet al. 2013), welches seit einigen Jahren im NWR Riedried zum Absterben der zahlreichen Eschen im Altbestand führt.
Die 1986 durchgeführten Vegetationsaufnahmen vonWevell von Krüger (1988) undWienstroth(1988) bieten eine hervorragende Grundlage, um mit der Wiederholungserfassung im Jahr 2018 die Veränderungen der Vegetation nach mehr als 30 Jahren zu beschreiben, zu analysieren und zu bewerten. Dabei sollen vorrangig folgende Fragen beantwortet werden:
(1) Welche Trends der Vegetationsentwicklung zeichnen sich für die Wälder des NWR Riedried ab?
(2) Erlaubt die Zu- oder Abnahme bestimmter Pflanzenarten Rückschlüsse auf sich verändernde Standortfaktoren?
(3) Ist Nichtbewirtschaftung der Waldbestände im NWR Riedried zielführend für die als schützenswert angegebenen Lebensraumtypen?
2 Untersuchungsgebiet
Das NWR Riedried liegt linksrheinisch, etwa 12 km westlich vom auf der anderen Rheinseite gelegenen Karlsruhe und circa 2,5 km nordwestlich der Ortschaft Hagenbach, gehört zum Forstamt Pfälzer Rheinauen mit zwei Unterabteilungen (28a, 28b) im Forstrevier Rheinauen. Es befindet sich mit einer Höhe von 105–107 m ü. NN direkt unterhalb des Rheinhochufers in einer Randsenke der Rheinaue. Das Klima zeichnet sich mit einer Jahresmitteltemperatur von 10,3 °C (17 °C in der Vegetationsperiode) und einer mittleren Jahresniederschlagssumme von 700 mm (335 mm in der Vegetationsperiode) durch warme Sommer und milde Winter aus. Als Böden herrschen meso- bis eutrophe Niedermoore, Gleye und Pseudogleye vor (Bartschet al. 1990, Wevell von Krüger1988).
Die Niedermoore der Randsenken wurden im Vergleich zu anderen, meist fruchtbareren Bereichen der Rheinaue aufgrund des hier dauerhaft hoch anstehenden Grundwassers erst relativ spät kultiviert oder auch länger nur extensiv genutzt. Nach der ältesten vorhandenen Bestandesbeschreibung von 1837 wird für das Riedried ein „ziemlich geschlossener“ Bestand angegeben, dominiert von Erlenstockausschlägen und eingebrachten Eschenstangen im Unterstand, mit Eichen, Buchen, Hainbuchen, Aspen, Eschen, Erlen, Ulmen, Birken und Kiefern im lichten Oberstand (Bartschet al. 1990,Wevell von Krüger1988). Die Nutzung des Unterstands erfolgte damals im Stockausschlagbetrieb mit kurzer Umtriebszeit von circa sechs Jahren und diente lange Zeit der Faschinenerzeugung für die Rheinausbauten vor und während der Rheinkorrektur (1817–1883,Volk2006). Mit Fortschreiten der Rheinbegradigung nahm Ende des 19. Jahrhunderts die Bedeutung dieser Niederwälder stark ab. Gleichzeitig begann um die Jahrhundertwende eine intensive Torfnutzung im Riedried, die bis 1924 andauerte, als mit sinkenden Kohlepreisen der Torfstich nicht mehr rentabel war. Die größte Ausdehnung der Torfnutzungsfläche wird 1903 mit 17,3 ha angegeben, das heißt mit rund zwei Dritteln der Fläche des heutigen NSG Riedried. In der Zeit des Torfabbaus gab es auch eine zwischenzeitliche Nutzung als Grünland, indem die noch nicht abgetorften Flächen als Wiesen an Bauern der Umgebung verpachtet wurden. 1935 wird für die ehemaligen Torfstiche eine Dominanz von Erlenjungbeständen angegeben, die als dicht geschlossen und sehr wüchsig beschrieben werden. Vermutlich wurden in den 1920er- bis 1930er-Jahren am Rand des Hochgestades auch Kulturen mit Esche und Ahorn begründet. Es finden sich zu der Zeit wenige Altholzreste mit Birken und Erlen, aber auch Eiche, Esche, Buche und Ahorn sowie 8,8 ha Nichtholzbodenfläche. Der größte Teil wird als „Ödung“ bezeichnet, ein ursprünglich wohl auch zur Torfgewinnung vorgesehener Bereich, dominiert von Moorbirke mit geringer Wüchsigkeit. Umfangreiche Entwässerungsbauten mit Gräben und Dämmen zur Verbesserung der Bewirtschaftbarkeit der Bestände fanden im gesamten Untersuchungsgebiet von 1935 bis zum Kriegsbeginn 1939 statt. Eine letzte Öffnung der zahlreichen Entwässerungsgräben fand Anfang der 1970er-Jahre statt (Bartschet al. 1990,Wevell von Krüger1988).
In den Kriegsjahren unterblieben die im Betriebswerk von 1935 vorgesehenen Bestandespflegemaßnahmen. Auch der Forstwirtschaftsplan von 1965 lässt auf eine nur geringe Nutzung des Riedrieds in der Nachkriegszeit schließen. Die 1935 als Ödung bezeichnete Fläche hatte sich bis 1965 – wahrscheinlich durch natürliche Sukzession – mit circa 10–30-jährigen Birken, Pappeln, Erlen, Eschen, Ulmen und Kiefern bestockt. Ansonsten dominierten Erlen-, Eschen-Erlen- und Birkenbestände, die 1974 letztmals genutzt wurden. Bei der Ausweisung als NWR (1979) und als NSG (1984) entfielen von den insgesamt 25 ha noch circa 4 ha auf Wiesen- und Röhrichtflächen. 1995 erfolgte eine östliche Erweiterung des NWRs (nicht aber des NSG) auf eine Gesamtfläche von 45 ha, die heute auch Teil des FFH-Gebiets „Bienwaldschwemmfächer“ ist.
3 Methoden
Von den 1986 im älteren Bereich des NWRs angefertigten 69 Vegetationsaufnahmen konnten 64 Aufnahmeflächen anhand der in einer Karte eingezeichneten Aufnahmepunkte sowie den Angaben zum damaligen Bestandesaufbau als Quasi-Dauerflächen (Röderet al. 1996) relokalisiert und 2018 erneut aufgenommen werden (die nach floristischer Ähnlichkeit sortierten Gesamttabellen der Aufnahmen von 1986 und 2018 finden sich in Tab. A1 des Online-Supplements unter Webcode NuL2231 ). Davon entfallen sieben Aufnahmeflächen auf Standorte, die für einen Vergleich mit den Aufnahmen von 1986 aufgrund ihrer Lage und Struktur sowie von Störungseinflüssen für eine Darstellung der Waldentwicklung nicht geeignet erschienen.
Die Aufnahmeflächengröße beträgt aufgrund der kleinräumigen Heterogenität vieler Bestände in der Regel 100 m². Bei Aufnahmeflächen mit Frühjahrsgeophyten erfolgten Vegetationsaufnahmen sowohl im Frühjahr (April 2018, Zeitpunkt der beginnenden Blüte von Allium ursinum und Vollblüte von Anemone ranunculoides ) als auch im Sommer (Juli bis August 2018), bei den übrigen Flächen nur ein Mal im Sommer. Für die Aufnahmen 1986 liegen keine genauen Aufnahmezeitpunkte vor, es wurden jedoch ebenfalls Frühjahrs- und Sommeraufnahmen durchgeführt. Dabei wurde der Deckungsgrad der Vegetationsschichten (Baum-, Strauch-, Kraut- und Moosschicht) in Prozent geschätzt, für die einzelnen Arten in den Vegetationsschichten 1986 nach der Braun-Blanquet-Skala und 2018 ebenfalls direkt in Prozent (Dierschke1994). Für den Vergleich zwischen Erst- und Wiederholungsaufnahme wurden bei den einzelnen Arten die direkt in Prozent geschätzten Deckungsgrade der Aufnahmen 2018 zuerst in die Braun-Blanquet-Skalenwerte und dann in die beiDierschke(1994) dazu angegebenen mittleren Deckungsgrade transformiert. Die Nomenklatur der Gefäßpflanzen richtet sich nachWisskirchen & Haeupler(1998).
Für die über die pflanzensoziologische Auswertung (siehe Abschnitt 4.1) unterschiedenen drei Hauptgruppen beziehungsweise Waldgesellschaften wurden für den Vergleich der Aufnahmen von 1986 und 2018 die mittleren Artenzahlen sowie die mittleren Deckungsgrade sowohl der einzelnen Schichten als auch der einzelnen Arten berechnet und zur Kennzeichnung der Streuung jeweils der Standardfehler angegeben. Die Moose wurden dabei nicht berücksichtigt, da sie 1986 nicht oder nur teilweise aufgenommen wurden. Um Veränderungen ökologischer Bedingungen über den Untersuchungszeitraum aufzuzeigen, wurden außerdem die mittleren ungewichteten sowie nach Deckungsgrad gewichteten Zeigerwerte nachEllenberget al. (2001) für Licht, Stickstoff sowie die Bodenfeuchte für die Krautschicht berechnet. Für einen statistischen Vergleich zwischen der Erst- und Wiederholungserfassung wurden der gepaarte t -Test bei normalverteilten Daten und der Wilcoxon-Test bei fehlender Normalverteilung innerhalb einer Hauptgruppe verwendet. Die Prüfung auf Normalverteilung erfolgte mit dem Kolmogorov-Smirnov-Test. Als Signifikanzniveau wurde einheitlich p 0,05 akzeptiert. Die statistischen Berechnungen wurden mit dem Programm SigmaPlot 11.0 (Systat Software) durchgeführt, die Berechnung der mittleren ökologischen Zeigerwerte und Diversitätsmaße mit dem datenbankbasierten Auswertungsprogramm FRIDOLINO (Parth & Fischer2009).
4 Ergebnisse
4.1 Waldgesellschaften im Riedried
Die Waldbestände im Riedried ließen sich hinsichtlich der Zusammensetzung der Baumarten wie auch der begleitenden Krautschicht nach den Aufnahmen des Jahres 2018 grob in drei Gesellschaften gliedern. Dabei können die Fagus - und Quercus -Gesellschaften der Klasse der Buchen-Mischwälder (Querco-Fagetea) zugeordnet werden, während die Alnus -Gesellschaften standörtlich wie soziologisch am Übergang von Au- zu Bruchwäldern (Alno-Ulmion beziehungsweise Alnetea glutinosae) stehen. Da in der vorliegenden Untersuchung nicht die pflanzensoziologische Stellung oder synsystematische Zuordnung der Gesellschaften des NWR Riedried im Mittelpunkt steht (was einen umfassenderen Vergleich erfordern würde), sondern der zeitliche Vergleich zwischen Erst- und Wiederholungsaufnahme, werden hier rangneutrale Bezeichnungen verwendet, die die lokale Vergesellschaftung klar zum Ausdruck bringen. Für die Benennungen wurden jeweils eine wichtige, differenzierende Baumart und eine charakteristische Art der Krautschicht verwendet.
Im Frühjahr gehören die dichten Alliumursinum -Bestände der Allium-Fagus- Gesellschaft zu den auffälligsten Erscheinungen im NWR Riedried (Abb. 1). Sie ziehen sich im Westen und Nordwesten bandartig am Fuß der Niederterrasse entlang und bilden im neuen Teil des Untersuchungsgebietes auch flächigere Bestände aus. Allium ursinum ist regelmäßig von Rot-Buchen ( Fagus sylvatica ) überstanden, die meist die erste und/oder zweite Baumschicht dominieren. Manchmal gelangen auch Gewöhnliche Esche ( Fraxinus excelsior ) oder Berg-Ahorn ( Acer pseudoplatanus ), in wenigen Fällen Stiel-Eichen ( Quercus robur ) zur Dominanz. In kleineren Mengen ist die Schwarz-Erle ( Alnus glutinosa ) beigemischt. Neben dem oft dominanten A. ursinum ist die stete Beteiligung weiterer Geophyten wie Ranunculus ficaria, Anemone nemorosa oder A. ranunculoides sowie typischer Fagetalia-Arten wie Viola reichenbachiana kennzeichnend.
In weiten Teilen des NWR Riedried herrschen Waldbestände vor, die auf den ersten Blick sehr inhomogen aufgebaut erscheinen. Auffällig in dieser Convallaria-Quercus -Gesellschaft sind der hohe Anteil alter Stiel-Eichen sowie stattlicher Moor-Birken ( Betula pubescens ) und die fast durchweg gut entwickelte Strauchschicht. Die Moor-Birken wie auch die ehemals wohl häufigeren Gewöhnlichen Eschen sind stark von Windwurf und Windbruch betroffen. Dadurch wechseln kleinräumig dicht bestockte Partien mit fast baumlosen Lichtungen, was ein insgesamt „struppiges“ Bild erzeugt (Abb. 2). Über die Krautschicht sind diese Stiel-Eichenbestände nur sehr schwach charakterisiert. Lediglich Convallaria majalis zeigt einen deutlichen Verbreitungsschwerpunkt in dieser Gesellschaft. Ansonsten ist eine Mischung aus weit verbreiteten Fagetalia-Arten ( Fagus sylvatica , Melica uniflora , Sanicula europaea ) und Auwaldarten beziehungsweise Feuchtezeigern ( Prunus padus , Circaea lutetiana , Stachys sylvatica ) typisch.
Große Flächen des Untersuchungsgebietes sind mit sehr homogen aufgebauten, meist jüngeren Schwarz-Erlenbeständen der Solanum-Alnus -Gesellschaft bestockt. In der Regel beherrscht die Schwarz-Erle allein die Baumschicht und tritt auch in allen weiteren Schichten auf. Lediglich einzelne Gewöhnliche Eschen sind regelmäßig beigemischt. In der meist gut ausgebildeten Strauchschicht herrscht die Gewöhnliche Traubenkirsche ( Prunus padus ) vor (Abb. 3).
4.2 Vergleich 1986 und 2018
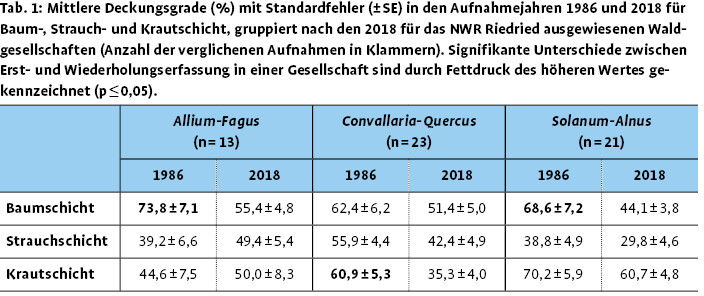
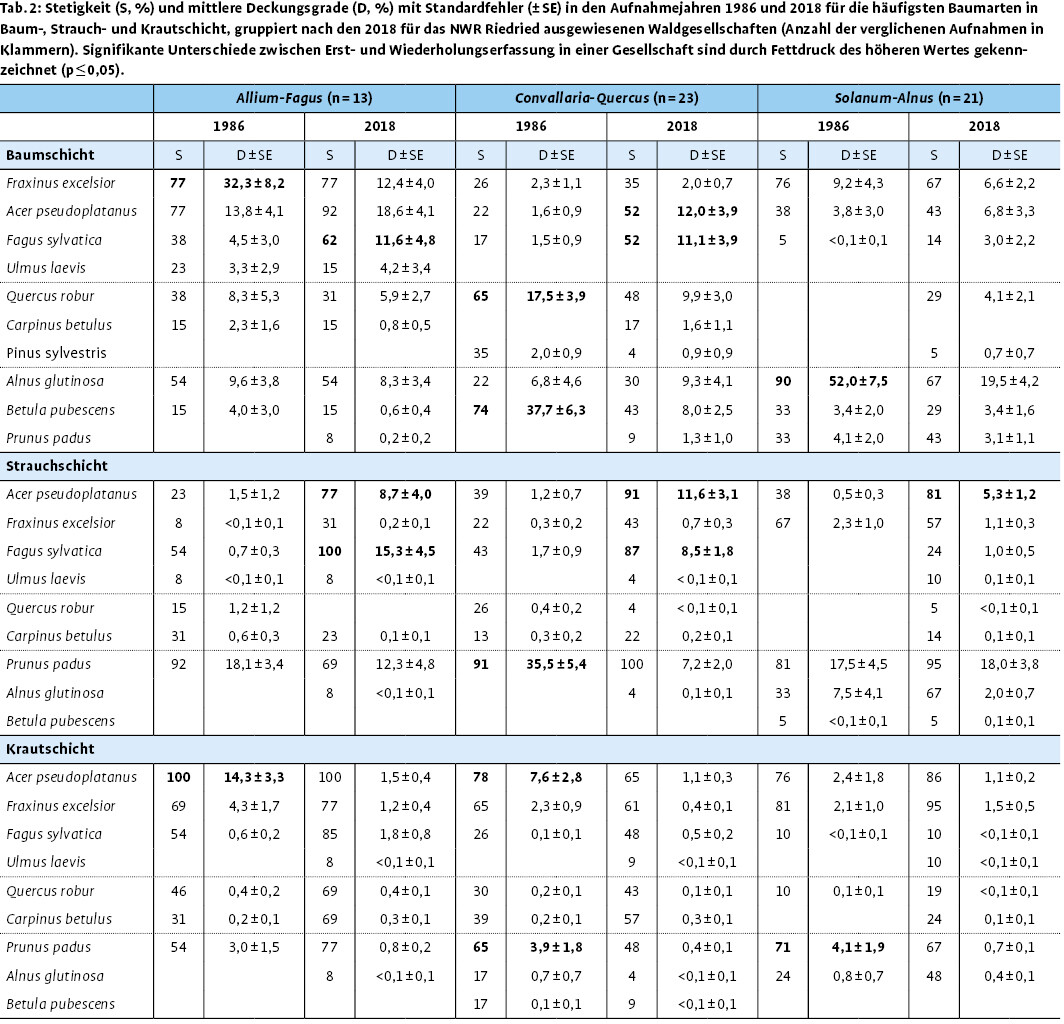
Die Entwicklung der Deckungsgrade der einzelnen Bestandesschichten bestätigt das bei den Aufnahmen im Gebiet wahrgenommene Bild, dass viele Bestände inzwischen stark aufgelichtet sind (Tab. 1). Insbesondere durch den Ausfall von Gewöhnlicher Esche und Moor-Birke ist der mittlere Deckungsgrad der Baumschicht deutlich zurückgegangen, wirkt sich aber in den drei Waldgesellschaften unterschiedlich aus (Tab. 2). So ist in der Allium-Fagus -Gesellschaft der Deckungsgrad der Baumschicht von 74 % im Jahr 1986 auf 55 % im Jahr 2018 im Mittel signifikant gesunken. Dies ist hier vor allem auf den Ausfall der Gewöhnlichen Esche zurückzuführen. Gleichzeitig profitierte die Strauchschicht mit einem mittleren Deckungsgrad von rund 50 % im Jahr 2018 von den günstigeren Lichtbedingungen. Der Deckungsgrad der Krautschicht von 45 % im Jahr 1986 beziehungsweise von 50 % im Jahr 2018 in der Allium-Fagus -Gesellschaft beruht auf dem hohen Anteil an Frühjahrsgeophyten, die im Vergleich zu 1986 deutlich zugenommen haben. Die der Convallaria-Quercus-Gesellschaft zugeordneten Aufnahmen zeigten nur einen geringen Rückgang der Baumschichtdeckung um knapp 10 %. Hier haben vor allem die Moor-Birke und auch die Stiel-Eiche deutlich abgenommen, während Berg-Ahorn und Rot-Buche neu in die Baumschicht einwuchsen. Durch die Zunahme von Rot-Buche und Berg-Ahorn auch in der Strauchschicht erhöhte sich der gemeinsame Deckungsgrad von Baum- und Strauchschicht. Gleichzeitig verringerte sich der Deckungsgrad der Krautschicht signifikant, wobei Frühblüher in der Convallaria-Quercus -Gesellschaft eine Zunahme zeigten. Der signifikante Rückgang der Baumschichtdeckung in der Solanum-Alnus -Gesellschaft betraf hauptsächlich die Schwarz-Erle, die 2018 aufgrund der extremen Trockenheit, möglicherweise auch durch Phytophthora -Befall nur sehr schütter belaubt war.
Die mittlere Gesamtartenzahl im Jahr 2018 bewegt sich zwischen 28 Arten pro Aufnahmefläche in der Solanum-Alnus -Gesellschaft und rund 30 Arten pro Aufnahmefläche in der Allium-Fagus - und der Convallaria-Quercus -Gesellschaft (Abb. 4). Sowohl die mittleren Gesamtartenzahlen als auch die Gesamtartenzahlen der Krautschicht zeigen keine signifikanten Veränderungen gegenüber 1986. Dagegen ist in allen Vegetationseinheiten die Artenzahl der Strauchschicht signifikant angestiegen. In der Convallaria-Quercus -Gesellschaft zeigte sich zudem ein signifikanter Anstieg der Artenzahl der Baumschicht. Moor-Birke, Wald-Kiefer ( Pinus sylvestris ) und Stiel-Eiche sind hier zwar deutlich in Deckungsgrad und Stetigkeit zurückgegangen, jedoch noch nicht vollständig ausgefallen. Zusammen mit dem Einwachsen von Berg-Ahorn, Rot-Buche und Hainbuche in die Baumschicht erklärt sich so die Zunahme der Artenzahl (Tab. 2). In der Strauchschicht sind ebenfalls Berg-Ahorn und Rot-Buche 2018 deutlich häufiger vertreten als bei der Erstaufnahme 1986.
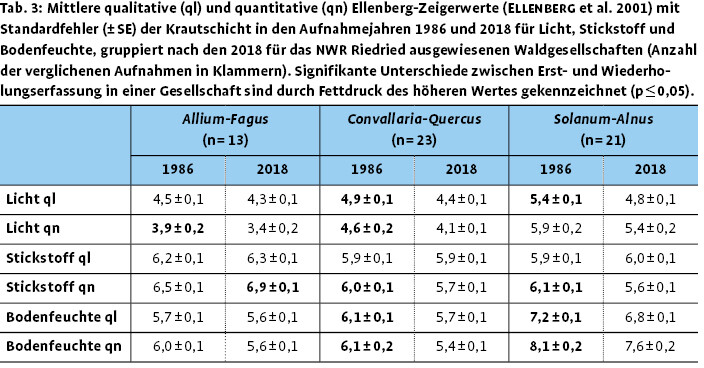
Hinsichtlich der ökologischen Zeigerwerte der Krautschicht zeigt sich für die drei Waldgesellschaften im Mittel ein leichter, teils signifikanter Rückgang der mittleren Lichtzahl (Tab. 3). Dies korrespondiert nicht mit der Abnahme des Deckungsgrads der Baumschicht, ist jedoch in der Summe mit der Zunahme im Deckungsgrad der Strauchschicht erklärbar. Die lichtesten Bedingungen weisen noch die Bestände der Solanum-Alnus -Gesellschaft auf. Die mittlere Feuchtezahl zeigt für die Convallaria-Quercus - und die Solanum-Alnus -Gesellschaft einen signifikanten Rückgang im Vergleich von 1986 zu 2018. Dies korreliert mit dem Rückgang beziehungsweise Ausfall zahlreicher lichtliebender Arten, die gleichzeitig auch hohe Zeigerwerte für Bodenfeuchte aufweisen. So waren 1986 mit Berula erecta, Cardamine amara, Euphorbia palustris, Glyceria fluitans, Phragmites australis und Scrophularia umbrosa noch typische Arten der Röhrichte und Seggenrieder in der Solanum-Alnus -Gesellschaft vorhanden, die 2018 völlig fehlten. Die mittlere gewichtete Stickstoffzahl nahm in der Allium-Fagus -Gesellschaft signifikant zu, in den anderen beiden Gesellschaften ab. In der Allium-Fagus -Gesellschaft geht dies vor allem auf Arten wie Allium ursinum, aber auch Urtica dioica und Alliaria petiolata zurück, die Zeiger für stickstoffreiche Standorte sind. Dagegen hat unter anderem Urtica dioica in den anderen beiden Gesellschaften im Deckungsgrad abgenommen.
5 Diskussion
In ehemals bewirtschaften Wäldern kommt es nach Einstellung der forstlichen Nutzung häufig zu einer Zunahme des Bestandesschlussgrades und damit zu einem Rückgang der Lichtverfügbarkeit am Waldboden (unter anderemHeinrichs&Schmidt2013,Meyeret al. 1998, 2015). Damit einhergehend nehmen lichtbedürftige und magerkeitszeigende Arten sowie Störzeiger ab, während sich gleichzeitig stickstoffliebende, konkurrenzstärkere Arten ausbreiten (unter anderemDittmannet al. 2018, Dölleet al. 2013, Heinrichset al. 2012, Mölderet al. 2014, Schmidtet al. 2019, Schmidt & Heinrichs2016). Oftmals kommt es insgesamt zu einer Abnahme der Artenzahlen bei den Gefäßpflanzen (Dierschke2013,Heinrichset al. 2011, ysik2008,Pailletet al. 2010,Schmidt2005).
Dies trifft bisher für das NWR Riedried nicht zu. Insbesondere in der Allium-Fagus - und der Convallaria-Quercus -Gesellschaft sind die Bestände durch eine große Dynamik und ein kleinflächig engverzahntes Nebeneinander an unterschiedlichen Bestandesentwicklungsphasen gekennzeichnet. Seit der Ersterfassung haben sich durch das altersbedingte Ausscheiden der Moor-Birke, dem durch das Eschentriebsterben verursachten Ausfall der Gewöhnlichen Esche und mit dem beginnenden Rückgang der Stiel-Eiche sowie der Zunahme der Rot-Buche die Wuchsbedingungen am Waldboden stark verändert.Bartschet al. (1990) haben diese Waldgesellschaften, die zwar außerhalb der für die Randsenken der Rheinaue typischen Niedermoore liegen, jedoch durch hoch anstehendes Grundwasser mit engem Kontakt zu den angrenzenden Moorböden geprägt sind, dem Stellario-Carpinetum zugeordnet und – wie lange Zeit üblich – als natürlich eingestuft, da unter diesen Standortbedingungen die Stiel-Eiche gegenüber der Rot-Buche als überlegen gilt. Durch ihre hohe Konkurrenzkraft, ihre stark abschattende Krone sowie ihre hohe Schattentoleranz in der Jugend wird die Rot-Buche aber auch hier langfristig dominieren und die Stiel-Eiche verdrängen (Ellenberg & Leuschner2010). Nichtstun wird selbst bei der hohen Störungsdynamik, wie sie im NWR Riedried mit zahlreichen, auch größeren Lücken und damit einem hohen Lichtangebot aktuell vorhanden ist, nicht ausreichen, um die Stiel-Eiche langfristig zu erhalten (Heinrichs & Schmidt2013).
Dies entspricht der Entwicklung im NWR „Pretzetzer Landwehr“ im niedersächsischen Elbeurstromtal, dessen stark von Stau- und Grundwasser beeinflusste moorige Böden von der Rot-Buche keineswegs gemieden werden. Bei fehlender Bewirtschaftung konnte diese sich sogar zur konkurrenzstärksten Baumart entwickeln (Schmidt2000). Ein Erhalt des Stellario-Carpinetums mit einem hohen Eichen-Anteil, wie er als schutzwürdiger Lebensraumtyp für das NSG Riedried gefordert wird, ist nur über entsprechende, die Stiel-Eiche fördernde waldbauliche Maßnahmen möglich.
Derzeit halten sich in der Allium-Fagus - und der Convallaria-Quercus -Gesellschaft immer noch zahlreiche weitere Mischbaumarten, von denen aber nur der Berg-Ahorn ein erfolgreiches Aufwachsen in die Baumschicht zeigt. Ob dies jedoch von Dauer sein wird, kann nur ein langfristiges Monitoring der Bestände im NWR Riedried aufzeigen. Auf lange Sicht ist mit Zunahme des Rot-Buchenanteils eine Abnahme der Diversität an Pflanzenarten in den unteren Vegetationsschichten zu erwarten. Die derzeit sehr hohe Dynamik und Heterogenität der Bestände sorgt aktuell noch für eine vergleichsweise hohe Artenvielfalt, vor allem durch zahlreiche Saumarten, die die nach wie vor recht lichten Bedingungen im NWR Riedried besonders kennzeichnen. Es ist jedoch zu vermuten, dass aufgrund der Zunahme der stark abschattenden Rot-Buche weitere lichtbedürftige Arten zurückgehen oder sogar ganz verschwinden werden.
Im Vergleich zu den buchen- und eichenreichen Waldgesellschaften dürften die Schwarz-Erlenbestände einigermaßen stabil bleiben. Die in der Baumschicht dominante Schwarz-Erle tritt auch in den unteren Vegetationsschichten regelmäßig auf und sollte bei Störungen (etwa Erlensterben durch Phytophthora ,Hartmann1995) die Verjüngung sichern. Für die Gewöhnliche Esche, die aktuell in der Baumschicht bereits eine geringere Rolle spielt als in den 1980er-Jahren, zeichnet sich im Zuge des Eschentriebsterbens ein weiterer Rückgang ab. Andere Baumarten spielen jetzt und wegen fehlender Verjüngung wohl auch zukünftig kaum eine Rolle in der Solanum-Alnus -Gesellschaft.
Typisch für die in den Randsenken der Rheinaue gelegenen Wälder sind die hier vorherrschenden Niedermoortorfe als klassische Standorte für Erlenbruch-Wälder (Carici-Alnetum) (Döring-Mederake1991, Mast1999). Solche wären unter natürlichen Bedingungen auch im NWR Riedried zu erwarten, wenn hier nicht zwischen etwa 1880 und 1925 intensiv Torfabbau erfolgt wäre. Die verbliebenen Torfauflagen sind meist unter 1 m mächtig, zum größten Teil gut zersetzt oder mit Mineralboden vermengt. Das dürfte auch die Ursache für das aktuelle Fehlen des Carici-Alnetums sein. Im Untersuchungszeitraum hat auf den ehemaligen Abtorfungsflächen die Vernässung jedoch wieder zugenommen, vermutlich, weil die zahlreichen Entwässerungsgräben ihre Funktion nicht mehr erfüllen (Abb. 5). Dies könnte mittelfristig eine Zunahme der Nässezeiger bewirken und langfristig wieder zur Entwicklung „echter“ Erlenbrücher führen. Dazu müssen jedoch nicht nur die Wasserstände hoch bleiben, sondern muss sich auch neuer Niedermoortorf aufbauen, wofür allerdings Zeiträume von mehreren hundert Jahren zu erwarten sind.
Literatur
- Bartsch, N., Wevell von Krüger, A., Wienstroth, A. (1990): Das Naturschutzgebiet Riedried im Staatlichen Forstamt Kandel (Rheinland-Pfalz). Mitt. Landesforstverwaltung Rheinland-Pfalz 8, 1-66.
- Dierschke, H.(1994): Pflanzensoziologie: Grundlagen und Methoden. Ulmer, Stuttgart.
- – (2013):Konstanz und Dynamik in einem artenreichen Kalkbuchenwald. Veränderungen in einem Großtransekt 1981–2011. Tuexenia 33, 49-92.
- Dittmann, T., Heinken, T., Schmidt, M.(2018): Die Wälder von Magdeburgerforth (Fläming, Sachsen-Anhalt) – eine Wiederholungsuntersuchung nach sechs Jahrzehnten. Tuexenia 38, 11-42.
- Dölle, M., Heinrichs, S., Schmidt, W., Schulte, U.(2013): Zwischen Anspruch und Wirklichkeit – die Entwicklung der Naturwaldzelle „Am Sandweg“, ein Eichen-Hainbuchenwald in einem FFH-Gebiet der Niederrheinischen Bucht. Archiv für Forstwesen und Landschaftsökologie 6, 1-12.
- Döring-Mederake, U. (1991): Feuchtwälder im nordwestdeutschen Tiefland; Gliederung – Ökologie – Schutz. Scripta Geobotanica 19, 1-122.
- Ellenberg, H., Leuschner, C.(2010): Vegetation Mitteleuropas mit den Alpen in ökologischer, dynamischer und historischer Sicht. 6. Aufl. Ulmer, Stuttgart.
- –, Weber, H.E., Düll, R., Wirth, V., Werner, W.(2001): Zeigerwerte von Pflanzen in Mitteleuropa. Scripta Geobotanica 18, 9-166.
- Hartmann, G. (1995): Wurzelhalsfäule der Schwarzerle (Alnus glutinosa ) – eine bisher unbekannte Pilzkrankheit durchPhytophthora cambivora . Forst und Holz 50, 555–557.
- Heinrichs, S., Schmidt,W. (2013): Windwurf und Eisbruch im Buchenwald: Eine Chance für Eiche und andere Baumarten? Ergebnisse aus vier Naturwaldreservaten. Forstarchiv 84, 181-197.
- –,Schulte, U.,Schmidt, W. (2011): Veränderung der Buchenwaldvegetation durch Klimawandel? – Ergebnisse aus ausgewählten Naturwaldzellen Nordrhein-Westfalens. Forstarchiv 82, 48-61.
- –,Winterhoff, W.,Schmidt, W. (2012): Vegetation dynamics of beech forests on limestone in central Germany over half a century – effects of climate change, forest management, eutrophication or game browsing? Biodiversity & Ecology 4, 49-61.
- ysikM. (2008): Ten years of change in ground-layer vegetation of European beech forest in the protected area (Ojców National Park, South Poland). Polish Journal of Ecology 56, 17-31.
- Mast, R. (1999): Vegetationsökologische Untersuchung der Feuchtwald-Gesellschaften im niedersächsischen Bergland – mit einem Beitrag zur Gliederung der Au-, Bruch- und Moorwälder in Mitteleuropa. Archiv naturwissenschaftliche Dissertationen 8, 1-307.
- Meyer, P.,Schulte, U.,Balcar, P.,Kölbel, M. (1998): Entwicklung der Baumarten- und Strukturdiversität in Buchen-Naturwaldreservaten. Beispiele aus Niedersachsen, Nordrhein-Westfalen, Rheinland-Pfalz und Bayern. NUA-Seminarbericht 4, 40-53.
- –,Lorenz, K.,Mölder, A.,Steffens, R.,Schmidt, W.,Kompa, T.,Wevell v. Krüger,A. (2015): Naturwälder in Niedersachsen – Schutz und Forschung, Band 2. Nordwestdeutsche Forstl. Versuchsanstalt, Göttingen (Hrsg.).
- Mölder, A., Streit, M., Schmidt, W. (2014): When beech strikes back: How strict nature conservation reduces herb-layer diversity and productivity in Central European deciduous forests. Forest Ecology and Management 319, 51-61.
- Paillet, Y., Bergès, L., Hjältén, J., Ódor, P., Avon, C., Bernhardt-Römermann, M., Bijlsma, R.-J., De Bruyn, L., Fuhr, M., Grandin, U., Kanka, R., Lundin, L., Luque, S. Magura, R., Matesanz, S., Mézáros, I., Sebastià, M.-T., Schmidt, W., Standovár, T., TÓthmérész, B., Uotila, A., Valladares, F., Vellak, K., Virtanen, R.(2010): Biodiversity differences between managed and unmanaged forests: meta-analysis of species richness in Europe. Conservation Biology 24, 101-112.
- Parth, A., Fischer, C. (2009): Fridolino – eine Datenbank zur Berechnung vegetationsökologischer Kenngrößen. Forstarchiv 80, 236-240.
- Pautasso, M., Aas, G., Queloz, V., Holdenrieder, O.2013: European ash (Fraxinus excelsior) dieback – A conservation biology challenge. Biological Conservation 158, 37-49.
- Röder, H., Fischer, A., Klöck, W.(1996): Waldentwicklung auf Quasi-Dauerflächen im Luzulo-Fagetum der Buntsandsteinrhön (Forstamt Mittelsinn) zwischen 1950 und 1990. Forstwissenschaftliches Centralblatt 115, 321-335.
- Schmidt, W. (2000): Eiche, Hainbuche oder Rotbuche? – Zur Vegetation und Baumartenzusammensetzung von stau- und grundwasserbeeinflußten Wäldern des nordwestdeutschen Tieflandes. Ergebnisse aus den Naturwäldern Hasbruch und Pretzetzer Landwehr. Tuexenia 20, 21-43.
- –(2005): Herb layer species as indicators of biodiversity of managed and unmanaged beech forests. Forest Snow and Landscape Research 79, 111-125.
- –,Dölle, M.,Heinrichs, S.,Balcar, P. (2019): Gebietsfremde Gefäßpflanzen in Naturwaldreservaten von Rheinland-Pfalz. Braunschweiger Geobotanische Arbeiten 13, 141-170.
- –,Heinrichs, S. (2016): Struktur und Artenreichtum der Vegetation in Naturwaldreservaten von Rheinland-Pfalz – Einfluss von Schalenwild und früherer Bewirtschaftung. Forstarchiv 87, 182-193.
- Volk, H. (2006): Landschaftsgeschichte und Natürlichkeit der Baumarten in der Rheinaue. Waldschutzgebiete Baden-Württemberg 10: 159-167.
- Wevell von Krüger, A. (1988): Das Naturschutzgebiet „Riedried“ in seiner geschichtlichen und standörtlichen Abhängigkeit. Unveröffentl. Diplom-Arb. Forstl. Fak. Univ. Göttingen.
- Wienstroth, A. 1988: Die Pflanzengesellschaften im Naturschutzgebiet Riedried und seine Stellung im Rahmen der Auenwälder am Oberrhein. Unveröffentl. Diplom-Arb. Forstl. Fak. Univ. Göttingen.
- Wisskirchen, R., Haeupler, H.(1998): Standardliste der Farn- und Blütenpflanzen Deutschlands. Ulmer, Stuttgart.
Fazit für die Praxis
- Die Vegetationsdynamik in Wäldern nach Einstellung der Bewirtschaftung ist in hohem Maße abhängig von den Ausgangsbeständen und damit der ehemaligen Nutzung. Beide Aspekte sind bei der Prognose der zukünftigen Waldentwicklung zu berücksichtigen.
- Azonale Auenwälder in Mitteleuropa haben durch Gewässerausbau, Eindeichung oder Stauregulierung ihre ursprünglichen Standortsbedingungen irreversibel verloren.
- Selbst auf vielen vermeintlich „buchenfeindlichen“ Auwaldstandorten ist daher unter ungestörten Bedingungen langfristig mit der Etablierung buchendominierter Waldbestände zu rechnen.
- Sollen eichenreiche Wälder als schutzwürdige Lebensraumtypen erhalten bleiben, so ist dieses Ziel nur über geeignete waldbauliche Maßnahmen, nicht aber durch einen Totalschutz zu erreichen.
- Die Etablierung „echter“ Erlenbruchwälder ist auf einst abgetorften Flächen nur bei dauerhaft hohen Wasserständen und auch dann erst nach sehr langer Zeit (erneute Bildung von Niedermoortorf) zu erwarten.
Kontakt
Dr. Michaela Dölle studierte und promovierte an der Fakultät für Forstwissenschaften und Waldökologie der Universität Göttingen und ist dort seit 2010 als Lehrkraft im Bereich Vegetationskunde und Waldökologie tätig. Schwerpunkte ihrer Arbeit sind die Untersuchung langfristiger Veränderungen der Vegetation vor allem in Naturwäldern sowie des Einflusses der Nutzungsgeschichte auf Waldökosysteme.
Prof. i. R. Dr. Wolfgang Schmidt promovierte und habilitierte sich am Systematisch-Geobotanischen Institut der Universität Göttingen. Ab 1994 Professur für Forstliche Ökosystemwissenschaften an der dortigen Fakultät für Forstwissenschaften und Waldökologie. Seit seiner Pensionierung beschäftigt vor allem mit den langfristigen Veränderungen der Waldbodenvegetation unter dem Einfluss des Nutzungs- und Umweltwandels.
Dr. Bernd Gehlken studierte und promovierte an der Universität Kassel (Landschaftsplanung) und arbeitet seit 2010 als Lehrkraft für Vegetationskunde und Naturschutz an der Fakultät für Forstwissenschaften und Waldökologie der Universität Göttingen. Schwerpunkte seiner Arbeit sind die Verknüpfung von Vegetationskunde und Landnutzungsgeschichte, sowie Fragen der pflanzensoziologischen Systematik.
David Vollmuth (M.Sc.) , wissenschaftlicher Mitarbeiter und Doktorand an der Abteilung Naturschutz und Landespflege der Fakultät für Forstwissenschaften an der Universität Göttingen.
Dr. Patricia Balcar, Leiterin des Forschungsbereiches Ökologische Waldentwicklung, Forschungsanstalt für Waldökologie und Forstwirtschaft Rheinland-Pfalz in Trippstadt.
-
Abb. 1: Bärlauch-Dominanz mit weiteren Frühjahrsgeophyten (unter anderemAnemone ranunculoides ) am Fuß der Niederterrasse in derAlliumursinum-Fagus-sylvatica -Gesellschaft.M. Dölle, 18. April 2018 -
Abb. 2: Blick in eine typische Bestandeslücke nach dem Absterben von Eschen in derConvallaria-majalis-Quercus-robur -Gesellschaft.B. Gehlken, 12. Juli 2018 -
Abb. 3: VonCarex acutiformis dominierteSolanum-dulcamara-Alnus-glutinosa -Gesellschaft mitPrunus padus blühend im Vordergrund. M. Dölle, 18. April 2018 -
Abb. 4: Boxplots der Artenzahlen je Probefläche (meist 100 m²) von Baum- (BS), Strauch- (Str), Krautschicht (KS) sowie gesamt (Ges, ohne Moosschicht) in den Aufnahmejahren 1986 und 2018, unterteilt nach den 2018 für das NWR Riedried ausgewiesenen Waldgesellschaften. Signifikante Unterschiede zwischen den Untersuchungsjahren innerhalb einer Schicht und Waldgesellschaft sind durch * gekennzeichnet (p 0,05).Dölle et al -
Abb. 5: Stark vernässter und aufgelichteter Bereich in derSolanum dulcamara -Alnus glutinosa -Gesellschaft des NWR Riedried im Juli 2018.M. Dölle -
-
-
-
-
-













Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.