Bestandsentwicklung invasiver Neophyten am Unterlauf der Ilz (Bayerischer Wald)
Abstracts
Die Ilz ist neben dem Regen das bedeutendste Fließgewässer des Bayerischen Waldes. Sie entspringt in den Kammlagen des Mittelgebirges und mündet bei Passau in die Donau. Als Migrationskorridor verschneiden sich an der Ilz talwärts wandernde Elemente aus den Hochlagen des Bayerischen Waldes mit thermophilen Arten, die aus dem milden Donautal bergwärts migrieren. Das Ilztal gilt daher als ausgesprochen artenreicher Landschaftsausschnitt innerhalb des silikatischen (= artenarmen) Bayerischen Waldes. Diese Vielfalt ist bedroht durch die sukzessive Ausbreitung invasiver Neophyten. Viele invasive Neophyten sind in der Lage, sich aus verschleppten Rhizom- oder Sprossteilen zu regenerieren, weshalb gut gemeinte Bekämpfungsmaßnahmen vielerorts nicht immer den gewünschten, oft sogar einen gegenteiligen Effekt haben. Zudem können Hochwasserereignisse mit verstärkter Ufererosion die Rhizom-Verbreitung fördern. In hochdynamischen Flusstälern wie dem der Ilz sind daher Beobachtungen zur Bestandsentwicklung von Neophyten von großer Bedeutung, um Ausbreitungstendenzen erkennen und gegebenenfalls gezielte Gegenmaßnahmen einleiten zu können. 2009 wurde am Unterlauf der Ilz die Verbreitung invasiver Neophyten hochaufgelöst kartiert. Im Sommer 2015 wurde eine Wiederholungskartierung durchgeführt, um die Bestandsentwicklung am Unterlauf der Ilz im Zeitraum zwischen den beiden Kartierungszeitpunkten zu beurteilen, insbesondere auch aus dem Blickwinkel zwischenzeitlich durchgeführter Bekämpfungsmaßnahmen und des außergewöhnlichen Hochwassers im Juni 2013 heraus.
Development of invasive neophytes along the lower course of the Ilz River in the Bavarian Forest Changes between 2009 and 2015
Next to the Regen River the Ilz River is the most important watercourse of the Bavarian Forest. It rises in the uplands, and close to the city of Passau it enters the Danube River. The Ilz River functions as an important migration corridor for organisms, on the one hand for upland species moving downstream and on the other hand for thermophilous species from the Danube valley migrating upstream. Due to this function the Ilz valley is considered a rather species rich landscape section within the siliceous (i.e. species poor) Bavarian Forest. This high species diversity is at risk by the gradual spreading of invasive neophytes. Many invasive neophytes are able to disperse vegetatively by displaced fragments of rhizomes and shoots, which is why well-intentioned control measures in many places often even cause counterproductive effects. In addition, flood events can lead to major riparian erosion including the dispersal of rhizomes. In highly dynamic river valleys such as the Ilz valley observations on population development of invasive neophytes are important to identify tendencies of expansion and to implement control measures if required. In 2009 invasive neophytes were mapped with high resolution at the lower course of the Ilz River. The mapping was repeated in summer 2015 to evaluate the population development between the two mapping dates. An additional focus was laid on the effects of the control measures during this period of time and the massive flood event of June 2013.
- Veröffentlicht am

1 Einleitung
Floristisch betrachtet (und wohl nicht nur) ist Deutschland wieKowarik(2003) es treffend formuliert hat seit jeher ein traditionelles Einwanderungsland. Den Großteil der heute etablierten Pflanzen verdankt Deutschland der postglazialen Rückwanderung von im Pleistozän auf südlich gelegene Refugialräume zurückgedrängten Arten. Hinzu kommen anthropogene Veränderungen in der Artenzusammensetzung, die sich für Deutschland mindestens bis zur neolithischen Revolution zurück nachweisen lassen (Brandes2008,Kowarik2003,Petrischak2014). Bei nicht-einheimischen Arten, die nach der Entdeckung der Neuen Welt durch Christopher Kolumbus im Jahr 1492 bewusst oder unbewusst durch den Menschen verschleppt wurden, dabei geographische Barrieren (Böhmer2011) überwanden und sich schließlich in einem neuen Raum etablieren konnten, spricht man von Neophyten (im Gegensatz zu Archäophyten, deren Verschleppung vor diesem Zeitpunkt stattfand). Viele dieser Neubürger wurden sicher über kontaminierte Importgüter (Heu und Stroh als Polster fragiler Waren, Wolle, agrarische Produkte etc.) eingebracht, der weitaus größere Teil zählt aber zu den Ergasiophygophyten (aus Kultur verwilderten Pflanzen), und hier insbesondere zu Gartenflüchtlingen (Loos&Schmitt2011,Nentwig2010,Petrischak2014).
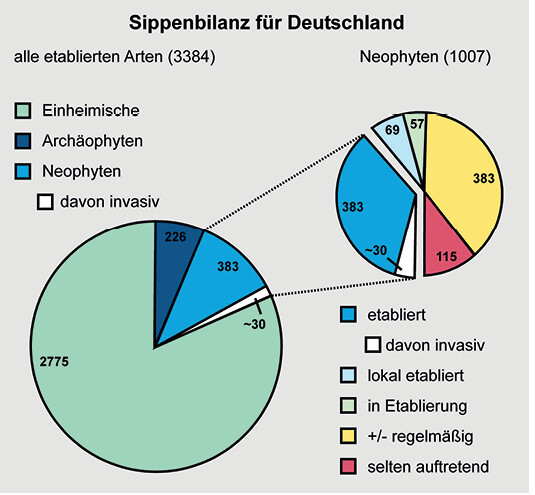
Die Sippenbilanz der Flora Deutschlands (Abb. 1) weist 383 Pflanzen (11,3 %) als etablierte Neophyten aus, von denen 30 als invasiv gelten. In Niederbayern liegt der Anteil fest eingebürgerter Neophyten an der heimischen Flora mit 17 % sogar noch etwas höher als in Gesamt-Deutschland(Zahlheimer2001). Wie andernorts verhält sich aber auch in Niederbayern der überwiegende Teil der Neophyten völlig unauffällig, nur ein kleiner Teil hier sind es 15 Arten (vgl.Zahlheimer2006) entpuppt sich als invasiv und/oder ökologisch problematisch (u.a. aufgrund von Verdrängung seltener heimischer Elemente, Stickstoffanreicherung im Boden durch Knöllchenbakterien, gesundheitlicher Beeinträchtigungen des Menschen oder genetischer Verfremdung durch Hybridisierung verwandter Taxa). NachKlingenstein(2004) ist in Deutschland bisher noch keine Art durch invasive Neobiota ausgelöscht worden und auch bei Pflanzenarten, die vom Aussterben bedroht sind, geht die Gefahr der Auslöschung nur in den wenigsten Fällen (ca. 5 %) tatsächlich von Neubürgern aus. Ihre grundsätzliche Ausrottung ist daher weder notwendig noch wünschenswert, schließlich stellen sie auch Glieder unserer Kulturlandschaft dar (Brandes2008).
Dennoch, invasive nicht einheimische Arten wurden als eine wichtige Ursache für das globale Artensterben erkannt (vgl.Böhmer2008, 2011, SCBD 2000) und nach Artikel 8h der Biodiversitätskonvention der UN-Konferenz für Umwelt und Entwicklung in Rio de Janeiro im Jahr 1992 sind alle Vertragsstaaten verpflichtet, Einfuhr und Ausbreitung gebietsfremder Taxa mit artspezifischen Maßnahmen soweit möglich entgegenzuwirken. Aufgrund einer meist außergewöhnlichen Robustheit der invasiven Neophyten sind Bekämpfungsmaßnahmen dabei oft über lange Zeiträume und unter hohem Kostenaufwand durchzuführen (Kowarik2010).
Etliche der invasiven Neophyten Deutschlands regenerieren sich aus verschleppten Rhizomstücken, weshalb Fließgewässer ideale Ausbreitungsvektoren darstellen (Schwabe&Kratochwil1991). Dies trifft auch auf den Unterlauf der Ilz zu, wo sich ausgedehnte Neophytenfluren im Uferbereich finden (Zahlheimer2001, Abb. 2). Gerade in diesen hochdynamischen Arealen ist eine Beobachtung der Bestandsentwicklung von großer Bedeutung, um Ausbreitungstendenzen erkennen und gegebenenfalls Gegenmaßnahmen einleiten zu können.
Die Neophytenvorkommen am Unterlauf der Ilz wurden 2009 hochaufgelöst kartiert und die artspezifischen Flächenanteile ermittelt (Stummer2009). 2015 wurde der Unterlauf der Ilz mit exakt gleicher Vorgehensweise erneut bearbeitet (Sompek2016). Ein Vergleich der beiden Erhebungen ist aus zweierlei Gründen interessant:
- Erstens wurden in dem zwischen den Erhebungen liegenden Zeitraum verschiedenerlei Bekämpfungsmaßnahmen durchgeführt, so dass sich über die Wiederholungsaufnahmen eventuell Aussagen zur Effizienz solcher Maßnahmen machen lassen. Diese wiederholt durchgeführten Maßnahmen konzentrieren sich auf die Ufer im Bereich des Gleithanges der westlichen Ilz-Schleife. Nachdem ein Mulchen mit Maschineneinsatz dort nicht möglich ist, werden die Neophyten von Hand entfernt, in Säcken gesammelt und anschließend abtransportiert.
- Ein Vergleich der beiden Kartierungen ist zum anderen interessant, da der Unterlauf der Ilz im Zeitraum zwischen den beiden Kartierungen wiederholt von kleineren, aber auch vom extremen Hochwasserereignis im Juni 2013 (vgl. auchFickert&Scheffer2013) betroffen war, wodurch es möglicherweise in Folge von Ufererosion zu einer verstärkten Verschleppung von Rhizomstücken und damit zu einer Ausbreitung der Neophyten gekommen sein könnte.
2 Naturräumliche Grundzüge des Untersuchungsraums
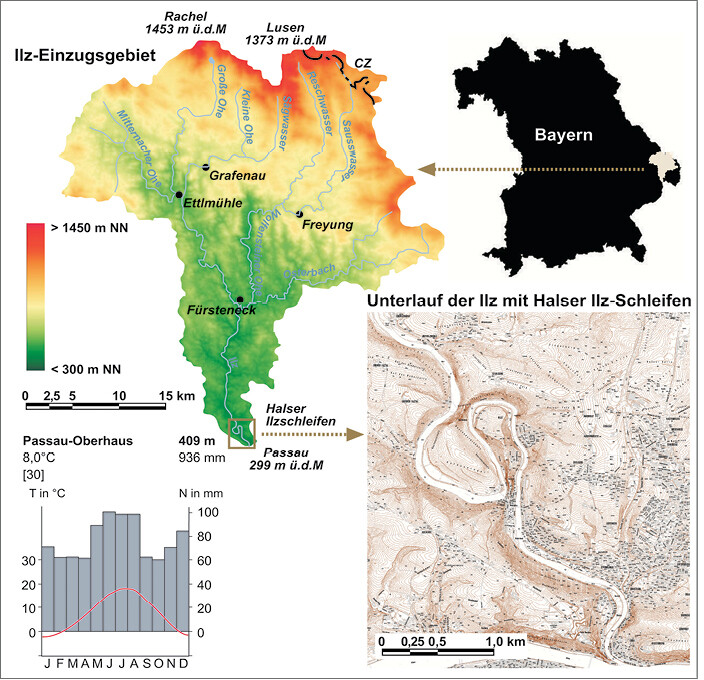
Die Ilz hat ihren Ursprung in verschiedenen Quellbächen in den Hochlagen des Bayerischen Waldes im Südosten Bayerns (Abb. 3). Im Stadtgebiet von Passau mündet sie bei 299 m ü.d.M. in die Donau. Bei einem Höhenunterschied von knapp 1 000 m fungiert die Ilz als bedeutende Migrationsachse zwischen dem milden Donautal und dem klimatisch rauen Mittelgebirge des Bayerischen Waldes, entlang derer auf der einen Seite thermophile Pflanzen- und Tierarten aus dem Donautal in Gunstlagen bis weit in den Bayerischen Wald vordringen können, auf der anderen Seite aber auch etliche hochmontane bis subalpine Arten aus dem hohen Bayerischen Wald in Schattenlagen noch weit unterhalb ihres angestammten Lebensraumes geeignete Lebensbedingungen finden (sieheButzet al. 2002,Fickert2013,Leibl2004,Zahlheimer2004).
Kurz vor ihrer Mündung in die Donau im Umfeld des Passauer Ortsteils Hals bildet die Ilz einen eigenwilligen doppelten Talmäander (Abb. 3), der von der Störungszone des Halser Nebenpfahls hervorgerufen wird. Da die hier auftretenden Pfahlschiefer (Mylonite) aufgrund tektonischer Beanspruchung leichter verwitterbar waren als die angrenzenden Kristallingesteine, bildete sich während des Miozän eine flache Mulde. Sie wurde später von wenig verfestigten Molassesedimenten erfüllt, in denen die Ur-Ilz in weiten Mäandern pendeln konnte. Bruchtektonische Hebung im Pliozän und Pleistozän als Fernwirkung der Alpenorogenese weiter südlich forcierte die Tiefenerosion und führte zu einer zunehmenden Eintiefung. Als schließlich die anstehenden Mylonite erreicht wurden, schnitt sich die Ilz unter Beibehaltung ihres vorher angelegten Flussverlaufs epigenetisch in die Tiefe und vererbte die ursprünglich freien Mäander auf das Festgestein (Eitel2002).
Klimatisch betrachtet ist der Unterlauf der Ilz als gemäßigt-humid zu bezeichnen, wobei sich in einer jahreszeitlichen Temperaturamplitude von über 20 K (Januar: -2,9 °C; Juli: 17,7 °C) und einem Niederschlagsmaximum in den Sommermonaten (Ergänzung der ganzjährig wirksamen advektiven Niederschläge durch konvektive Sommergewitter), bereits subkontinentale Klimazüge bemerkbar machen (vgl. Klimadiagramm in Abb. 3). Dem subkontinentalen Klimacharakter entsprechend finden sich in anthropogen wenig überprägten Hanglagen z.T. recht artenreiche Waldformationen mit Eichen ( Quercus petraea , Q. robur ), Hainbuchen ( Carpinus betulus ) und Buchen ( Fagus sylvatica ) sowie in geringeren Anteilen eine Vielzahl weiterer Edellaubhölzer (vgl.Fickert2009,2013;Zahlheimer2004). In den Uferbereichen ist an vielen Stellen die Banater Segge ( Carex buekii ) dominant vertreten, eine Rote-Liste-Art für Niederbayern (vgl.Zahlheimer2001). Aufgrund seiner relativen Naturnähe und seiner topographisch und ökologisch reichhaltigen Ausstattung wurde der Bereich der Halser Ilzschleifen (etwa die nördliche Hälfte des Untersuchungsgebietes) im Jahr 1993 als Naturschutzgebiet ausgewiesen.
3 Methode
Grundlage der Untersuchung sind in einem Abstand von sechs Jahren durchgeführte detaillierte Kartierungen der Neophytenbestände am Unterlauf der Ilz. Das Kartier-Areal erstreckt sich von der Mündung der Ilz in die Donau ca. 8 km flussaufwärts bis zur Stauanlage des Kraftwerks Oberilzmühle (Abb. 3). Die Kartierung beschränkt sich auf die unmittelbaren Uferbereiche des Fließgewässers, wo sich die ausgedehntesten Neophytenbestände befinden und wo aufgrund der hohen Flussdynamik mit Veränderungen zu rechnen ist. Die erste Kartierung erfolgte im Sommer 2009 bei ausgedehnten Begehungen (Stummer2009), die Wiederholung nach exakt gleicher Vorgehensweise im Jahr 2015 (Sompek2016).
Bei den Kartierungen wurde zwischen flächiger Verbreitung und Einzelvorkommen (Ausdehnung < 5 m) unterschieden. Die einzelnen Neophytenvorkommen wurden mit Hilfe von Maßbändern ausgemessen und per GPS lokalisiert, um Aussagen zu den Flächenanteilen der einzelnen Arten zu ermöglichen und deren Verbreitungsareale (re)lokalisieren zu können. Dabei wurde zwischen Reinbeständen und Mischbeständen unterschieden, zudem wurden für alle Arten Deckungsanteile (in % Bodenbedeckung) geschätzt, aus denen drei Deckungsklassen (Klasse 1: 1049,9 %; Klasse 2: 5074,9 %; Klasse 3: 75100 %) aggregiert wurden. Die im Gelände im Maßstab 1 : 1 000 kartierten Areale wurden im Anschluss in ArcGIS digitalisiert, um Flächenveränderungen zwischen den beiden Aufnahmezeitpunkten quantifizieren zu können (die erstellten Karten sind im Download-Bereich von Naturschutz und Landschaftsplanung unter www.nul-online.de , Webcode 2231, abrufbar). Neben den Neophyten wurde zudem die dominant uferbegleitende heimische Banater Segge ( Carex buekii ) kartiert, um beispielhaft Aussagen zu interspezifischen Licht- und Wurzelkonkurrenz zwischen nativen Arten und invasiven Neophyten treffen zu können. Die Nomenklatur der Arten folgt Haeupler&Muer(2007).
4 Flächenanteile und Bestandstruktur der kartierten Neophyten
Bei der ersten Erhebung 2009 wurden acht invasive Neophyten am Unterlauf der Ilz kartiert: drei verschiedene Staudenknötericharten (Sachalin-Staudenknöterich Fallopia sachalinensis , Japanischer Staudenknöterich Fallopia japonica und der aus diesen beiden Arten erst in Mitteleuropa durch Hybridisierung hervorgegangene Bastard-Staudenknöterich Fallopia x bohemica ), das aus Indien eingeschleppte Drüsige Springkraut ( Impatiens glandulifera ) sowie vier aus den Prärien Nordamerikas stammende Vertreter der Korbblütler: Kanadische und Späte Goldrute ( Solidago canadensis , S. gigantea ), Schlitzblättriger Sonnenhut ( Rudbeckia laciniata ) und Topinambur ( Helianthus tuberosus ).
Neben generativer Vermehrung sind alle genannten Neophyten in der Lage, sich aus verschleppten Rhizom- bzw. Sprossteilen zu regenerieren (letzteres z.B. bei Impatiens glandulifera , von dem abgerissene Pflanzenteile an Stängelknoten wieder wurzeln können, wenngleich diese vegetative Form der Ausbreitung gegenüber einer Ausbreitung über Samen untergeordnet bleibt, vgl.Ludwiget al. 2000). Die Anlandung abgerissener Pflanzenfragmente erfolgt im Bereich der Halser Ilzschleifen bevorzugt an den Innenseiten der Mäander mit geringerer Fließgeschwindigkeit, wo sich besonders großflächige Neophytenbestände finden, während an den Außenbögen der Mäander bei deutlich höherer Fließgeschwindigkeit und fortwährender Seitenerosion eine Etablierung erschwert ist (vgl. Kartierungen 2009 und 2015 im Downloadbereich von NuL).
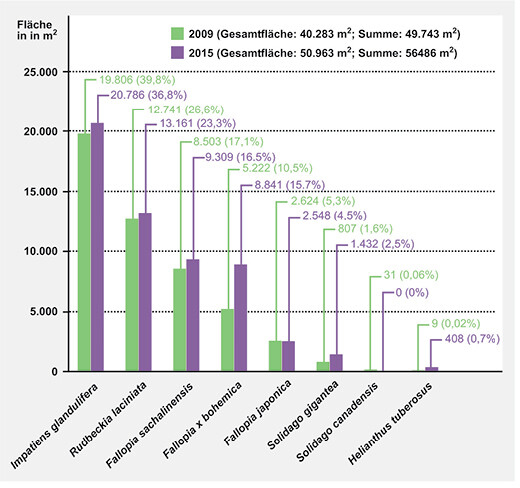
Die Neophytenkartierung aus dem Jahr 2009 (Stummer2009) erbrachte eine von Neophyten bestandene Gesamtfläche von 40 283 m², das ist etwa ein Sechstel der gesamten Uferfläche im kartierten Bereich (ca. 0,25 km2). Abb. 4 zeigt die Anteile der einzelnen Arten an der gesamten Neophytenfläche. Da es sich zum Teil um Mischbestände handelt, liegen die kumulierten Werte der einzelnen Arten mit 49 743 m² etwas höher als die von Neophyten bestandene Gesamtfläche. Impatiens glandulifera nahm 2009 mit ca. 40 % die größte Fläche ein, gefolgt von Rudbeckia laciniata mit gut 25 %. Etwa ein Drittel der Fläche beanspruchten die drei verschiedenen Staudenknöterich-Arten, wobei Fallopia sachalinensis den größten Anteil stellte. Relativ geringe Anteile von weniger als 2 % der Gesamtflächen nahmen Helianthus tuberosus , Solidago gigantea und Solidago canadensis ein.
Die Wiederholungs-Kartierung im Juli 2015 (Sompek2016) erbrachte eine deutliche Zunahme der von Neophyten bestandenen Fläche. Insgesamt hat sich die Gesamtfläche der Neophyten auf ca. 51 000 m2(das entspricht etwa einem Fünftel der gesamten Uferfläche) und damit um etwa ein Viertel (26,5 %) binnen sechs Jahren erhöht. Die aufsummierten Deckungsanteile der einzelnen Arten belaufen sich auf 56 486 m² (gegenüber 49 743 m² im Jahr 2009). Mit Ausnahme von Solidago canadensis , die bei der Kartierung im Jahr 2015 nicht mehr angetroffen wurde, und Fallopia japonica , die ein minimal kleineres Areal bedeckt, haben alle anderen Neophyten an Fläche dazugewonnen, wobei sich die Zugewinne bei den einzelnen Arten unterschiedlich darstellen (Abb. 4). Die relativ höchsten Zugewinne verzeichnen Fallopia x bohemica und Solidago gigantea mit nahezu verdoppelten Werten gegenüber 2009 sowie Helianthus tuberosus mit einer exponentiellen Zunahme von 9 m2auf 408 m2(Abb. 4). Impatiens glandulifera , Rudbeckia laciniata und Fallopia sachalinensis haben flächenmäßig leicht zugenommen, ihr prozentualer Anteil an der gesamten von Neophyten bestanden Fläche hat jedoch gegenüber 2009 leicht abgenommen.
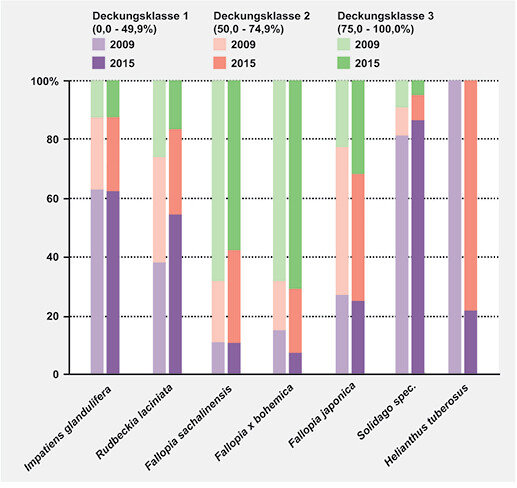
Die einzelnen Neophytenbestände unterscheiden sich hinsichtlich ihrer Struktur deutlich (vgl. Abb. 5). Während die beiden Staudenknöteriche Fallopia sachalinensis und F. x bohemica überwiegend sehr dichte Bestände bilden (zumeist Klasse 3 mit Deckung > 75 %, Abb. 5), weisen die Bestände der übrigen Neophyten überwiegend geringe bis mittlere Deckungswerte auf (v.a. Deckungsklasse 1 und 2, Abb. 5). Zwischen den beiden Erhebungen zeigen sich keine grundlegenden Unterschiede, lediglich bei Helianthus tuberosus zeichnet sich neben der starken Ausbreitung (siehe oben) zugleich eine Verdichtung der Bestände gegenüber 2009 ab, als die wenigen Vorkommen des Topinambur durchwegs sehr lockeren Wuchs aufwiesen.
5 Ursachen und Auswirkungen der Bestandsentwicklung sowie mögliche Gegenmaßnahmen
Wie im Falle anderer deutscher Fließgewässer dürfte die Ausweitung der Gesamtfläche von Neophyten am Unterlauf der Ilz mit großer Wahrscheinlichkeit auf das Verschleppen von Rhizomstücken zurückzuführen sein. Während Impatiens glandulifera zwar auch große Mengen an ballochoren (d.h. mit Schleudermechanismus versehenen) und vom Wasser verschleppten Samen produziert, die sicher zur Expansion der Art beitragen, überwiegt insbesondere bei Fallopia -Arten (vgl.Böhmeret al. 2006) und bei Helianthus tuberosus die vegetative Ausbreitung deutlich gegenüber der generativen. Nachdem Bauaktivitäten im Uferbereich (mit Ausnahme der Neugestaltung des Bschütt-Areals unmittelbar vor der Mündung) im entsprechenden Zeitraum nicht stattgefunden haben und bei den Bekämpfungsaktionen dafür Sorge getragen wurde, dass keine Rhizom- oder Pflanzenteile ins Wasser gelangen, ist davon auszugehen, dass Ufererosion in Zusammenhang mit gelegentlich auftretenden kleineren Hochwasserereignissen, insbesondere aber mit dem massiven Hochwasser im Juni 2013, für die Ausbreitung der Rhizomstücke verantwortlich ist.
Die am Unterlauf der Ilz auftretenden Neophyten sind sehr robuste und potente Pflanzen, die nach ihrer Etablierung Struktur und Artenzusammensetzung betroffener Uferbereich nachhaltig verändern und alteingesessene Pflanzen verdrängen können.Böhmeret al. (2006) haben dies z.B. für Fallopia japonica eindrücklich aufgezeigt. Auch am Unterlauf der Ilz lassen sich vergleichbare Effekte durch Staudenknöterich-Arten erkennen, insbesondere auf die Bestände der im Bayerischen Wald eher seltenen, jedoch am Unterlauf der Ilz durchaus weit verbreiteten Banater Segge ( Carex buekii ).
Bereits bei der ersten Kartierung wurden vonStummer(2009) keine Mischbestände der Segge mit Fallopia -Arten angetroffen. Ein Vergleich der beiden Kartierungen belegt zudem, dass Carex buekii -Bestände durch die Expansion der Staudenknöteriche teilweise unterbrochen bzw. ganz verdrängt wurden (Stummer2009,Sompek2016). Die Verdrängung der Segge durch die Fallopia -Arten ist kritisch zu sehen. Während die Segge als mehrjährige, mächtige Horste bildende Art die Ufer nachhaltig vor Ufererosion schützt, ist ein solcher Schutz in jenen Abschnitten nicht gegeben, wo die Segge von den einjährigen und im Winter bis auf die Rhizome zurücksterbenden Knöterichen verdrängt wurde. Da sich aufgrund der massiven Beschattung während der Sommermonate keine sonstige Unterwuchsvegetation bilden kann, fehlt ein effektiver Erosionsschutz für die Uferbereiche nach dem Absterben der Knöteriche (vgl. auchBöhmeret al. 2006). So sind die von Fallopia dominierten Uferabschnitte in den Wintermonaten mehr oder weniger unbewachsen der Erosion ausgesetzt, wodurch gleichzeitig ein weiteres Verschleppen von Rhizomstücken der Fallopia -Arten begünstigt wird.
Trotz lokaler Verdrängung von Carex buekii durch die Fallopia -Arten hat sich die Bestandsfläche der Segge im Zeitraum zwischen den beiden Kartierungen von ca. 1,8 auf gut 2 ha sogar leicht erhöht, was belegt, dass Carex buekii den Neophyten im Unterlauf der Ilz nicht generell unterlegen ist. Mit Impatiens glandulifera bildet die Segge beispielsweise weitverbreitet Mischbestände, die scheinbar in Gleichgewicht stehen (Sompek2016). Ähnliches berichtenEssl&Hauser(2003) aus dem niederösterreichischen Thaya-Tal. Die Konkurrenzstärke invasiver Neophyten stellt sich demnach artspezifisch unterschiedlich dar (vgl. auchBöhmer2011). Dies wird auch deutlich, wenn man die Konkurrenz der Neophyten untereinander betrachtet. So zeigt sich beispielsweise, dass Rudbeckia laciniata in Mischbeständen mit anderen Neophyten offenbar unterlegen ist. Die Wiederholungskartierung von 2015 ergab, dass der Sonnenhut von den Staudenknöterichen lokal verdrängt wurde. Insbesondere Fallopia sachalinensis erwies sich an einem Standort, an dem 2009 noch ein Mischbestand mit Rudbeckia laciniata und Impatiens glandulifera zu finden war, als besonders konkurrenzstark und hat im Zeitraum von sechs Jahren diese Fläche praktisch gänzlich vereinnahmt (Abb. 6). Gleiches gilt für im Jahr 2009 kartierte Mischbestände aus Springkraut ( Impatiens glandulifera ) und Rudbeckia laciniata (Stummer2009), die 2015 ausschließlich von Impatiens glandulifera bestanden waren (Sompek2016). Insgesamt erscheinen die Staudenknöteriche das höchste Durchsetzungsvermögen aller hier auftretenden Neophyten zu besitzen, gefolgt vom Springkraut.
6 Management
Grundsätzlich sollten Neophyten nicht unter Generalverdacht gestellt werden. Sicher verhalten sich einige von ihnen invasiv, das tun aber etliche einheimische Pflanzen auch, insbesondere dort, wo die Nutzung aufgegeben wurde. Dort können bestimmte heimische Arten überhandnehmen und alteingesessene, z.T. seltene Pflanzen auf ehemals extensiv genutzten Flächen verdrängen. In Niederbayern zählen z.B. Pioniergehölze wie die Schlehe ( Prunus spinosa ) oder der Liguster ( Ligustrum vulgare ), Kletterpflanzen wie Brombeeren ( Rubus spec . ), Waldreben ( Clematis spec . ) und Efeu ( Hedera helix ), aber auch Kräuter wie Brennnessel ( Urtica dioica ) und verschiedene Gräser zu diesen besonders durchsetzungsfähigen heimischen Pflanzen (vgl.Zahlheimer2001). Wie die meisten einheimischen Pflanzen verhalten sich auch die meisten Neophyten völlig unauffällig und gliedern sich in die heimische Flora ein, ohne Probleme zu verursachen.Kowarik(2010) führt sogar einige positive Aspekte in Zusammenhang mit den Neophyten an, etwa ihre Funktion als Bienenweide im blütenarmen Spätsommer, ihre visuelle Attraktivität und ihre kulturhistorische Bedeutung.
Nachdem insbesondere die Fallopia -Arten und auch Impatiens glandulifera als besonders konkurrenzstark und z.T. heimische Elemente verdrängend erkannt wurden, sind Bekämpfungsmaßnahmen für diese Arten in Betracht zu ziehen. Solche Maßnahmen können von den Naturschutzbehörden veranlasst werden (vgl.Schepker2004), oftmals sind es aber auch private Initiativen oder Vereine, die hier tätig werden. Am Unterlauf der Ilz hat beispielsweise der Bayerische Wald-Verein wiederholt Aktionen (1-2 pro Jahr) zur Bekämpfung besonders unliebsamer Neophyten (v.a. Impatiens glandulifera und die drei Fallopia -Arten) durchgeführt, bei denen die Pflanzen von Hand entfernt, gesammelt und abtransportiert werden. Während beim Indischen Springkraut durchaus eine Eindämmung mit diesen Maßnahmen festzustellen ist, bleibt der Erfolg bei den Staudenknöterichen mit ihrer Fähigkeit zum Wieder-Austrieb aus Rhizomstücken gering.
Für eine langfristige und nachhaltige Zurückdrängung der Fallopia -Arten an der Ilz muss über andere Strategien nachgedacht werden. Total-Herbizide wie Glyphosat kommen für die ufernahen Bestände an der Ilz nicht in Frage. Um Gewässerkontamination zu vermeiden, müssen bei einem Einsatz von Herbiziden mindestens 5-10 m Entfernung bis zum Fließgewässer eigehalten werden, was aber bei den meisten Fallopia -Beständen an der Ilz nicht gegeben ist. Abdunkelung durch Folien hat sich bei Fallopia -Arten als wenig effektiv herausgestellt, ist doch Fallopia japonica beispielsweise in der Lage, selbst 5 cm dicken Asphalt zu durchstoßen (vgl.Böhmeret al. 2006). Hinzu kommt, dass großflächiges Ausbringen von Folien die gesamte Vegetation im abgedunkelten Bereich absterben lässt, wodurch die Gefahr von Bodenerosion steigt. Als erfolgversprechend haben sich andernorts mehrfache Beweidung im Jahr durch Schafe herausgestellt, ebenso wie häufige Mahd und gezieltes Schlegeln, wodurch die Konkurrenzkraft der Neophyten nachlässt und heimische Hochstauden aufkommen können (Böhmeret al. 2016). Da Mahd allerdings nicht selektiv erfolgen kann, werden durch die hohe Wiederholungsfrequenz der Maßnahme auch eigentlich erwünschte Pflanzen geschwächt. Pflanzmaßnahmen und gezielte Förderung indigener Arten haben sich oft als wenig erfolgreich herausgestellt, da die meisten der Licht- und Wurzelkonkurrenz der Neophyten unterliegen. Untersuchungen vonSchepker(1998) undSchlüpmann(2000) zeigen, dass z.B. der Straußfarn ( Matteuccia struthiopteris ), der auch am Unterlauf der Ilz auftritt, Fallopia japonica Paroli bieten kann. Auch Carex buekii scheint sich, abgesehen von den Staudenknöterichen, gegen die übrigen Neophyten behaupten zu können. Weitere konkurrenzstarke indigene Arten scheinen das Rohrglanzgras ( Pahalaris arundinacea ), Schilf ( Phragmites communis ), Pestwurz-Arten ( Petasites spec.) und die Schwarzrele ( Alnus glutinosa ) zu sein.
Literatur
Böhmer, H.J. (2008): Biologische Invasionen lokales Problem oder globale Herausforderung. Natur und Landschaft 83 (9/10), 394-398.
(2011): Biologische Invasionen Muster, Prozesse und Bioglobalisierung. Geogr. Rundschau 63 (3), 4-10.
, Heger, T.,Alberternst, B.,Walser, B. (2006): Ökologie, Ausbreitung und Bekämpfung des Japanischen Staudenknöterich (Fallopia japonica ) in Deutschland. Anliegen Natur 30, 29-34.
Brandes, D. (2008): Invasive Pflanzen: Naturkatastrophen oder Spiegel unserer Kulturgeschichte. In: Abhandlungen der Braunschweigischen Wissenschaftlichen Gesellschaft 59, 9-36.
Butz, L.,Eden, D.,Feuchtgruber, J.(2002): Die Ilz Flusslandschaft der Jahre 2002 / 2003. Der Bayerische Wald 16 (1 & 2) NF, 4-7.
Eitel, B. (2002): Flächensystem und Talbild im östlichen Bayerischen Wald (Großraum Passau-Freyung). In:Ratusny, A., Hrsg., Flusslandschaften an Inn und Donau, Passauer Kontaktstudium Erdkunde 6, 19-34.
Essl, F.,Hauser, E. (2003): Verbreitung, Lebensraumbindung und Managementkonzept ausgewählter invasiver Neophyten im Nationalpark Thayatal und Umgebung (Österreich). Linzer biol. Beitr. 35 (1), 75-101.
Fickert, T. (2009): Geoökologische Untersuchungen im Bereich der Halser Ilzschleifen. Der Bayerische Wald 22 (1+2) NF, 29-41.
(2013): Das Ilztal Natur- und Kulturlandschaftsentwicklung an der schwarzen Perle des Bayerischen Waldes. In:Gamerith, W.Anhuf, D.,Struck, E.,Hrsg., Passau und seine Nachbarregionen; Orte, Ereignisse, Verbindungen ein geographischer Wegweiser, 64-77.
, Scheffer, J.(2013): Jahrtausendhochwasser 2013 in Passau. Geogr. Rundschau 65 (9), 61-65.
Haeupler, H.,Muer, T.(2007): Bildatlas der Farn- und Blütenpflanzen Deutschlands. Ulmer, Stuttgart, 789 S.
Klingenstein, F. (2004): Neophyten aus Sicht des Naturschutzes auf Bundesebene. In: Neophyten in Schleswig-Holstein: Problem oder Bereicherung? Dokumentation einer Tagung im LANU am 31.03.2004, Schr.-R. LANU SH, 21-32 www.floraweb.de/neoflora/neophyten_naturschutz_deutschland.pdf.
Kowarik, I. (2003): Biologische Invasionen durch nichteinheimische Pflanzenarten. In: Leibniz-Institut für Länderkunde, Hrsg., Nationalatlas Bundesrepublik Deutschland, Bd. 3: Klima, Pflanzen- und Tierwelt, 110-111.
(2010): Biologische Invasionen Neophyten und Neozoen in Mitteleuropa. Ulmer, Stuttgart, 362 S.
Leibl, F. (2004): Die Bedeutung der Ilz für die Fauna Niederbayerns. In: Der Bayerische Wald 18 (1) NF, 18-20.
Ludwig, M.,Gebhardt, H.,Ludwig, H.W.,Schmidt-Fischer, S.(2000): Neue Tiere und Pflanzen in der heimischen Natur. BLV, München, 127 S.
Nentwig, W. (2010): Invasive Arten. UTB Profile, Haupt, 128 S.
Petrischak, H. (2014): Neophyten in Mitteleuropa Flucht aus dem Garten: Die Erfolgsgeschichte invasiver Pflanzenarten. Biologie in unserer Zeit 44, 410-419.
SCBD (Secretariat of the Convention on Biological Diversity, 2000): Sustaining life on Earth. How the Convention on Biological Diversity promotes nature and human well-being. Montreal.
Schepker, H. (1998): Wahrnehmung, Ausbreitung und Bewertung von Neophyten. Eine Analyse der problematischen nichteinheimischen Pflanzenarten in Niedersachsen. Stuttgart.
(2004): Problematische Neophyten in Deutschland Ergebnisse einer bundesweiten Befragung von Naturschutzbehörden unter besonderer Berücksichtigung Schleswig-Holsteins. In: Landesamt für Natur und Umwelt des Landes Schleswig-Holsteins, Hrsg., Neophyten in Schleswig-Holstein: Problem oder Bereicherung? Kiel, 39-50.
Schlüpmann, M. (2000): Zur Neophyten-Flora der Volmeaue im Hagener Stadtgebiet. Decheniana 153, 37-49.
Schwabe, A.,Kratochwil, A.(1991): Gewässerbegleitende Neophyten und ihre Beurteilung aus Naturschutzsicht unter besonderer Berücksichtigung Südwestdeutschlands. NNA-Ber. 4 (1), 15-27.
Sompek, E. (2016): Neophyten am Unterlauf der Ilz. Unveröff. Zulassungsarb., Lehrstuhl für Physische Geographie, Univ. Passau, 111 S.
Stummer, A. (2009): Neophytenpflanzen in Deutschland eine regionalspezifische Bestandsaufnahme potentiell invasiver Arten am Unterlauf der Ilz. Unveröff. Zulassungsarb., Lehrstuhl für Physische Geographie, Univ. Passau, 99 S.
Zahlheimer, W.A. (2001): Die Farn- und Blütenpflanzen Niederbayerns, ihre Gefährdung und Schutzbedürftigkeit mit Erfassung einer Roten Liste. Hoppea 62, 5-347.
(2004): Die Pflanzenwelt an der Ilz Anforderungen aus der Sicht des Naturschutzes. Der Bayerische Wald 18 (1) NF, 21-28.
(2006): Pflanzenwelt Niederbayern Wachsende Gefahr unduldsame Gewächse. flora-niederbayern.de/unduldsame.htm.
Fazit für die Praxis
Kontakt
Elisabeth Sompek beginnt nach Abschluss ihres Lehramtsstudiums mit den Fächern Geographie und Wirtschaft an der Universität Passau (2011 bis 2017) in Kürze ihr Referentariat. Ihre Zulassungsarbeit verfasste sie 2016 zur Neophytenverbreitung am Unterlauf der Ilz.
Andreas Stummer arbeitet als Lehrer an der Realschule Arnstorf. Lehramtsstudium mit den Fächern Geographie und Englisch an der Universität Passau (2004-2010), Zulassungsarbeit zur Neophytenverbreitung am Unterlauf der Ilz (2009).
PD Dr. Thomas Fickert ist Akademischer Oberrat auf Zeit am Lehrstuhl für Physische Geographie im Fachbereich Geographie der Universität Passau. 1989 bis 1997 Diplom-Studium der Physischen Geographie (Nebenfächer Geologie, Biologie) und 2004 Promotion an der Friedrich-Alexander-Universität Erlangen-Nürnberg). 2014 Habilitation an der Universität Passau. Arbeitsgebiete: Vegetationsgeographie mit Schwerpunkt Störungsökologie, Sukzession und Phytoindikation; Mangroven; Ökologie der ariden und mediterranen Subtropen, Hochgebirgsforschung, Geomorphologie.
> thomas.fickert@uni-passau.de > www.phil.uni-passau.de/fachbereich-geographie/team/thomas-fickert/
Christian Kern arbeitet als Fachkraft für Netztdatenservice in München. Von 2004 bis 2010 Studium der Kartographie und Geomedientechnik an der Hochschule München (Dipl.-Ing. FH), 2012 bis 2014 an derselben Hochschule Studium in Printmedien, Technologie und Management (M. Eng.).
-
Abb. 2: Flächig ausgebildeter Bestand des Sachalin-Staudenknöterichs (Fallopia sachalinensis ) an der Ilz. Extensive stands of Sakhalin knotweed (Fallopia sachalinensis) along the Ilz River.Thomas Fickert -
Abb. 1: Sippenbilanz der Flora Deutschlands (nach Klingenstein 2004). Balance of clans of the Flora of Germany.Autoren -
Abb. 3: Lage des Untersuchungsgebietes im Südosten Bayerns und innerhalb des Ilz-Talsystems. Location of the study area in southeastern Bavaria and within the Ilz river system (elevation model acc. to SRTM data, mapping unit provided by Bayerisches Landesamt für Vermessung und Geoinformation).Höhenmodell nach SRTM-Daten, Kartenausschnitt des Unterlaufs der Ilz bereitgestellt vom Bayerischen Landesamt für Vermessung und Geoinformation -
Abb. 4: Flächenanteile der einzelnen Neophyten am Unterlauf der Ilz in m2und in Prozent für die beiden Erhebungszeitpunkte 2009 und 2015. Da es sich zum Teil um Mischbestände handelt ist die Summe der Deckungsanteile größer als die Gesamtfläche. Area coverd by the different neophytes (in m² and %) along the lower course of the Ilz River for the two different sampling dates (2009 and 2015). Note: As there are some mixed stands the sum of cover values is larger than the total ground cover of neophytes.Autoren -
Abb. 5: Veränderung der Anteile der einzelnen Arten an den Deckungsklassen 1, 2 und 3 zwischen 2009 und 2015. Change in the share of cover classes for the different neophytes between 2009 and 2015.Autoren -
Abb. 6: Foto-Vergleich eines Neophyten-Bestandes am sog. Stockbauersteg: 2009 (oben) handelte es sich noch um einen Mischbestand ausImpatiens glandulifera ,Rudbeckia laciniata undFallopia sachalinensis , 2015 (unten) wird derselbe Standort fast ausschließlich vonFallopia sachalinensis eingenommen. | Comparison of an extensive neophyte stand next to the so-called Stockbauer-bridge. In 2009 the vegetation was mixed stand of Impatiens glandulifera, Rudbeckia laciniata and Fallopia sachalinensis, in 2015 the same site was covered almost exclusively by Fallopia sachalinensis.Andreas Summer (2009), Sompek (2015) -
Elisabeth Stompekprivat -
Andreas Stummerprivat -
PD Dr. Thomas Fickertprivat -
Christian Kernprivat


Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.