Der Libellen-Schmetterlingshaft in Südwestdeutschland
Abstracts
Magere Trockenstandorte sind wichtige Lebensräume thermophiler Insekten, in Mitteleuropa aber durch Nutzungsaufgabe und Sukzession gefährdet. Die vorliegende Arbeit untersucht im Regierungsbezirk Tübingen Grundlagen für den Schutz und die Pflege solcher Habitate am Beispiel des Libellen-Schmetterlingshafts(Libelloides coccajus) , einer Leitart rohbodenreicher Trockenrasen und Schutthalden. 32 bekannte Fundstellen und 24 Potenzialgebiete wurden 2020 auf Vorkommen von Imagines und Eigelegen beprobt sowie die Habitatansprüche bezüglich makro- und mikrostruktureller Parameter charakterisiert.
Die nur 20 bestätigten Vorkommen deuten einen Bestands- und Arealrückgang an. Besiedelt wurden Wacholderheiden und Halbtrockenrasen mit Offenbodenstellen, Trockenmauern, Abbruchkanten oder Steinbrüche, aber auch Schutthalden in lichten Wäldern. Eigelege wurden fast ausschließlich über Roh- und Offenboden gefunden. Sie wurden bevorzugt in Südexposition an dürre, blattlose Stängel abgelegt, die waagerecht über voll besonnte, vegetationsarme Oberflächen geneigt waren.
Die Studie verdeutlicht das Gefährdungspotenzial durch mangelnde Nutzung und der daraus resultierenden Sukzession, die zu Lebensraumverlust sowie verstärkter Beschattung und Isolation der verbliebenen Habitate führt. Empfohlen werden eine intensive Gehölzpflege in Steillagen und an Schutthängen sowie die gezielte Schaffung von Roh- und Offenbodenstellen. Extensive Beweidung oder Mahd können die Habitate des Libellen-Schmetterlingshaft langfristig offenhalten, sofern sie durch regelmäßige Gehölzentnahmen ergänzt werden. The owly sulphur in southwestern Germany Habitat use, threats, and protection
Dry grasslands are core habitats of many thermophilic insects, but threatened by abandonment and succession in central Europe. To support their conservation and management, we investigated habitat use by the owly sulphur(Libelloides coccajus) , a flagship insect species for dry grasslands and scree slopes. We surveyed 32 previously inhabited sites and 24 previously unoccupied but suitable sites for adults and eggs, and characterized their association with macro- and microstructural habitat parameters.
20 sites occupied in 2020 indicate a decline in population sizes and range. Inhabited sites were juniper heaths and dry grasslands rich in open soil, drystone walls, erosion edges, or old stone quarries, but also scree slopes within coppiced woodland. Eggs were mostly found above open soil or rock surfaces, where they were laid south-facing on dry plant stalks that protruded horizontally above directly exposed, barren surfaces.
Our study highlights how land use abandonment and the resulting vegetation succession deteriorates habitat quality by increasing shading and isolation of the remaining dry grassland habitat patches. As a conservation measure, we recommend intense coppicing of emerging shrubs and trees, in particular along steep (scree) slopes as well as actively creating open soil and bare ground habitats. Extensive grazing and mowing can maintain open habitats of the owly sulphur, but only if combined with rotational removal of shrubs.
- Veröffentlicht am

1 Einleitung
Magere Trockenstandorte wie Wacholderheiden, Halbtrockenrasen oder Fels- und Schutthalden zählen zu den artenreichsten und gleichzeitig am stärksten rückläufigen Biotopen in Mitteleuropa (LUBW 2019). Waren zunächst vor allem Erstaufforstungen für den Rückgang dieser Lebensräume verantwortlich, sind es heute primär Sukzessionsprozesse in der Folge von Nutzungsaufgabe oder Pflegedefiziten, die durch die steigenden Nährstoffeinträge durch die Luft (Schaap et al. 2018) und eine verlängerte Vegetationsperiode infolge des Klimawandels noch beschleunigt werden können.
Durch mangelnde oder gänzlich aufgegebene Nutzung verschwinden zuerst lückige roh- und offenbodenreiche Stellen infolge der Verfilzung der Vegetation. Für zahlreiche Insektenarten trockener Standorte ist eine starke Abhängigkeit von derartigen Stellen belegt (etwa Buchweitz 1993, Streitberger et al. 2014). Die mikroklimatischen Gunstbedingungen (Wärmeeinstrahlung) sind vor allem für Präimaginalstadien (Eier und Larven) entscheidend für eine erfolgreiche Entwicklung (Fartmann & Hermann 2006).
Dieser Anforderung wird bei Schutz- und Pflegemaßnahmen für auffällige, als Imago flugaktive Insektenarten (etwa Schmetterlinge, Wildbienen) häufig zu wenig Rechnung getragen, da primär auf die (meist deutlich weniger spezifischen) Ansprüche der Imagines fokussiert wird.
Der Libellen-Schmetterlingshaft (Libelloides coccajus ) ([Denis & Schiffermüller, 1775]) aus der Ordnung der Netzflügler (Neuroptera) ist mit seinen schwarz-gelb gemusterten Flügeln und seinem schwirrenden Flug eine auffällige Erscheinung an Trockenstandorten. Aufgrund seines markanten Aussehens und seiner engen Biotopbindung kann er eine ideale Ziel- oder Schirmart zur Umsetzung von Naturschutzzielen darstellen.
Trotzdem sind bisher nur wenige grundlegende Informationen zu Lebenszyklus, Verbreitung oder Habitatansprüchen bekannt. Offene Fragestellungen wurden daher in einer im Jahr 2020 absolvierten Masterarbeit (Stork 2021) bearbeitet.
1.1 Autökologie des Libellen-Schmetterlingshafts
Der Libellen-Schmetterlingshaft ist ein räuberisch lebendes Insekt, das sich als Larve und Imago von kleinen Wirbellosen ernährt (Abb. 1). Hierbei orientiert sich die flugfähige, äußerst wendige Imago mithilfe ihrer gut ausgebildeten Facettenaugen und erbeutet Insekten im Flug (Aspöck & Aspöck 2007). Je nach Witterung und klimatischer Lage können Imagines zwischen Ende April und Ende Juni beobachtet werden (Wolf 2004).
Zur Paarung fliegen die Männchen die Weibchen in der Luft von unten an und versuchen diese zu umklammern. War dies erfolgreich, lässt sich das Paar in die Vegetation fallen und es kommt zur Kopulation (Mader 2018, Wachmann & Saure 1997, Abb. 1). Die Eiablage erfolgt an direkt besonnte, meist trockene Pflanzenstängel, häufig mit Mehrfachbelegungen in mikroklimatisch besonders begünstigten Bereichen (Fetz 1999, 2002). Die Gelegegrößen schwanken stark, mit im Mittel 20–70 Eiern (Wolf 2004, Abb. 2). Nach 2–4 Wochen schlüpfen die charakteristischen Larven, die optisch den Larven der Ameisenjungfern („Ameisenlöwen“) ähneln (Abb. 3).
Nach dem Aushärten der Kutikula lassen sie sich in die Vegetation fallen und leben als räuberische Lauerjäger im Substrat (Abb. 4). Hierbei sind ihnen die im Unterkiefer sitzende Giftdrüse mit ihrem lähmenden Sekret sowie wirkungsvolle Tarnung durch kleine Partikel aus der Umgebung (Staub, Sand, Detritus) behilflich (Wachmann & Saure 1997). Die Larven durchlaufen drei Stadien, die sich in der Regel über eine Dauer von zwei Jahren erstrecken (Badano & Pantaleoni 2014). Somit existieren zwei voneinander unabhängige Populationen gerader und ungerade Jahre, wobei eine fakultative ein- oder dreijährige Entwicklung einzelner Individuen nicht auszuschließen ist.
Seine holometabole Entwicklung schließtL. coccajus nach zweimaliger Überwinterung der Larve im Frühjahr mit dem Puppenstadium ab, aus dem nach einer Puppenruhe von circa 30 Tagen die Imago schlüpft (Badano & Pantaleoni 2014).
1.2 Bearbeitete Fragestellungen
Wir vergleichen die historische mit der aktuellen Verbreitung der Art im Regierungsbezirk Tübingen im Jahr 2020. Zudem beschreiben wir detailliert die Habitatansprüche von Imagines und Larven anhand folgender Fragestellungen:
- Wodurch sind besiedelte Imaginal- und Larvalhabitate charakterisiert?
- Mit welchen Parametern ist die Besiedlungswahrscheinlichkeit einzelner Untersuchungsgebiete assoziiert?
- Welche Parameter erklären innerhalb besiedelter Gebiete die Wahl der Eiablage-Standorte?
- Welche Veränderungen gab es in Gebieten, in denen Vorkommen der Art zwischenzeitlich erloschen sind?
- Aus diesen Befunden leiten wir Gefährdungspotenziale sowie Vorschläge für effektive Schutz- und Pflegemaßnahmen ab.
1.3 Untersuchungsgebiet
Als Untersuchungsgebiet diente der Regierungsbezirk Tübingen im südlichen Baden-Württemberg (Abb. 5). Der Libellen-Schmetterlingshaft ist hier in den Naturräumen 3. Ordnung Neckar- und Tauber-Gäuplatten, Schwäbisches Keuper-Lias-Land mit Schönbuch und Glemswald, Mittleres Albvorland Südwestliches Albvorland, Hohe Schwabenalb, Mittlere Kuppenalb, Mittlere Flächenalb sowie Baaralb und Oberes Donautal verbreitet (Tröger 2005).
2 Material und Methoden
2.1 Literaturrecherche und Expertenbefragung
Vor Beginn der Freilanderfassungen wurden Funddaten des Libellen-Schmetterlingshafts im Untersuchungsraum recherchiert und dazu Arten- und Gebietskenner befragt, Publikationen und Internetquellen sowie interne Gutachten zu geplanten und bestehenden Naturschutzgebieten im Regierungsbezirk Tübingen ausgewertet. Für die Freilanderfassungen wurden neben den Gebieten mit historischen und aktuellen Nachweisen weitere im räumlichen Zusammenhang zu den Fundstellen befindliche Potenzialgebiete in die Untersuchungen einbezogen. Hierdurch sollten zusätzlich zu den bereits bekannten Vorkommen neue Nachweise der Art erbracht werden.
2.2 Gebietskartierung – Imagines
Alle Gebiete wurden zur Flugzeit des Libellen-Schmetterlingshafts vom 01.05. bis zum 02.06.2020 einmal aufgesucht. Imagines wurden kartiert, indem die Habitate in möglichst gleichmäßigem Tempo über eine Zeitdauer von 20 Minuten abgeschritten wurden. Die Erfassungen erfolgten in Anlehnung an die Methodik des Tagfaltermonitorings Deutschlands (Kühn et al. 2008).
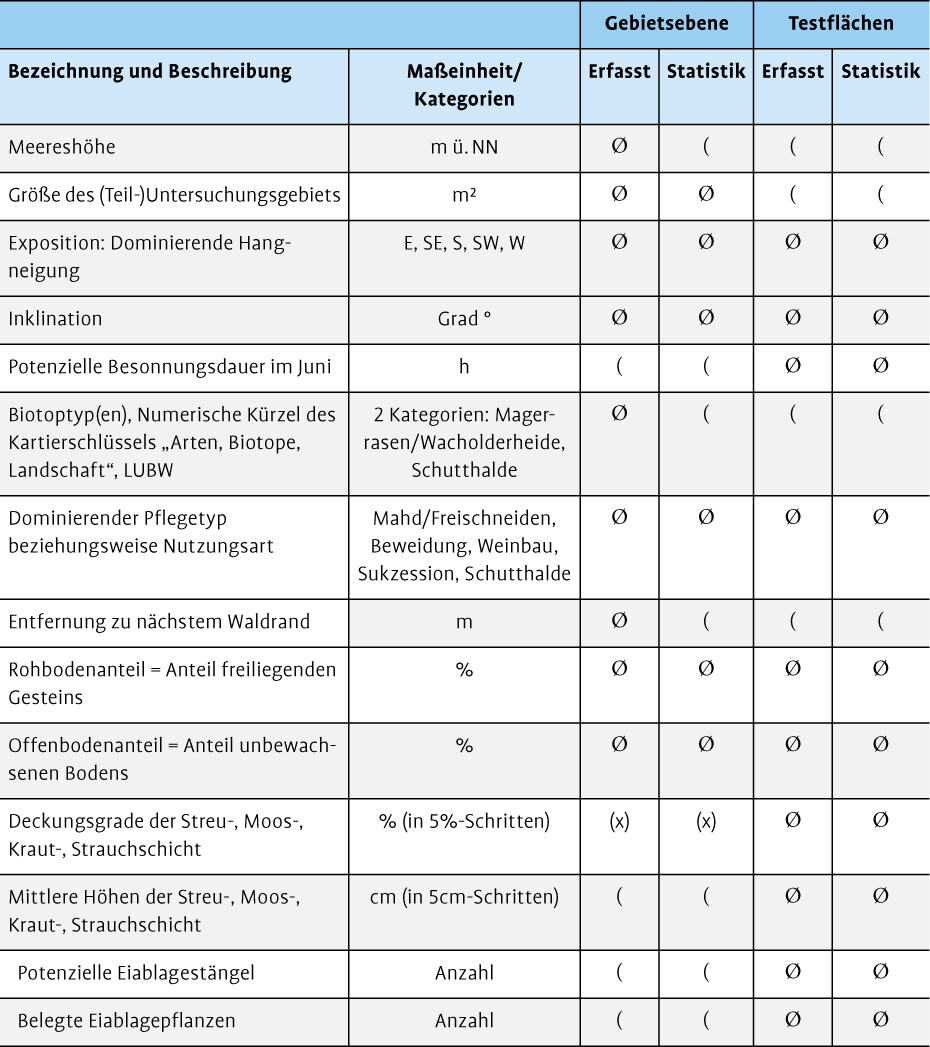
Alle gesichteten Hafte wurden mittels GPS (Garmin eTrex 10) erfasst. Da die Imagines nur bei Wärme und Sonnenschein aktiv sind, erfolgten die Kartierungen bei Temperaturen > 20 °C und wurden bei Bewölkung oder starkem Wind so lange unterbrochen, bis wieder geeignete Witterungsverhältnisse herrschten (Müller et al. 2012). Pro Gebiet wurden verschiedene Gebietsparameter erhoben, um die Ansprüche des Libellen-Schmetterlingshafts an seinen Lebensraum besser beschreiben zu können (Tab. 1).
2.3 Gebietskartierung – Eigelege
Gegen Ende der Flugzeit wurden Gebiete mit Nachweisen von Imagines zwischen dem 08.06. und dem 26.06.2020 zur Erfassung von Eigelegen erneut begangen. Eine wiederholte Kartierung der Gebiete ohne Imaginalnachweis konnte aus Zeitgründen nicht geleistet werden. Bei der Kartierung der Eigelege stand keine vollständige Erfassung des jeweiligen Kartiergebiets im Vordergrund, sondern eine qualitative Analyse bevorzugter Eiablagestellen. Hierfür wurden auf Literaturbasis (etwa Wolf 2004) Mikrohabitatparameter definiert, die in 1 × 1 m² großen Testflächen erhoben wurden (Tab. 1).
Die Auswahl der Testflächen erfolgte nicht zufällig, da mit dieser Methode keine ausreichende Anzahl an Testflächen mit Eigelegen erwartet werden konnte. Stattdessen wurden als geeignet erachtete Bereiche gezielt angesteuert, um dort nach Eigelegen zu suchen. Immer wieder wurden jedoch auch strukturell andersartige Flächen auf das Vorhandensein von Eigelegen überprüft. Wurde ein Eigelege gefunden, wurde um dieses herum ein Quadrat von 1 m Kantenlänge markiert und darin die Mikrohabitatparameter aufgenommen.
Pro Gebiet sollten auf diese Weise mindestens vier Probestellen untersucht werden. Gelangen weniger als vier Nachweise von Eigelegen an unterschiedlichen Stellen, wurden die Parameter in zufällig ausgewählten Nullflächen erhoben. Zusätzlich erfolgten im Hochsommer und Herbst sporadische Nachsuchen nach den Larven der Hafte, um neue Erkenntnisse über die bisher weitgehend unbekannten Larvalhabitate zu erlangen.
2.4 Statistische Auswertungen
Für die statistische Analyse berechneten wir generalisierte lineare Modelle (GLM) im Paket glmmTMB (Brooks et al. 2017) für die Software R (Version 4.04, R Core Team 2021). Zur Beschreibung der Verteilung adulter Schmetterlingshafte über die 56 Untersuchungsgebiete haben wir ein sogenannteshurdle -Modell erstellt, das zur Modellierung des hohen Anteils an Nullwerten zwei Prozesse unterscheidet:
Der erste modelliert die Absenz (0 Individuen) versus Präsenz ( 1 Individuum) von Imagines pro Fläche unter Verwendung einer Bernoulli-Verteilung, der zweite die Anzahl festgestellter Imagines auf den besiedelten Flächen unter Verwendung einer Poisson-Verteilung mit strikt positiven Werten. Als kategorischer Hauptprädiktor diente das Pflege- beziehungsweise Managementregime (Tab. 1).
Gebiete mit den Kategorien „Sukzession“ (n = 2) und „Weinbau“ (n = 4) waren mit zu geringen Stichprobenzahlen vertreten und zudem durchweg unbesiedelt, sodass sie in der Analyse nicht berücksichtigt wurden. Als kontinuierliche Kovariaten verwendeten wir die Flächengröße (Spanne: 0,4–33,8 ha, aufgrund stark rechts-schiefer Verteilung log10-transfomiert) sowie die Deckungsgrade von Rohboden (Spanne: 5–90 %, log10-transfomiert) und Gehölzen (Spanne: 5–55 %).
Die grafische Rohdatenexploration ergab keine Hinweise auf Interaktionen zwischen den Prädiktorvariablen, deutete jedoch einen nichtlinearen Zusammenhang zwischen Flächengröße und Imaginesanzahl an. Daher haben wir die Flächengröße zusätzlich als quadratischen Term in das Modell integriert. Die Prädiktoren wurden anhand der Varianz-Inflationsfaktoren (VIF < 3) auf hinreichende gegenseitige Unabhängigkeit, die Modellgüte unter Verwendung des Pakets DHARMa (Hartig 2021) auf Konformität der Residuenverteilung und Dispersion mit den Modellannahmen sowie auf Konsistenz zwischen den Rohdaten und anhand des Modells simulierten Daten überprüft. Für Post-hoc-Vergleiche zwischen den verschiedenen Gruppen in kategorischen Prädiktoren verwendeten wir das Paket emmeans (Lenth et al. 2019).
Das zweite Modell beschreibt die Anzahl Eigelege pro m²-Plot in Abhängigkeit mehrerer Charakteristika der kleinräumigen Mikrohabitate (Tab. 1). Das Vorgehen entsprach der oben aufgeführten Beschreibung, außer dass keinhurdle -Modell notwendig wurde, da die Verteilung der Zählvariable „Eigelege“ mit einer Poisson-Verteilung hinreichend angenähert wurde.
Numerische Prädiktoren umfassten die Inklination, die Dauer direkter Sonneneinstrahlung, die Deckungsgrade von Offenboden, Streu- und Krautschicht (jeweils log10-transformiert), die Höhe der Krautschicht sowie die Anzahl an Stängeln, die potenziell als Eiablagesubstrat dienen können (log10-transformiert). Zudem wurden die Anwesenheit von Sträuchern im Plot sowie die Exposition des Plots (Ost/Südost, Süd, Südwest) als kategorische Prädiktoren in das Modell übernommen.
Der Rohbodenanteil war stark mit mehreren anderen Prädiktoren korreliert und wurde daher nicht berücksichtigt. Für keinen der Prädiktoren deutete sich ein quadratischer Zusammenhang mit der Gelegeanzahl an. Da in der Regel mehrere 1m²-Plots innerhalb eines Untersuchungsgebiets lagen, wurde zur Vermeidung von Pseudoreplikation die Gebiets-ID alsrandom intercept -Faktor in das Modell integriert.
Karten wurden mit der freien Geoinformationssoftware QGIS, Version „Madeira“, erstellt. Für alle kartierten Gebiete wurde zudem ein Luftbildvergleich mittels historischer Orthofotos aus dem Jahre 1968 sowie aktueller Luftbilder angefertigt. Die historischen Orthofotos sind für ganz Baden-Württemberg über die Website www.leo-bw.de abrufbar. Sie sind sehr gut geeignet, strukturelle Veränderungen in Landschaften im Zeitraum der vergangenen fünf Jahrzehnte nachzuvollziehen.
3 Ergebnisse
3.1 Verbreitung im Regierungsbezirk
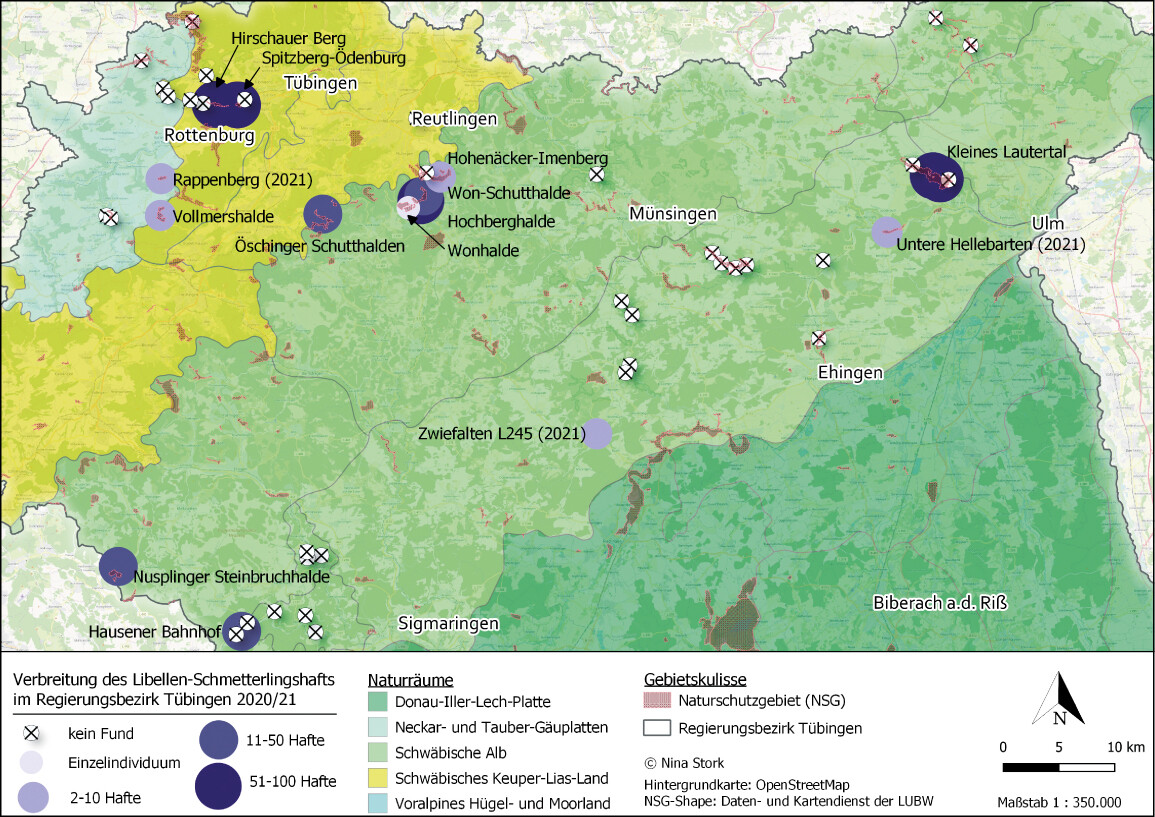
Die Datenrecherche ergab im Untersuchungsraum 20 Gebiete mit historischen oder aktuellen Nachweisen des Libellen-Schmetterlingshafts (Abb. 5). Daraus ergaben sich für die Kartierung 32 voneinander räumlich abgrenzbare Untersuchungsflächen. Hinzu kamen 24 Potenzialflächen, sodass insgesamt 56 Gebiete erfasst wurden. In 18 der 32 Untersuchungsflächen (56 %) und in zwei der 24 Potenzialflächen (8 %) konnte im Jahr 2020 ein Nachweis der Art erbracht werden.
Der Libellen-Schmetterlingshaft besitzt im Untersuchungsraum wenige Schwerpunktvorkommen, in denen er alljährlich und individuenreich nachgewiesen werden kann (2021 konnte ein weiteres, in der Verbreitungskarte dargestelltes Schwerpunktvorkommen im Raum Zwiefalten nach Hinweisen von Dr. Hendrik Turni erfasst werden). Diese Schwerpunktvorkommen befinden sich räumlich voneinander getrennt in verschiedenen Naturräumen (Abb. 5):
- Schönbuch & Glemswald (Spitzberg bei Tübingen: NSGs Hirschauer Berg und Spitzberg-Ödenburg),
- Lonsee-Flächenalb (NSG Kleines Lautertal),
- Albtrauf (NSGs Wonhalde-Spielberg und Imenberg-Hochäcker sowie Rutschungen bei Öschingen),
- Südwestalb (Steinbruchhalde bei Nusplingen),
- Oberes Donautal (Schutthalde bei Hausen im Tal).
Daneben existieren kleine Reliktvorkommen an den Südhängen des Rammerts und Nachweise von Einzeltieren, die möglicherweise auf verdriftete Individuen oder versuchte Neubesiedlungen zurückzuführen sind (etwa Großes Lautertal und NSG Salenberg auf der Schwäbischen Alb).
3.2 Habitatansprüche
Grundsätzlich werden zwei Habitattypen vom Libellen-Schmetterlingshaft besiedelt. Zum einen sind dies Trockenrasen im weiteren Sinne, inklusive ehemals weinbaulich genutzter Trockenhänge mit Lesesteinmauern im Albvorland (etwa Spitzberg bei Tübingen), die zwischenzeitlich mittels Mahd zu Naturschutzzwecken offengehalten werden (Abb. 6).
Diesem Lebensraumtyp sind ebenso flachgründige, lückige und rohbodenreiche Wacholderheiden am Albtrauf zuzuordnen. Wüchsigere Heiden werden nur besiedelt, wenn in Teilbereichen oder angrenzend Steinbrüche oder Abbruchkanten mit Rohbodenstellen existieren. Individuenarme Reliktvorkommen am Rammertsüdhang (NSG Vollmershalde und NSG Rappenberg) in Komplexen aus Halbtrockenrasen und Salbei-Glatthaferwiesen weisen darauf hin, dass in diesen Gebieten einst extremere Standortverhältnisse geherrscht haben müssen.
Zum anderen besiedeltL. coccajus sehr steile, vegetationsarme und von einer gewissen Dynamik geprägte Schutthalden mit hohem Rohbodenanteil. Diesem Typ sind Rutschungen und Schutthalden am nördlichen Albtrauf und im Oberen Donautal zuzuordnen. Eine intermediäre Stellung nimmt das NSG Kleines Lautertal im Naturraum Mittlere Flächenalb ein: Hier ist ein Komplex aus kleinflächigen Schuttbereichen und flachgründigen Trockenrasen ausgebildet – er bietet besonders günstige Voraussetzungen für den Libellen-Schmetterlingshaft (Abb. 7).
VonL. coccajus besiedelte Gebiete sind aufgrund der notwendigen Wärmegunst immer südwest- bis südostexponiert. Die Habitate sind meist stark geneigt (im Mittel 41°, Spanne 20–73 %) und weisen einen geringen Anteil an Gehölzdeckung auf. Imaginal- und Eiablagehabitate können sich deutlich voneinander unterscheiden. Während die mobilen Imagines häufig in Fluginsektenreichen Lebensräumen wie Streuobstwiesen oder extensiv genutzten Talwiesen jagen, sind die Eiablage- und Larvalhabitate deutlich enger abgegrenzt (siehe unten).
3.3 Assoziation aktueller Vorkommen mit Gebietsparametern
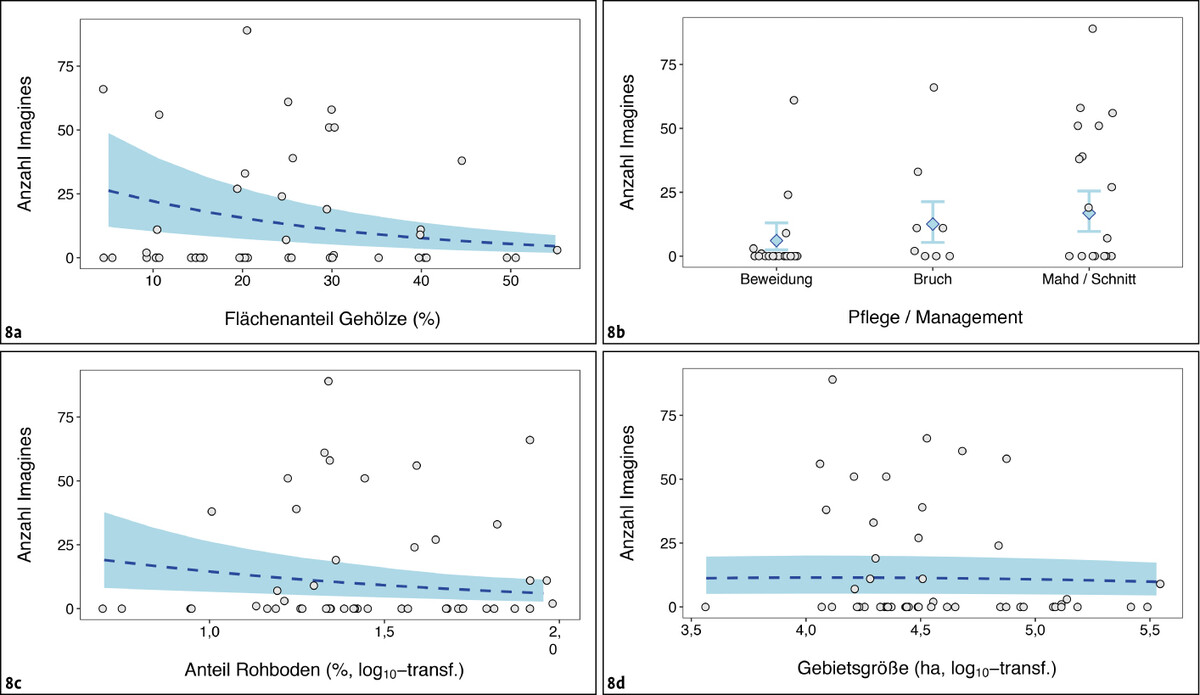
Die folgende statistische Analyse differenziert das Auftreten adulter Libellen-Schmetterlingshafte innerhalb von Gebieten, die aufgrund früherer Vorkommen oder ihrer Habitatausstattung grundsätzlich als geeignete Potenzialflächen eingestuft worden waren. Als stärkste Assoziation sank die ermittelte Anzahl Imagines deutlich mit zunehmenden Flächenanteilen an Gehölzen, im Mittel auf etwa 70,3 % des Ausgangswerts bei Zunahmen des Gehölzanteils um 10 % (95 % Konfidenzintervall 62,4–79,1 %; chi² = 41,21, df = 1, P < 0,001; Abb. 8 a).
Deutlich schwächer war das Auftreten mit dem Pflegeregime assoziiert. Sowohl gemähte und beweidete Flächen als auch Schotterhänge mit sporadischer Gehölzpflege wurden grundsätzlich besiedelt (Abb. 8 b). Die Antreffwahrscheinlichkeiten waren auf beweideten Flächen etwas geringer als auf Kalkschotterflächen (GLM z = 2,082, P = 0,037) sowie auf Flächen mit Mahd- oder Schnittregimen (z = 2,25, P = 0,024; Schnittregime entsprechen unregelmäßigem Rückschnitt von Stockausschlägen und aufkommenden Gehölzen).
In besiedelten Gebieten konnte aber kein signifikanter Unterschied in den mittleren Abundanzen ermittelt werden (GLM chi² = 4,89, df = 2, P = 0,087), sodass sich die in Summe vorhergesagten mittleren Abundanzen und ihrer Konfidenzintervalle stark überschneiden (Abb. 8 b). Zu beachten ist bei der Interpretation dieses Befundes, dass die Eiablagehabitate vielfach abseits der durch Mahd oder Beweidung erreichten Bereiche liegen dürften (siehe unten).
Ein relevanter Rohbodenanteil von meist > 10 % war eines der Auswahlkriterien für die bearbeiteten Untersuchungsgebiete – über diesen Anteil hinaus konnte keine Assoziation der Individuenzahlen mit diesem Parameter ermittelt werden (Abb. 8 c). Gleiches galt für die Gebietsgröße (Abb. 8 d).
3.4 Mikrohabitate
Insgesamt konnten 356 Eigelege vonL. coccajus gefunden werden, 153 davon in den 80 detailliert erfassten m²-Testflächen. In einigen Gebieten mit geringer Imaginaldichte konnte kein Gelegenachweis erbracht werden, während in anderen Gebieten mehr Eigelege als Imagines gezählt wurden.
Trotz häufig flächendeckender Suche konnten fast alle Eigelege (97 %) an vegetationsarmen Stellen gefunden werden (Abb. 9), nämlich 63 Gelege (41 %) über Offenboden, 86 Gelege (56 %) über Rohboden, aber lediglich vier Gelege (3 %) in Bereichen mit dichterer Vegetation. Die Eiablagestellen waren darüber hinaus meist durch eine leichte Nischen- oder Senkenlage gegen Wind geschützt und dadurch klimatisch zusätzlich begünstigt.
94 % der gefundenen Gelege waren an trockenen Stängeln überjähriger Vegetation abgelegt, durchschnittlich in 17 cm Höhe direkt über einem wärmeabstrahlenden Untergrund. In 38 % der Fälle konnte nach Erstfund eines Geleges mindestens ein weiteres Gelege innerhalb der dann näher untersuchten m²-Fläche gefunden werden und in 23 % der Fälle sogar am selben Pflanzenstängel, was die starke Selektion der Weibchen bezüglich der Auswahl der Eiablagestellen unterstreicht.
Wie schon auf Gebietsebene diente auch die statistische Analyse der Mikrohabitate dazu,innerhalb der oben als essenziell herausgearbeiteten, rohbodenreichen und stark besonnten Standorte die lokal genutzten Eiablagestandorte noch feiner zu differenzieren. Das Regressionsmodel erklärte etwa 30 % der beobachteten Variation (Pseudo-R²) in der Anzahl der Gelege pro m²-Plot.
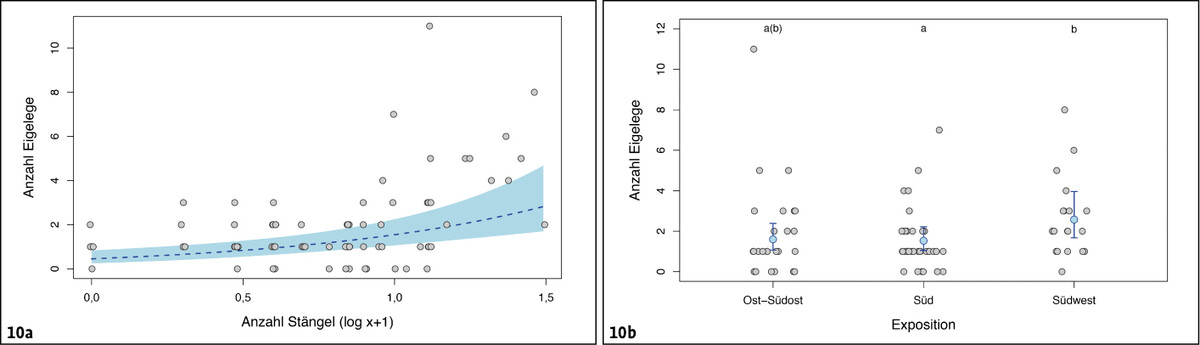
Von den zehn untersuchten Habitatparametern erreichten lediglich zwei statistische Signifikanz (für alle anderen: GLM chi² < 2,21 und P > 0,13). Die mittlere Anzahl an Eigelegen pro m² verdreifachte sich im Schnitt bei einer Zunahme der Anzahl vertrockneter Pflanzenstängel um den Faktor 10 (chi² = 12,99, df = 1, P = 0,0003, 95 % Konfidenzintervall 70–480 %, Abb. 10 a).
Nur als Tendenz deutete sich eine Assoziation der mittleren Anzahl Eigelege mit der Exposition an (chi² = 5,58, df = 2, P = 0,062). Demnach war die mittlere Anzahl Eigelege pro m²-Plot in Südwest-Exposition im Mittel um etwa 60 % höher als in Süd- beziehungsweise Ost-Südost-Exposition (Post-hoc t > 2,08, P < 0,10; 95 % Konfidenzintervall 8–105 %), während die beiden letzten nahezu identische Werte erreichten (Post-hoc t = 0,05, P = 0,99) (Abb. 10 b).
3.5 Larvensuche
Bei Nachsuchen am 27.07. und am 28.08.2020 konnten sieben Larven des Libellen-Schmetterlingshafts nachgewiesen werden. Die 5–8 mm großen Larven wurden im Bereich der Eiablagestellen mittels vorsichtigen Durchsuchens in der Substratschicht gefunden. Sie hatten sich mit verschiedenen Staub-, Gesteins- und Detrituspartikeln getarnt, verharrten bei Auffinden kurz, um dann die Flucht zu ergreifen.
Zwei Larven konnten auf Beute lauernd mit geöffneten Mundwerkzeugen am Eingang ihrer Verstecke aufgefunden werden. Bei zwei weiteren Nachsuchen im Frühjahr 2021 konnten in der Substratschicht auch circa 1,6 cm große Larven gefunden werden, die wahrscheinlich bereits 2019 geschlüpft waren und damit 2021 die Imago ergeben sollten.
4 Diskussion
4.1 Verbreitung
Der Libellen-Schmetterlingshaft konnte 2020 nur an etwa der Hälfte der zuvor bekannten Fundorte bestätigt werden, und alle sieben Gebiete mit letztem Nachweis vor 2010 blieben auch 2020 verwaist. In nur zwei der 24 untersuchten Potenzialflächen konnte die Art neu erfasst werden. Dies lässt auf einen Rückgang der Art im Laufe der vergangenen Jahre schließen. Vergleichbar ist die Situation im Kanton Aargau, wo in jüngerer Zeit nur noch vier von ursprünglich 22 bekannten Populationen bestätigt werden konnten (Müller et al. 2012).
Aufgrund der zweijährigen Entwicklung der Art besteht die Möglichkeit, dass Populationen gerader und ungerader Jahre unterschiedlich individuenreich sind und damit eine einjährige Erfassung nicht ausreichend ist. Dieses Phänomen ist auch von verschiedenen Tagfaltern wie dem Weißbindigen Mohrenfalter (Erebia ligea ) bekannt (Bamann 2016). Für eine verbesserte Datenlage müssten daher die Kartierungen zumindest in Gebieten ohne aktuellen Nachweis in einem ungeraden Jahr wiederholt werden. (Dies erfolgte zumindest regional 2021. Hierbei konnte die Art im hier als unbesiedelt eingestuften Gebiet NSG Rappenberg am Rammert-Südhang mit zwei Individuen bestätigt werden.)
Weiterhin lagen im Datenarchiv von einigen Gebieten nur Nachweise von Einzelindividuen vor. Hierbei könnte es sich um verdriftete Individuen aus benachbarten Populationen gehandelt haben, die am Fundort keine bodenständigen Vorkommen aufbauen konnten. Alternativ sind individuenarme Restbestände ehemals größerer Populationen denkbar. Generell war das Jahr 2020 ein schlechtes „Insektenjahr“ (Kühn et al. 2021), weshalb Wieder- und Neunachweise bei wiederholter Kartierung nicht auszuschließen sind.
Ein intaktes Metapopulationsnetz, wie es für viele mobile Insektenarten notwendig ist (Hanski 1998), lässt sich beim Libellen-Schmetterlingshaft angesichts der großen Distanzen zwischen den wenigen rezenten Vorkommen nicht (mehr) erkennen. Lediglich die wenigen räumlich noch stärker geclusterten Reliktvorkommen rund um den Spitzberg bei Tübingen oder im Kleinen Lautertal dürften Metapopulationen im kleinen Maßstab ausbilden, deren Teilpopulationen vermutlich noch im Individuenaustausch stehen – Nachweise über Markierungsstudien stehen aber noch aus.
4.2 Habitatansprüche
Die vonL. coccajus im Untersuchungsgebiet besiedelten Habitate stimmen weitgehend mit Literaturangaben überein (Fetz 1999, Müller et al. 2012, Wolf 2004). Auffällig sind wiederholte Beobachtungen jagender Imagines in Habitaten, die nicht den klassischen Trockenlebensräumen entsprechen (zum Beispiel Streuobstwiesen, mesophiles Grünland, Abb. 6, 7 und 11). Hierdurch kann der Eindruck entstehen, dass die Art in vielen unterschiedlichen Lebensräumen vorkäme und vergleichsweise anspruchslos sei, wodurch die gegenwärtige Gefährdungssituation gegebenenfalls falsch eingeschätzt wird.
Entscheidende Habitatbestandteile für eine erfolgreiche Reproduktion der Art sind nach den vorliegenden Befunden roh- und offenbodenreiche, schütter bewachsene und stark besonnte Steilhänge. Derartige Strukturen sind im südeuropäischen Hauptverbreitungsraum der Art erheblich häufiger, weshalb sie dort als euryök eingestuft wird (Aspöck et al. 1980).
Der offene Charakter besiedelter Habitate wird zudem durch die signifikante Bevorzugung gehölzarmer Lebensräume unterstrichen. Die weitgehende Indifferenz der Besiedlungswahrscheinlichkeiten gegenüber den Pflegeregimes in der vorliegenden Analyse halten wir für nur bedingt aussagekräftig. Vielmehr wird es von den lokalen strukturellen Standortbedingungen abhängen, ob selbst bei einem Mahdregime, das per se eine geschlossene Grasnarbe fördert, dennoch in räumlicher Verzahnung Rohboden- oder Schotterflächen vorhanden sind, die eine erfolgreiche Reproduktion ermöglichen (siehe Mikrohabitate).
In individuenreichen Kernvorkommen (Spitzberg und Kleines Lautertal) scheint dies aktuell noch gegeben, in anderen Gebieten wird es aber gezielter Bodenstörungen durch Beweidung oder maschinelle Eingriffe bedürfen, um derartige Strukturen zu erhalten oder wiederherzustellen.
4.3 Mikrohabitate
Obwohl wissenschaftliche Studien für verschiedene Insektentaxa deutlich die Bedeutung der Präimaginalhabitate gezeigt haben (zum Beispiel Fartmann & Hermann 2006), orientiert sich die Umsetzung von Schutzmaßnahmen häufig zu sehr an den Ansprüchen der Imagines (zum Beispiel Nektarquellen) und zu wenig an den mikroklimatischen und -strukturellen Ansprüchen der wenig mobilen Präimaginalstadien. Dabei ist die Bedeutung von offenen, lückigen Strukturen und warmen Mikroklimaten für die Eiablage und eine erfolgreiche Larvalentwicklung für zahlreiche Insektenarten hinreichend belegt (zum Beispiel Anthes et al. 2003, Fartmann 2004, Kettermann et al. 2020).
Der thermophile Libellen-Schmetterlingshaft ist in besonderem Maße von solchen Stellen abhängig: Sie müssen vollsonnig an stark geneigten Südhängen gelegen sein, einen hohen Roh- oder Offenbodenanteil und überjährige Stängel für die Eiablage aufweisen. Kleinflächige Schuttbereiche können diese Aspekte ebenso erfüllen wie Trockenmauern oder Offenbodenstellen. Auffällig ist, dass die Art auf der klimatisch ungünstigen Schwäbischen Alb an sich stark erwärmende Schuttbereiche gebunden scheint, während im wärmeren Albvorland (zum Beispiel Spitzberg) auch Offenbodenstellen in Halbtrockenrasen für eine erfolgreiche Reproduktion ausreichen können.
Sind die genannten Grundvoraussetzungen erfüllt, können besonders geeignete Mikrohabitate durch eine hohe Anzahl an überjährigen Pflanzenstängeln, bevorzugt in Südwestexpositionen, charakterisiert werden. Eine hohe Anzahl überjähriger Stängel bietet viele Möglichkeiten zur Eiablage mit einem sich kleinflächig ändernden Mikroklima. Südwestexpositionen zeichnen sich durch länger anhaltende Besonnung am Nachmittag aus, wodurch sich diese Standorte stark aufheizen und damit in besonderem Maße den xerothermen Ansprüchen des Libellen-Schmetterlingshafts entsprechen.
4.5 Gefährdung
Während die rezente Klimaerwärmung den thermophilen Libellen-Schmetterlingshaft mittelfristig begünstigen könnte, wirken sich Sukzessionsprozesse durch Nutzungsaufgabe und Pflegedefizite auf Grenzertragsstandorten negativ auf die Bestände der Art aus. Es handelt sich damit um gegenläufige Prozesse, wobei bisher die negativen Auswirkungen etwaige positive Effekte durch den Klimawandel bei Weitem überlagern.
Beim Vergleich historischer und aktueller Luftbilder der (ehemals) besiedelten Habitate ist eine Veränderung besonders augenfällig: Die Gehölzdeckung hat in den meisten Lebensräumen stark zugenommen (Abb. 12). Hierdurch verkleinert sich nicht nur die besiedelbare Habitatfläche, sondern die Gehölze tragen durch Beschattung und Zerschneidung zu einer Verminderung der Habitatqualität und einer Isolation der verbliebenen Habitatpatches bei.
In zwei Veröffentlichungen wird die für den Libellen-Schmetterlingshaft positive Wirkung von Gehölzreihen als Windschutz hervorgehoben (Müller et al. 2012, 2013). Dies mag für noch sehr großflächig offene Lebensräume zutreffen, in den Untersuchungsgebieten wird die zunehmende Gehölzdeckung jedoch als eindeutig wichtigster Gefährdungsfaktor angesehen. Zudem liegt der Fokus im Hinblick auf den Windschutz oftmals zu stark auf den Imagines, während die Ansprüche der Präimaginalstadien vernachlässigt werden (siehe oben).
Auch bei Tagfaltern wird immer wieder auf den notwendigen Windschutz für den Goldenen Scheckenfalter (Euphydryas aurinia ) oder den Blauschillernden Feuerfalter (Lycaena helle ) hingewiesen (Fischer et al. 1999, Ulrich 2007). Beide Arten können jedoch auch in vollständig gehölzfreien Habitaten vorkommen (Bamann & Dittrich 2017, Steiner et al. 2006), wobei an dieser Stelle nicht die grundsätzlich positive und strukturgebende Wirkung niedriger Gehölze etwa in Form von Niederhecken und junger Sukzessionen infrage gestellt werden soll.
Eine zunehmende Gehölzdeckung geht meist mit mangelhafter Beweidung oder Aufgabe der Mahd einher. Dies führt zu Verfilzung der Vegetation und zu Sukzession, welche ebenfalls in vielen Untersuchungsgebieten festgestellt wurde und einen weiteren Gefährdungsfaktor fürL. coccajus darstellt. Insbesondere steile und offenbodenreiche Stellen wie Schuttbereiche, Abbruchkanten oder Rutschungen werden häufig aufgrund ihrer Unzugänglichkeit bei der Pflege ausgespart, wodurch Schlüsselelemente für den Libellen-Schmetterlingshaft und andere thermophile Insektenarten verloren gehen.
Aufgrund der Eiablage an trockene, überständige Stängel und der etwa 30 Tage dauernden Eientwicklung von etwa Mitte Mai bis Mitte Juli (je nach Eiablagezeitpunkt) kann eine Nutzung des Habitats in diesem Zeitraum, etwa durch Abmähen oder Abweiden, ebenfalls negative Folgen haben. Verschiedene Autoren empfehlen daher eine jahreszeitlich späte Nutzung ab dem 1. August, wenn die Larven sicher geschlüpft sind (Fetz 1999, Wolf 2004).
Dem entgegen steht jedoch eine mögliche schleichende Verschlechterung der Habitatqualität durch die dauerhaft späte Nutzung, welche daher im Einzelfall abzuwägen ist. Aufgrund der spezifischen Lage der Eiablagestellen über Schutt oder Offenboden wird im Rahmen dieser Studie nicht von einer starken Gefährdung durch jahreszeitlich frühere Mahd oder Beweidung ausgegangen, solange diese nicht flächig erfolgt.
Auch in Schutthalden, Rutschungen und lichten Steppenheidewäldern stellt die Aufgabe oder Vernachlässigung der Pflege den Hauptgefährdungsfaktor durch forcierte Sukzession dar. Es ist mittlerweile belegt, dass diese extrem steilen und unzugänglichen Waldbereiche entlang des Albtraufs in früheren Zeiten regelmäßig von der lokalen Bevölkerung zur Brennholzentnahme genutzt wurden (Gatter & Mattes 2018, Schmidt 1995). Es handelt sich also keinesfalls, wie früher häufig angenommen (zum Beispiel Gradmann 1936), um vom Menschen unberührte Reliktstandorte.
Luftbildvergleiche zeigen, dass es auch an diesen extremen Standorten im Laufe der vergangenen Jahrzehnte zu einschneidenden Sukzessionsprozessen kam, die zahlreiche für den Libellen-Schmetterlingshaft potenziell geeignete Lebensräume zuwachsen ließen. Hiervon sind auch weitere anspruchsvolle Lichtwaldarten des Albtraufs wie Bergkronwicken-Widderchen (Zygaena fausta ), Elegans-Widderchen (Zygaena angelicae elegans ) oder Berglaubsänger (Phylloscopus bonelli ) negativ betroffen (Gatter 1997, Hofmann 1994, Kiefer 2015, Maurer 2016).
4.6 Pflegeempfehlungen
Die umfangreiche Rücknahme von Gehölzen kann als wichtigste Schutzmaßnahme für den Libellen-Schmetterlingshaft gelten. Hierbei sollten insbesondere steile und unzugängliche Offen- und Rohbodenbereiche, Abbruchkanten, ehemalige Steinbrüche und vergleichbare Strukturen freigestellt werden. Eine Freistellung sollte auch dann vorgenommen werden, wenn eine Folgepflege durch Beweidung nicht sichergestellt ist und gegebenenfalls in mehrjährigen Abständen eine motormanuelle Nachpflege notwendig werden kann.
Allein durch die Gehölzentnahme wird in der Regel ein höherwertiger Lebensraum geschaffen, der durch sofortige Bereitstellung junger Sukzessionsstadien für viele Arten Habitatfläche schafft (zum Beispiel für das Platterbsen-WidderchenZygaena osterodensis , Prosi & Dolek 2020, Wagner 2006).
Die Schaffung einer lückigen und offenbodenreichen Vegetation ist vor allem im klimatisch begünstigten Albvorland von Bedeutung, da hier keine größeren Schuttbereiche existieren. Dies kann – gegebenenfalls nach Abwägung von Zielkonflikten mit anderen lokalen Schutzzielen – durch eine Intensivierung der Mahd oder Beweidung erreicht werden, wobei darauf geachtet werden sollte, dass während der Zeit der Eiablage und Entwicklung der Eigelege nicht zu intensiv und flächig vorgegangen wird.
Frühe Mahd oder Beweidung auf Teilflächen kann wüchsigen Standorten effektiv Nährstoffe entziehen und lückige Vegetationsstrukturen schaffen (zum Beispiel Hely et al. 2018, Kapfer 2010). Kleinflächige Rohbodenstellen können zudem durch die vollständige Entnahme von Gehölzen mitsamt Wurzelwerk oder durch die Entnahme von Koniferen, wie Waldkiefer, Wacholder und Fichte, geschaffen werden. Eine weitere gut geeignete Methode zur Schaffung von Roh- und Offenbodenstellen sowie von offenen Böschungen und Abbruchkanten ist der Abtrag von Oberboden. Solche Flächen dienen zahlreichen weiteren, teilweise hochgradig gefährdeten Insektenarten als Habitat (zum Beispiel dem Deutschen SandlaufkäferCylindera germanica , Trautner 2018).
Auch in Lichtwaldhabitaten (Schutthalden, Steppenheidewälder) ist ein langfristiges Gehölzmanagement zu etablieren, das eine Offenhaltung der Lebensräume gewährleistet (vergleiche Jotz et al. 2017). Diese kann am ehesten durch Gehölzentnahme erfolgen. Aufgrund der extremen Standortverhältnisse sind die Eingriffe nur in größeren Abständen (zum Beispiel alle 5–10 Jahre je nach Wüchsigkeit des Standorts) notwendig, wobei aufkommender Jungwuchs gegebenenfalls in kürzeren Zeitabständen zurückgedrängt werden muss.
Keinesfalls dürfen derartige Standorte als Prozessschutzflächen (zum Beispiel Waldrefugien, Bannwälder, Kernzonen von Biosphärengebieten) ausgewiesen werden. Hierdurch wird eine zukünftig notwendige Pflege entweder deutlich erschwert oder sogar unmöglich gemacht. Dies ist leider in der Vergangenheit in Unkenntnis der Pflegeabhängigkeit und sehr hohen artenschutzfachlichen Bedeutung dieser Standorte bereits in größerem Umfang geschehen.
Fazit für die Praxis
- Der Libellen-Schmetterlingshaft ist eine hinsichtlich seiner mikrostrukturellen Anforderungen höchst anspruchsvolle und damit gut geeignete Leitart offener Trockenrasen und Schutthalden.
- Der Schlüssel zum Verständnis der Ansprüche dieser Art liegt bei den Präimaginalstadien.
- Wird die Nutzung in besiedelten Habitaten auf sie abgestimmt, profitieren zahlreiche weitere gefährdete Arten von diesen Maßnahmen.
- Vor allem der Gehölzreduktion an besonders steilen und schwer zugänglichen Standorten sollte zukünftig in der Naturschutzpflege mehr Aufmerksamkeit geschenkt werden.
Literatur
Aus Umfangsgründen steht das ausführliche Literaturverzeichnis unter Webcode NuL2231 zur Verfügung.
Dank
Wir bedanken uns herzlich bei Herrn Prof. Andreas Martens (PH Karlsruhe) für die Erstbetreuung der Masterarbeit sowie bei Gabriel Hermann, Florian Straub und Jürgen Trautner (Arbeitsgruppe für Tierökologie und Planung Filderstadt) für die Durchsicht des Manuskripts und zahlreiche hilfreiche Anmerkungen.
Kontakt

Thomas.Bamann@rpt.bwl.de

nils.anthes@uni-tuebingen.de

Die Arbeit wurde eingereicht am 10. 12. 2021, angenommen am 10. 07. 2022
-
Abb. 1: Imagines des Libellen-Schmetterlingshafts bei der KopulaThomas Bamann -
Abb. 2: EigelegeThomas Bamann -
Abb. 3: Schlüpfende Larven am EigelegeThomas Bamann -
Abb. 4: Larve im SubstratThomas Bamann -
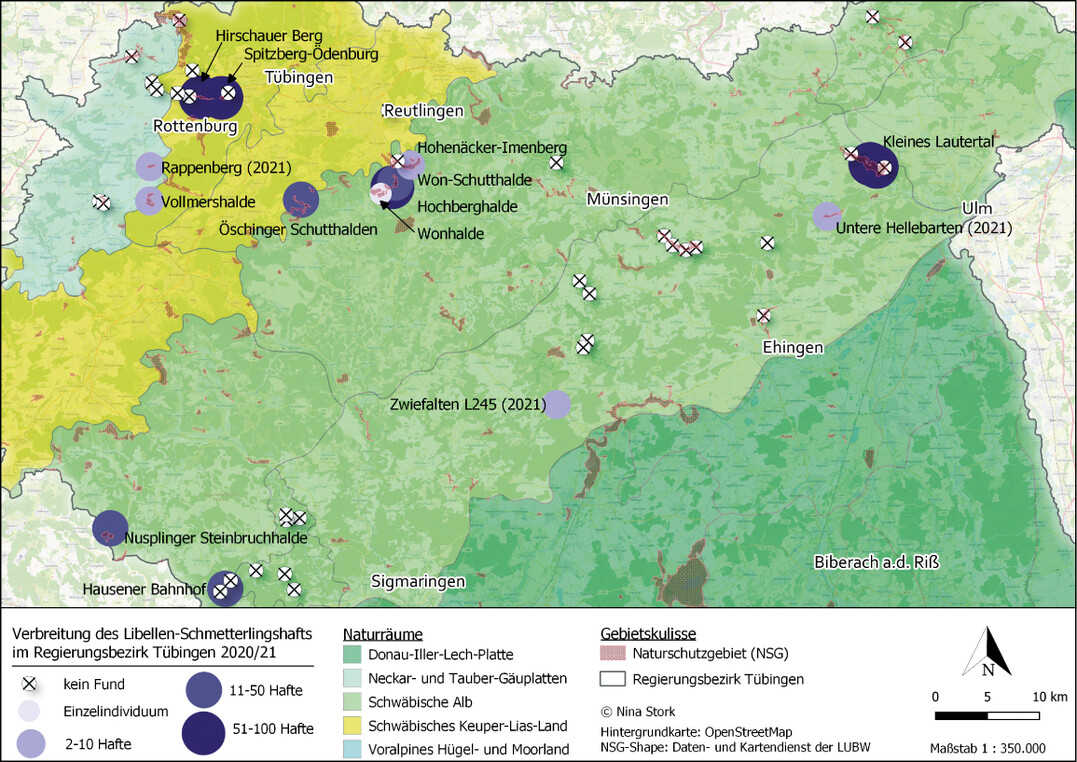
Abb. 5: Karte mit aktuellen und historischen Fundstellen des Libellen-Schmetterlingshafts im UntersuchungsgebietNina Stork -

Abb. 6: Imaginal- und Larvalhabitat im NSG Hirschauer Berg. Larvalhabtiate befinden sich oft in Offenbodenreichen und stark besonnten Mikrohabitaten (blau markierte 1m² Probefläche im linken Foto). Imagines fliegen auch in stärker versaumten Bereichen, wie der im rechten Foto blau markierten unteren Hanglage, während sich die Larvalhabitate in den exponiertesten Steillagen befinden (gelb markierter Bereich).Nina Stork -

Abb. 7: Imaginal- und Larvalhabitat im NSG Kleines Lautertal. Im Kleinen Lautertal finden sich die Eiablagestellen in Schuttbereichen, die von lückiger Vegetation durchsetzt sind (linkes Foto). Imagines nutzen die insektenreichen Talwiesen zur Jagd (blauer Bereich im rechten Foto), während die Larvalhabitate in den rohbodenreichen Steilhängen (gelber Bereich) zu finden sind.Nina Stork -
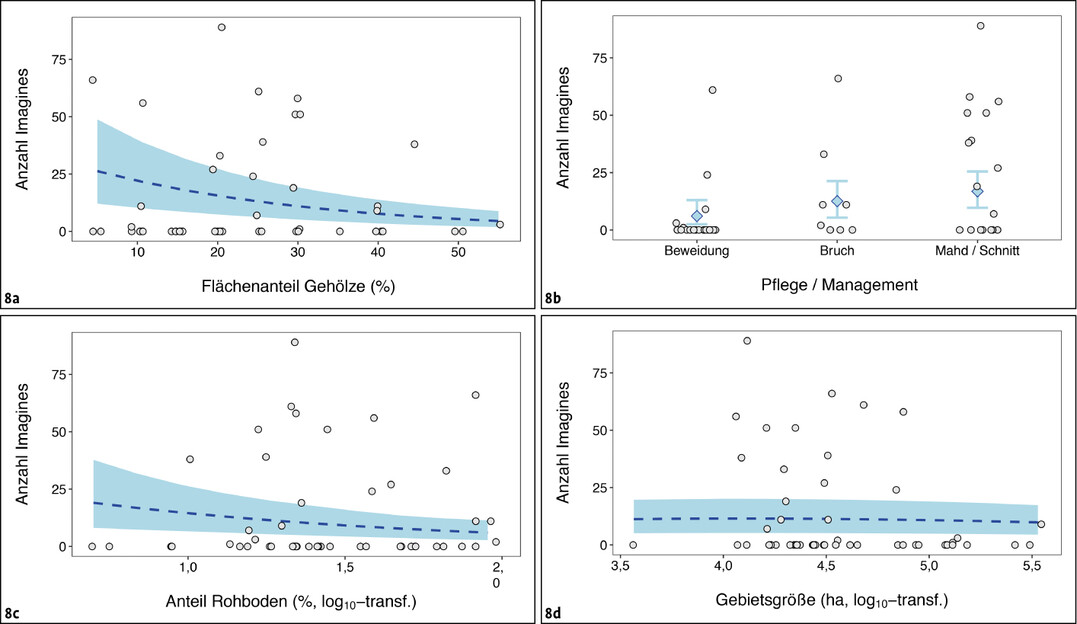
Abb. 8: Abundanz adulter Schmetterlingshafte in Abhängigkeit von verschiedenen Gebietscharakteristika. Rohdaten (graue Punkte) zeigen die Anzahl festgestellter Imagines, wobei Werte bei y = 0 unbesiedelte Gebiete darstellen. Blau dargestellt sind die aus den statistischen Modellen extrahierten Mittelwerte (gestrichelte Linie beziehungsweise Rauten) und deren 95 %-Vertrauensbereich (Schattierung beziehungsweise Fahnen).Nils Anthes -
Abb. 9: Mikrohabitat mit Eigelege im NSG Kleines LautertalNina Stork -
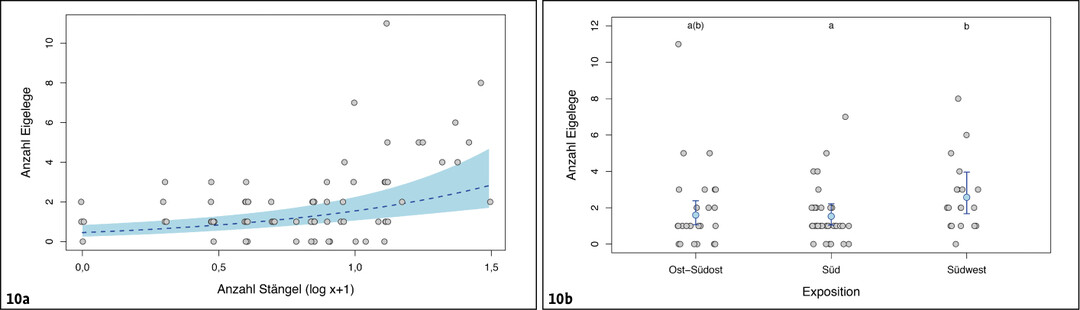
Abb. 10: Anzahl Eigelege des Schmetterlingshafts in Abhängigkeit von der Anzahl verfügbarer trockener Pflanzenstängel sowie der Exposition der untersuchten Fläche. Zur Darstellung siehe Abb. 8.Nils Anthes -
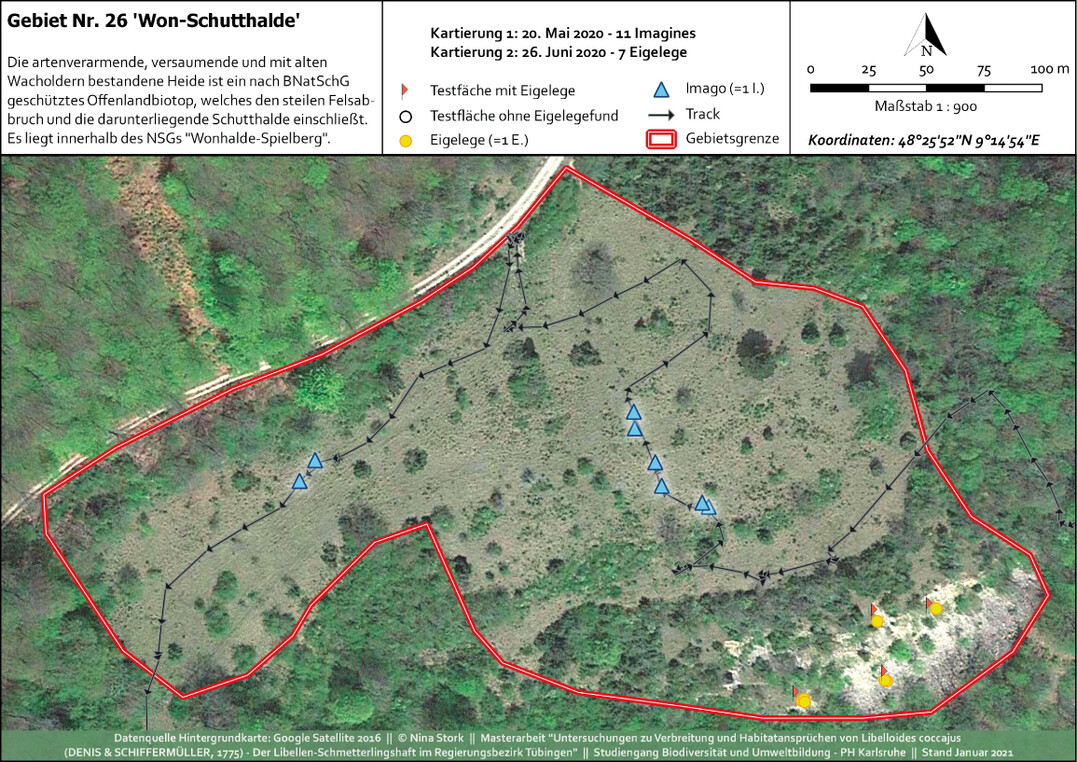
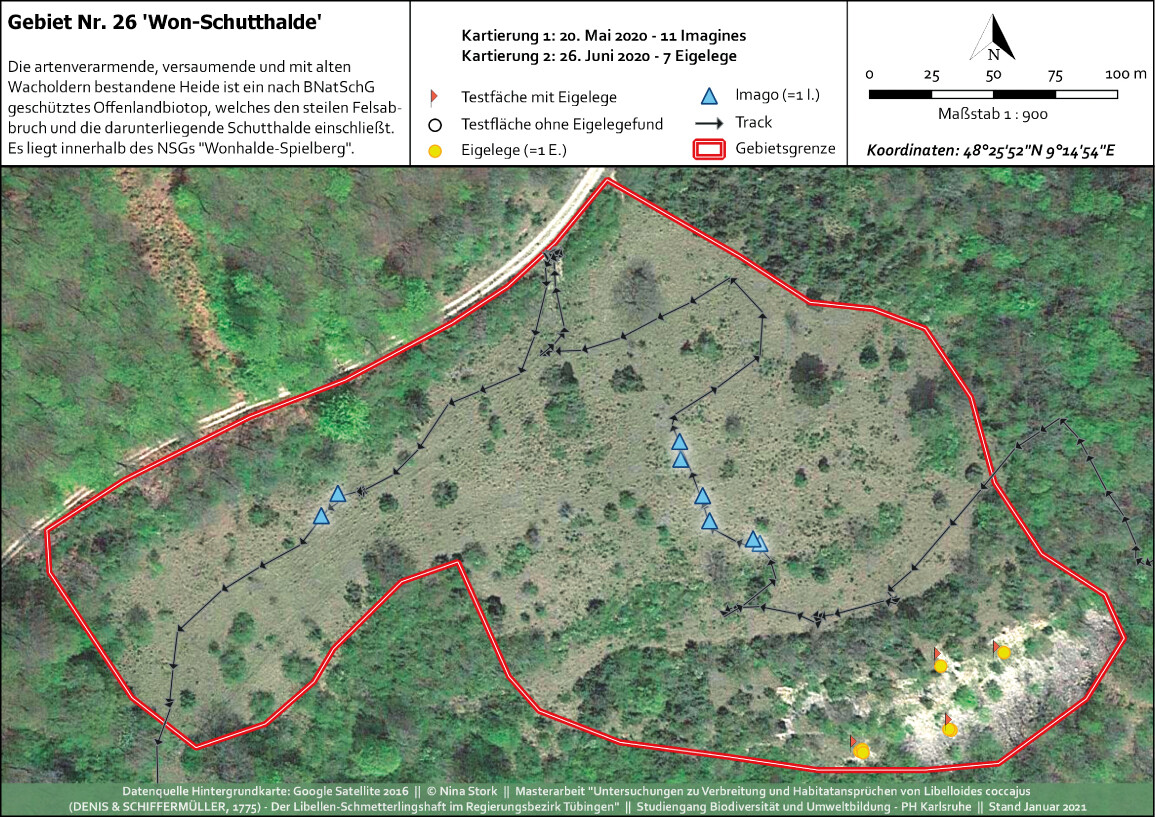
Abb. 11: Kartierkarte mit Imaginal- und Präimaginal-Nachweisen im NSG Wonhalde-SpielbergNina Stork -
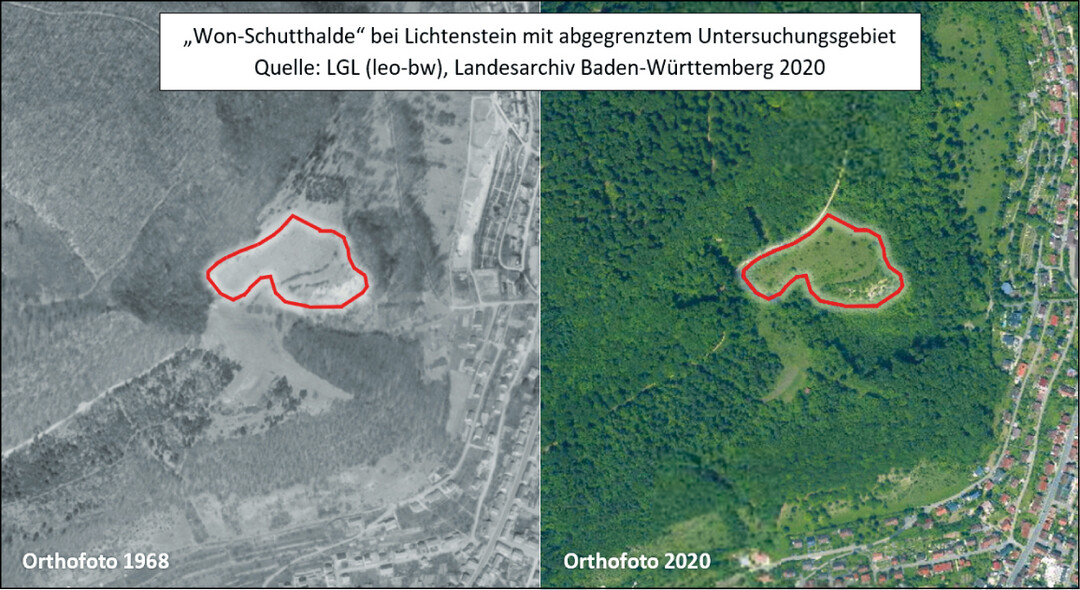
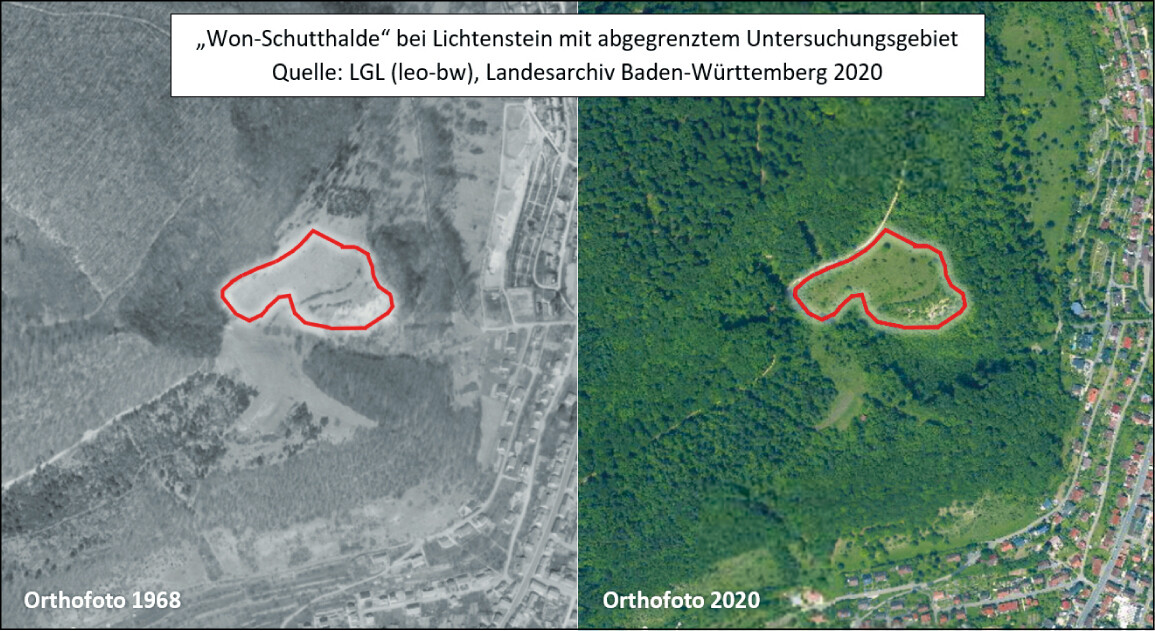
Abb. 12: Luftbildvergleich NSG Wonhalde-Spielberg 1968 und 2020.Nina Stork -
-
Bamann -
Anthes -
Stork
















Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.