Auswirkungen der Offshore-Windenergie auf Seevögel, Fische und Benthos
Abstracts
Der Beitrag liefert eine synoptische Zwischenbetrachtung der bau-, anlage- und betriebsbedingten Auswirkungen von Offshore-Windenergie auf die drei Tiergruppen Seevögel, Fische und Benthos. Im Zuge einer qualitativen Literaturanalyse wurden ca. 150 internationale, insbesondere europäische Publikationen verschiedenen Status bis zum Jahr 2015 ausgewertet. Es werden die (artspezifischen) Wirkungshypothesen aus den Quellen kategorisiert und tabellarisch gegenübergestellt. Nach mehr als einer Dekade intensiver Untersuchungen zeigt sich ein erstes Verständnis, welche Tierarten zu den vulnerablen gegenüber Kollision, Störung oder Verdrängung zählen, u.a. in Abhängigkeit von ihrer Morphologie, der Phänologie, anlage- oder betriebsbedingten Effekten von Offshore-Windparks sowie entsprechenden Verhaltensreaktionen. Infolge großer methodischer Herausforderungen auf dem Meer (z.B. für die Erhebung von Kollisionsopferraten) bleibt es jedoch anspruchsvoll, kausale Effekte, Vermeidungsverhalten oder gar Gewöhnungseffekte näher zu verstehen und v.a. nachzuweisen. So stellen etwa Sekundärwirkungen auf Fische und Seevögel durch den künstlichen Riffeffekt und veränderte Nahrungsgrundlagen neben den Fragen kumulativer Wirkungen und populationsbezogener Aussagen eine Langfristaufgabe dar. Auch bedarf es u.a. eines übergreifenden Monitorings bzw. Meta-Analysen, um den raumzeitlichen Variationen mit einem zunehmenden Zubau von Offshore-Windparks Rechnung tragen zu können.
Effects of offshore wind farms on sea birds, fish and benthos Synopsis of current professional literature
The paper provides a synoptical review of impacts related to the construction and operation of offshore wind farms on sea birds, fish and benthos. In conducting a qualitative literature review, approximately 150 primarily European publications up to the year 2015 were examined. Primarily species-specific hypotheses were categorized and compared.
After more than a decade of intensive study, an initial understanding has been developed regarding vulnerable species and collision risk, disturbance or displacement, relative importance of morphology or phenology, as well as operation related effects of offshore wind farms and changes in respective behavior, to name but a few. Due to greater methodological difficulties at sea, e.g. the collection of fatality data, better understanding and evidence of causal effects, including avoidance behavior or even habituation, remain challenging. It is a long-term task, to identify secondary impacts such as the artificial reef-effect on fish and sea birds and varying distributions of food resources, as well as cumulative effects and effects at the population level. Additionally, there is a need for comprehensive monitoring and meta-analyses, in order to stay abreast of spatio-temporal variations occurring alongside the massive build-up of offshore wind energy.
- Veröffentlicht am

Bird counting at sea.Graeme Pegram
1 Einleitung
In der letzten Dekade konnte nicht nur ein merklicher Wissenszuwachs zur Windenergie an Land, sondern auch zur Offshore-Windenergie und ihren Auswirkungen auf die wildlebende Fauna verzeichnet werden (Schuster et al. 2015). Dabei liegt ein Fokus auf Tiergruppen wie marinen Säugern (z.B. Brandt et al. 2011, Brasseur et al. 2012, Russel et al. 2015, Scheidat et al. 2012) und Zugvögeln (z.B. Aumüller et al. 2011, 2013; Bellebaum et al. 2010, Blew et al. 2013, Hill 2015, Hüppop et al. 2006). Seevögel, Fische und Benthos-Gemeinschaften wiederum stehen im Mittelpunkt der vorliegenden Synopse. Insbesondere eine Reihe europäischer Forschungsvorhaben sowie damit einhergehende Konferenzen und Publikationen erlauben eine solche synoptische Zwischenbetrachtung:
Conference on Wind Energy and Wildlife Impacts (CWW 2011, Trondheim),
Offshore Wind and Ecology Congress (OWE 2012, Amsterdam),
Conference on Wind Power and Environmental Impacts (CWE 2013, Stockholm),
Environmental Impact of Offshore Wind Farms (WinMon.BE 2013 mit Fokus auf die Windparks Bligh Bank und Thorntonbank, Brüssel),
Ökologische Begleitforschung am Offshore-Testfeldvorhaben alpha ventus (StUK plus 2013/2014, Berlin),
Conference on Wind Energy and Wildlife Impacts (CWW 2015, Berlin).
Dabei wurden auch Monitoring-Programme gestartet, etwa in Schottland das Marine Environmental Monitoring Programme (MEMP) und betreffende Analysen der Bau- und Betriebsphasen von Offshore-Windenergieanlagen sowie Maßnahmen, die auf das Standardkonzept Untersuchung der Auswirkungen von Offshore-Windenergieanlagen auf die Meeresumwelt (kurz StUK) des Bundesamtes für Seeschifffahrt und Hydrographie (BSH) für die deutsche ausschließliche Wirtschaftszone (AWZ) aufbauen.
Ziel dieser Synopse ist es, den Stand des Wissens über Auswirkungen der Offshore-Windenergie auf Seevögel (Abb. 1), Fische und Benthos kompakt zu bündeln und so die von Schuster et al. (2015) zusammengestellten Auswirkungen auf Zugvögel und marine Säuger zu ergänzen. Auswirkungen auf das marine Landschaftsbild bleiben hier unbeachtet. Die Erfassung art-, standort-, jahreszeitbedingter o.ä. Verhältnisse und Auswirkungen im konkreten Einzelfall sind von diesem synoptischen Überblick selbstverständlich unbenommen.
2 Methode
Der Synopse liegt eine systematische, qualitative Literaturanalyse zugrunde. Insgesamt wurden ca. 150 internationale Publikationen verschiedenen Status ausgewertet, darunter v.a. Fachartikel mit Peer-Review (1), Synthesen mit Peer-Review (2), andere Zusammenfassungen (3), Studien/Berichte bzw. graue Literatur (4), Buchbeiträge (5) und Konferenzbeiträge (6) bis 2015 und in Tabellen gemäß Schuster et al. (2015) kategorisiert. Die meisten Quellen stammen gemäß der Entwicklung der Offshore-Windenergie aus Europa, darunter Dänemark, Deutschland, die Niederlande, Belgien, Großbritannien, Norwegen, Schweden, Spanien und die USA im Gegensatz zur Windenergie an Land verläuft die Entwicklung auf dem Meer in den USA allerdings verzögert. Die Auswahl und Auswertung von Publikationen erfolgten für bestimmte Familien, Klassen und z.T. einzelne Arten der ausgewählten Tiergruppen.
Seevögel oder Meeresvögel (zusammenfassend kurz Seevögel) meinen Ordnungen, Familien bzw. Arten, die in ihrer Lebensweise vorwiegend an den Küsten- und/oder Meereslebensraum angepasst sind. Sie begeben sich an den Küsten, auf oder über dem offenen Meer auf Nahrungssuche und ruhen, brüten und ziehen ihre Jungen dort groß. Zu den Wasservögeln gehörend, werden sie gemäß ihres Verhaltens bzw. ihrer Nahrungssuche in Schwimm- und Tauchvögel sowie in zumeist fliegende Seevögel wie u.a. Seeschwalben, Sturmvögel und wenig spezialisierte Familien wie Möwen unterschieden (Peterson et al. 1961, Sauer 1982). Folgende Familien der Seevögel fanden hier Berücksichtigung (Ordnung in Klammern; Bezzel & Prinzinger 1990, Sauer 1982, Svensson et al. 2011):
Entenvögel (Gänsevögel: Anatidae),
Tölpel, Kormorane (Ruderfüßer: Pelecaniformes),
Sturmschwalben, Sturmvögel, Tauchsturmvögel (Röhrennasen: Procellariiformes),
See- und Lappentaucher (Seetaucher: Gaviiformes; Lappentaucher: Podicipediformes),
Raubmöwen, Möwen, Alken, Seeschwalben (Watvögel, Möwen und Alken: Charadriiformes).
Dabei müssen nicht alle Gruppen oder Arten einer Familie (z.B. Entenvögel Schwimm-, Tauch-, Eider-, Meer-, Ruderenten und Säger; Sauer 1982) zu den Seevögeln gehören, was die Abgrenzung bisweilen erschwert. Für einige Familien oder Gattungen wie Austernfischer und Wassertreter wurden keine hinreichenden Publikationen gefunden; Albatrosse, Pelikane und Fregattvögel blieben auf Grund ihrer geographischen Verbreitung unberücksichtigt. Auswirkungen auf aktiv ziehende (Sing-)Vogelarten werden hier bis auf einige Ausnahmen (z.B. Teilstreckenzieher wie bestimmte Schwimmenten) zum Großteil vernachlässigt (berücksichtigt sind sie bei z.B. Blew et al. 2013, Hill 2015 und Hill et al. 2014).
In der Tiergruppe der Fische waren folgende (Teil-)Klassen und/oder Arten in den Publikationen relevant und wurden entsprechend berücksichtigt:
Echte Knochenfische (Teleostei) bzw. Süßwasserfische,
Plattenkiemer (Elasmobranchii): Unterklasse der Knorpelfische.
Des Weiteren wurden Arten folgender Kategorien untersucht:
demersale Arten (Grundfischarten): Fische, die nahe am oder auf dem Meeresgrund leben,
pelagische Arten: Fische, die im offenen Meer zwischen der Wasseroberfläche und dem Meeresgrund leben,
Raubfische und
ziehende Arten.
Auch wurden verschiedene Lebenszyklen oder -stadien in einigen Publikationen betrachtet, so z.B. im Hinblick auf die Laichzeit.
Beim Benthos konnten in der Analyse folgende Artengruppen berücksichtigt werden:
mobile Wirbellose, z.B. Weichtiere (Mollusca wie Muscheln) und Flohkrebse (Amphiphoda),
demersale (Mega-)Fauna, z.B. Krabben, sowie teilweise
Nesseltiere, z.B. Blumentiere (Anthozoa) und
Flora, z.B. Seegras und Meeresalgen.
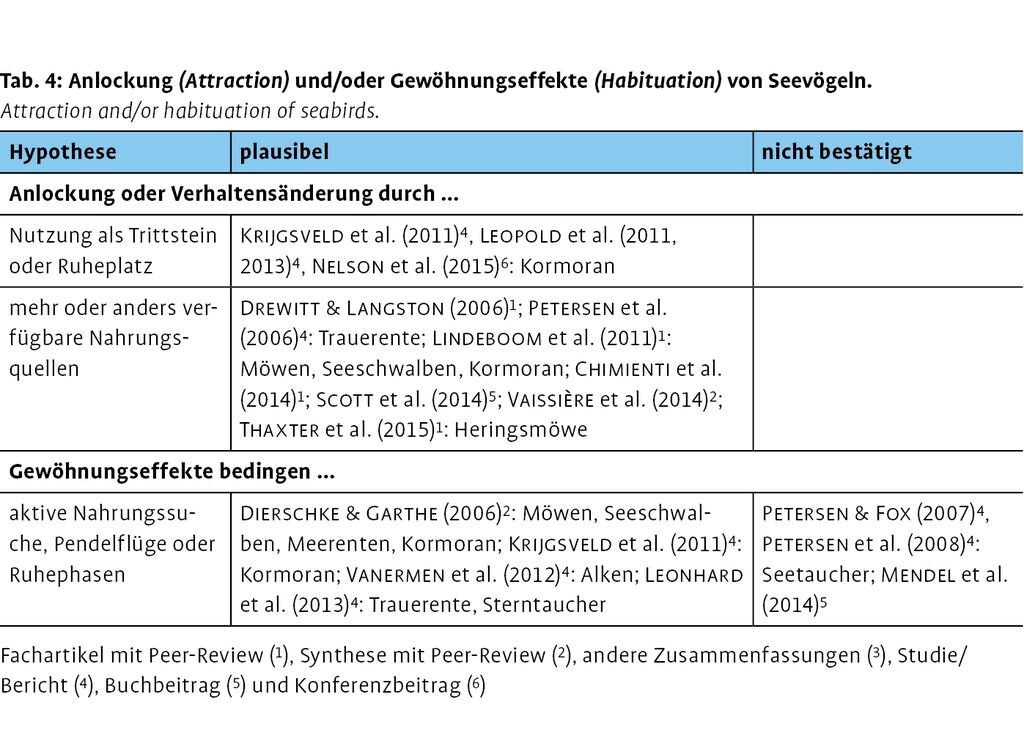
Nicht alle Auswirkungen werden im Detail (Abschnitt 3.1.1 bis 3.1.3 für Seevögel, 3.2.1 bis 3.2.3 für Benthos, 3.3.1 und 3.3.2 für Fische) durchweg für plausibel oder übertragbar erachtet oder stringent bestätigt und von verschiedenen Autorinnen und Autoren auch kontrovers diskutiert. Dies wurde im Zuge der Literaturanalyse berücksichtigt; es wurden Wirkungshypothesen aus den Quellen identifiziert, die hier tabellarisch gegenübergestellt werden, in welchen Untersuchungen sie eher für plausibel erachtet oder nicht bestätigt wurden. Der Eindeutigkeit wegen werden einige englische Fachbegriffe ohne Übersetzung beibehalten, die nachstehend dargelegt werden:
Die Tripod- oder Jacket-(Gründungs-)Strukturen der Windenergieanlagen (WEA) bestehen aus hartem Substrat, die gemäß vieler Studien in vorwiegend sandiges Substrat des marinen Bioms (wie häufig z.B. in der Nordsee) eingebracht werden. An den Oberflächen können sich sessile Organismen wie Muscheln leicht ansiedeln (Biofouling, z.B. Beiersdorf 2014, Kerckhof et al. 2010, Wilson & Elliott 2009). Die etablierten Gemeinschaften stellen wichtige Glieder in der Nahrungskette dar und können marine Säuger, Fische sowie andere Lebewesen anlocken (Attraction). Dieser gesamte sekundäre Prozess wird als Artificial Reef-effect (kurz Reef-effect) bezeichnet (z.B. Degraer et al. 2013b, De Mesel et al. 2013, Leonhard et al. 2011, Leonhard & Pedersen 2006, Reubens et al. 2013). Eher unklar ist bislang jedoch, ob der Reef-effect auch zur Zunahme der Häufigkeiten und der Biomasse von Fischen führen, d.h. die Production steigern kann (De Troch et al. 2013, Vanermen et al. 2013c).
Daneben behandeln einige Publikationen den sog. Reserve-effect, der mit dem Ausschluss von Fischerei und wesentlichem Schiffsverkehr (mit Ausnahme von Wartungsarbeiten) einhergeht. Daher wurden Offshore-Windparks früh als quasi marine Schutzgebiete betrachtet, die durch eine lokale Zunahme an marinen Beutetieren beispielsweise für Raubfische gekennzeichnet sein können (Buck et al. 2004).
Im Kontext von anlage- und betriebsbedingten Wirkungen und Seevögeln wird oft der sog. Barrier-effect angeführt, also die räumliche Barrierewirkung durch Anlagen(-reihen) oder Windparks als Auswirkung auf bestimmte Verhaltensweisen. Der Barrier-effect kann etwa zur sog. Avoidance, d.h. zu einem Meidungs- und Ausweichverhalten von Seevögeln führen. Dazu zählen sowohl unterschiedlich weiträumige Auswirkungen außerhalb als auch unterschiedliche Verhaltensweisen innerhalb von Windparks (Abschnitt 3.1.1). Werden anlage- und betriebsbedingte Wirkungen als (temporäre) Störung wahrgenommen und Tiere in ihren Nahrungs-, Rückzugs- und Fortpflanzungsräumen beschnitten bzw. aus diesen verdrängt, wird von sog. Displacement gesprochen (Abschnitt 3.1.2). Demgegenüber spricht man von Attraction, wenn die Tiere nicht mit Meidungsverhalten auf anlage- und betriebsbedingte Wirkungen reagieren, sie also vielmehr von den bautechnischen Strukturen auf offener See und neuen potenziellen Nahrungsquellen infolge des Reef- und Reserve-effects angelockt werden (Abschnitt 3.1.3). Dies kann mithin eine sog. Habituation bedingen, d.h. das gezielte und regelmäßige Aufsuchen der baulichen Strukturen und Nahrungsgründe innerhalb von Windparks als Folge von Gewöhnungseffekten (Abschnitt 3.1.3).
3 Ergebnisse
Die wesentlichen Wirkungshypothesen von Offshore-Windenergie auf Seevögel, Fische und Benthos, die im Zuge der qualitativen Literaturauswertung identifiziert wurden, stellen zunächst übergeordnete Phänomene dar; sie variieren im Einzelnen dennoch art-, standort- und jahreszeitenabhängig.
3.1 Auswirkungen auf Seevögel
Seevögel werden erst nach einigen Jahren fortpflanzungsfähig und weisen eine geringe Gelegestärke, d.h. Eieranzahl pro Gelege, auf. Entsprechend können geringfügige Veränderungen in der Überlebensrate von adulten Tieren einen erheblichen Effekt auf Populationsebene haben (Stienen et al. 2007). Häufig werden Kollision, Habitatveränderung, Habitatverlust und Barrierewirkung als wesentliche Auswirkungen auf Seevögel gesehen (z.B. Vanermen et al. 2013a, b). Diese können wie folgt gegliedert werden:
(1) direkte Mortalität durch Kollisionen mit WEA bzw. Anlagenteilen (z.B. Rotorblättern), wobei das Kollisionsrisiko von
(a) der Morphologie und den Flugeigenschaften oder
(b) anlage- und betriebsbedingten Wirkungen (Barrierewirkung/Barrier-effect) bzw. Meidungs- und Ausweichverhalten (Avoidance)
abhängig sein kann,
(2) Habitatverlust durch (temporäre) Störung und Vertreibung aus Nahrungsgebieten (Displacement) sowie
(3) indirekte Habitatveränderung und resultierende Verhaltensänderung durch
(a) die Attraktivitätssteigerung einzelner Anlagenstandorte oder Windparks (Attraction) und/oder
(b) Gewöhnungseffekte (Habituation).
Antworten zur konkreten Ursache (warum?) sowie der näheren Art und Weise (wie?) von beobachteten Auswirkungen und spezifischen Verhaltensweisen von (See-)Vögeln zu finden, näher verstehen und beurteilen zu können (May 2015), lässt der aktuelle Forschungsstand dabei nur ansatzweise zu.
3.1.1 Kollisionsrisiko und Mortalität
Der direkten Kollision und Mortalität von (See-)Vögeln wird ein hoher Grad an Aufmerksamkeit zuteil, nicht zuletzt auf Grund der artenschutzrechtlichen Belange (Cook et al. 2012; Vanermen et al. 2012, 2013b). In den letzten Jahren nahm die weitere Entwicklung und Anwendung von Collision Risk Models (CRM, z.B. nach Band 2012) zu, also die Modellierung des Kollisionsrisikos. Dabei gelten einige Vogelarten in Abhängigkeit von ihrem Flugverhalten oder ihrer Morphologie als besonders kollisionsgefährdet (z.B. Furness et al. 2013). Nachfolgend werden die Faktoren für das Kollisionsrisiko infolge bestimmter Verhaltensweisen unterschieden.
Kollisionsrisiko in Abhängigkeit von der Morphologie und den Flugeigenschaften
Insbesondere die folgenden Faktoren, die sich aus der Morphologie und dem Verhalten der Arten ergeben, nehmen offenbar Einfluss auf die Kollisionsgefahr (Tab. 1). Allerdings beruhen viele dieser Angaben auf Experteneinschätzungen, ein auf dem Meer schwieriger empirischer Nachweis fehlt häufig:
Manövrierfähigkeit (Fox et al. 2006a, b; Furness et al. 2013; Garthe & Hüppop 2004, Silbermöwe; Krijgsveld et al. 2011),
Flughöhe innerhalb eines Windparks (Cook et al. 2012, Corman & Garthe 2014, Furness et al. 2013, Johnston et al. 2014, Krijgsveld et al. 2011, Mendel et al. 2014),
Witterungsverhältnisse (z.B. Krijgsveld et al. 2011, Vanermen et al. 2013b),
Tages- und Jahreszeiten (Hill 2015, Krijgsveld et al. 2011, Richardson 2000, Skov & Heinänen 2015, Vaissière et al. 2014, Vanermen et al. 2013b) sowie
bei schlechten Sichtverhältnissen die Anziehung und Desorientierung durch künstliche Lichtquellen (Drewitt & Langston 2006, Erickson et al. 2001, Hüppop & Hilgerloh 2012, Vaissière et al. 2014).
Nach Stumpf et al. (2011) und Furness et al. (2013) nimmt die bevorzugte Flughöhe, die sich je nach Vogelart deutlich unterscheiden kann, eine Schlüsselrolle im Flugverhalten ein. Auf zentraler Nabenhöhe ist das Vogelschlagrisiko theoretisch am größten. Der entsprechende Risiko- bzw. Rotorbereich variiert je nach Anlagentyp zwischen 20 bis 120 m Höhe; die Nabenhöhe befindet sich bislang häufig auf ca. 70 m oder bis zu 110 m über dem Meeresspiegel (Johnston et al. 2014). Krijgsveld et al. (2011) bestimmten daher vier allgemeine Risikozonen für unterschiedliche Flugmuster bzw. -höhen:
025m: mittleres Risiko,
25 150 m: hohes Risiko,
150250 m: mittleres Risiko,
über 250 m: geringes Risiko.
In den letzten fünf Jahren befassten sich viele Studien mit der Analyse und Bewertung artspezifischer Flughöhen in verschiedenen Windparks bzw. Regionen, hauptsächlich in Europa (z.B. Cook et al. 2012, UK; Corman & Garthe 2014, südliche Nordsee; Johnston et al. 2014, UK, Niederlande, Dänemark; Krijgsveld et al. 2011, Egmond aan Zee, Niederlande; Mendel et al. 2014, alpha ventus, AWZ). Sofern Seevögel tatsächlich in einen Windpark fliegen, können einige Familien oder Arten eine bewusste Anpassung der Flughöhe vornehmen. Großmöwen, allen voran die Heringsmöwe (Vanermen et al. 2013a) fliegen bei Pendelflügen und bei der Nahrungssuche in Offshore-Windparks tendenziell höher, d.h. auf Rotorhöhe (Corman & Garthe 2014; Johnston et al. 2014) und können so grundsätzlich einem hohen Kollisionsrisiko ausgesetzt sein (Furness et al. 2013). Furness et al. (2013) sowie Skov & Heinänen (2015) konnten auch Basstölpel auf Höhe des Rotorbereichs beobachten. Nach Krijgsveld et al. (2011) hingegen fliegen Basstölpel innerhalb von Windparks eher unterhalb des Rotorbereichs, weichen diesem also aus, sofern sie den Windpark nicht großräumig meiden (vgl. nächster Abschnitt).
Grundsätzlich wird eine signifikante Variation der Flughöhe im Zusammenhang mit der (lokalen) Witterung als plausibel erachtet (Krijgsveld et al. 2011, Vanermen et al. 2013b). Darüber hinaus kann die Flughöhe sowohl von Sing- als auch Seevögeln von der Wahl der Nahrungsquellen, der Nahrungssuche sowie von Pendel- und Migrationsflügen beeinflusst werden (Blew et al. 2008, Garthe & Hüppop 2004, Johnston et al. 2014, Krijgsveld et al. 2011, Stumpf et al. 2011, Wright et al. 2012). Verschlechtern sich die Sichtverhältnisse, wie u.a. in der Nacht, bei Nebel oder starkem Gegenwind, fliegen ziehende Vogelarten (Hill et al. 2014, Richardson 2000) und z.T. auch Seevögel vergleichsweise tiefer und können somit ihre Kollisionsgefahr erhöhen (Krijgsveld et al. 2011, Vaissière et al. 2014). Entgegen der Barrierewirkung von baulichen (WEA-)Strukturen am Tag auf z.B. Tölpel, Alken und Entenvögel, werden v.a. ziehende Vögel in der Nacht sowie im Herbst bei zunehmendem Gegenwind und schlechten Sichtverhältnissen von WEA-Strukturen angezogen. Bei solch schlechten Witterungsverhältnissen wurden die höchsten Zahlen von ziehenden Vögeln unterhalb 200 m erfasst (Aumüller et al. 2011, Hill et al. 2014). Insbesondere können unter schlechten Witterungsbedingungen künstliche und intensive Lichtquellen ziehende Sing- und Seevögel anziehen, desorientieren (Drewitt & Langston 2006, Erickson et al. 2001, Hill 2015, Hüppop & Hilgerloh 2012, Vaissière et al. 2014) und folglich das Kollisionsrisiko an WEA beeinflussen (z.B. Aumüller et al. 2011, Massen-Kollision von nächtlich ziehenden Arten wie Drosseln, FINO1; Blew et al. 2013; Erickson et al. 2001; Hill et al. 2014; Schuster et al. 2015).
Bestimmte Familien wie Großmöwen (Mendel et al. 2014 und Vanermen et al. 2015, Heringsmöwe; Vanermen et al. 2015, Dreizehenmöwe) und Alken (Vanermen et al. 2012) werden einigen Einschätzungen zufolge durch das veränderte Nahrungsangebot (Abschnitt 3.2.2 und 3.3.2) in Windparks angelockt und gelten so als grundsätzlich kollisionsgefährdet (Tab. 1). Überdies können Offshore-WEA bzw. -Strukturen von Möwen wie der Zwergmöwe und vereinzelt von Seeschwalben wie Fluss- und Brandseeschwalbe als Trittsteinstruktur oder Ruheplatz genutzt werden (Vanermen et al. 2012; Vanermen et al. 2013a, Thorntonbank). Grundsätzlich können erhöhte Interaktionen von Seevögeln mit errichteten Windparks durch ihre Bindung an Offshore-Habitate bzw. marine Beutetiere eine erhöhte Kollisionsgefahr bedingen, so die pauschale Schlussfolgerung einiger Autorinnen und Autoren (z.B. Furness et al. 2013: Heringsmöwe; Vanermen et al. 2012: Zwerg-, Mantel-, Sturm-, Silber- und Dreizehenmöwe, Fluss- und Brandseeschwalbe). Daneben können auf Grund von Verdrängung oder des Barriereeffekts auch Auswirkungen auf den Bruterfolg verbunden sein (Thaxter et al. 2015, Heringsmöwe).
Ferner haben einige Studien art- und standortspezifische Mortalitätsraten pro WEA pro Jahr modellhaft abzubilden versucht. Die Anzahl der Schlagopfer variiert dabei, teilweise in Gradienten je nach Entfernung zur Küste (Brabant et al. 2015, Poot et al. 2011). Als stark kollisionsgefährdete Seevogelfamilien werden in diesem Kontext (Groß-)Möwen, Seeschwalben und Entenvögel angesehen (geschätzte Kollisionsopferzahlen pro Jahr und WEA z.B. nach Brabant et al. 2015, belgische Nordsee: küstenfern 1,8 Möwen; Everaert et al. 2002, Flanders, Belgien: 4-23; Everaert & Stienen 2007, Zeebrugge, Belgien: 27-34, v.a. Seeschwalben; Poot et al. 2011, Egmond aan Zee: küstennah 6,8 Möwen; Vanermen et al. 2013b, belgische Nordsee: 2,4 Möwen).
Barrierewirkung bzw. Meidungs- und Ausweichverhalten (Avoidance)
Zur Beurteilung der (barrierebedingten) Auswirkungen von Offshore-Windparks wird der Anteil der Seevögel, die tatsächlich in einen Windpark fliegen, zumeist über reine Radarbeobachtungen bestimmt (z.B. Desholm 2006, Krijgsveld et al. 2011, Pettersson 2005, Petersen et al. 2006), teils in Kombination mit installierten Kameras und zusätzlichen visuellen Erfassungen. Aber auch eine visuelle Objektverfolgung (visual tracking) per (Schnell-)Boot (Perrow et al. 2015, Abb. 2) findet dafür Anwendung.
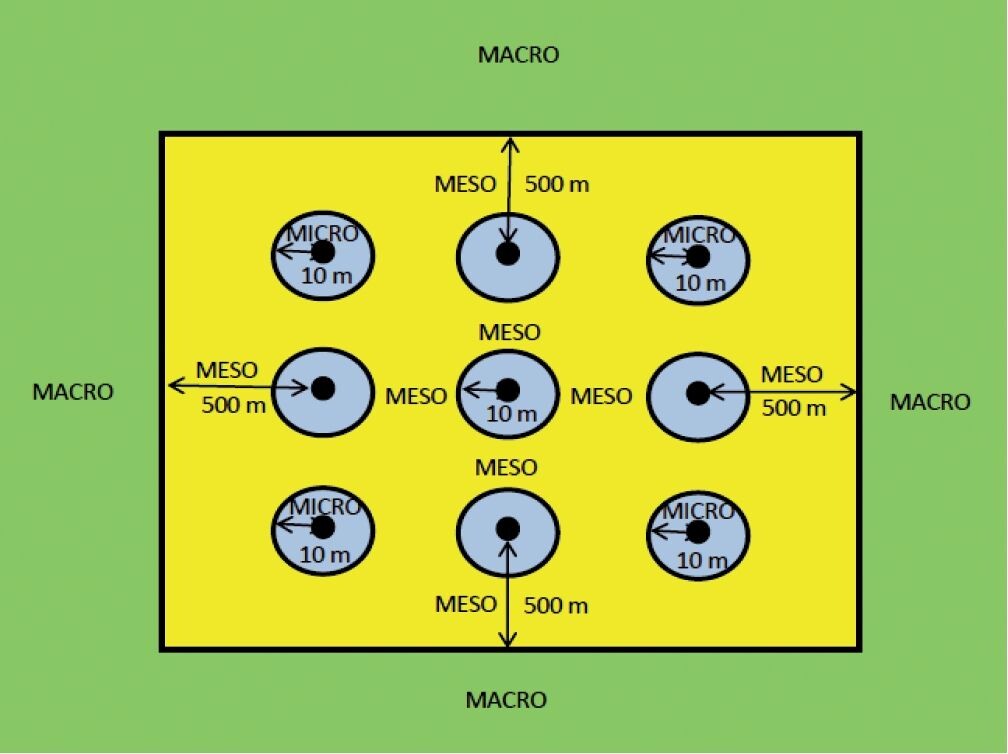
Darüber hinaus werden Kollisionsrisiken modelliert (Offshore-CRM nach Band 2012), um Aufschluss über die spezifische Vermeidungsrate (total avoidance) von im Flug befindlichen Seevögeln zu erhalten (Cook et al. 2014). Das Meidungs- und Ausweichverhalten vor dem Einflug bzw. im Falle des Einflugs in einen Windpark kann nach unterschiedlichen räumlichen Skalen differenziert werden (Band 2012, Blew et al. 2008, Cook et al. 2014, Desholm & Kahlert 2005, Furness et al. 2013, May 2015, Perrow et al. 2015):
(a) Macro-avoidance, d.h. die Flugroute wird außerhalb des Windparks auf Grund seiner Barrierewirkung angepasst, der Windpark wird weiträumig umflogen oder gemieden;
(b) Meso-avoidance, d.h. innerhalb des Windparks findet eine andersartige Verteilung der Vögel bzw. horizontale oder vertikale Anpassung der Flugrichtung infolge der WEA statt; Pendelflüge, Nahrungssuche, Rast und ggf. das Nisten im Windpark erfolgen gezielt zwischen den Anlagenreihen unterhalb der Reichweite der Rotorblätter;
(c) Micro-avoidance, d.h. im Rotorbereich bzw. dem weiter gefassten Risikozylinder
V= Rotorlänge2 Rotorbereich Durchmesser
einer WEA wird quasi in letzter Sekunde horizontal oder vertikal ausgewichen, etwa in Abhängigkeit von der Ausrichtung der WEA zur Flugrichtung und zum Flugwinkel des Vogels.
Die Zuordnung von konkreten Distanzen zu den drei Wirkungsbereichen ist nicht einfach, Cook et al. (2014) definierten sie wie in Abb. 3 dargestellt. Je kleiner die Entfernung zu einzelnen WEA bzw. zum Rotorbereich wird, desto komplizierter werden die Analyse und Bewertung von Verhaltensänderungen (Vanermen et al. 2013a) und entsprechend unsicherer die Erkenntnisse zu Kollisionsursachen.
Zahlreiche Studien stützen sich auf das Macro-avoidance-Verhalten (bzw. Macro-response bei Cook et al. 2014) von überwiegend großen Seevögeln auf Grund der räumlichen Barrierewirkung von Offshore-WEA, z.B. nachgewiesen bei Tölpeln und Alken (Krijgsveld et al. 2011, Leopold et al. 2011, Vanermen et al. 2013a), Entenvögeln sowie Seetauchern (Lindeboom et al. 2011, Tab. 2). Befinden sich diese oder ähnliche Familien etwa auf Nahrungs-, Pendel- oder Kurzstreckenflügen, verändern sie ihre Flugrouten großräumig, um einem Windpark direkt auszuweichen, teilweise mit artspezifischen Distanzen von 0,5 3 km (z.B. Krijgsveld et al. 2011, Basstölpel: 1 2 km; Lindeboom et al. 2011, Tölpel: 0,5km, Meerenten, Alken, Lummen und Seetaucher: > 2 4 km; Petersson 2005, Eiderente: 1 2 km; Vanermen et al. 2015, Trottellumme: 0,5 3 km).
Nach Krijgsveld et al. (2011, Egmond aan Zee) meiden 64 % , nach Skov & Heinänen (2015, Horns Rev 1+2, Rødsand II zwischen 2010-2012) 84 % der erfassten Individuen des Basstölpels das Umfeld der jeweils untersuchten Windparks. Leopold et al. (2013) fanden zudem vergleichend an niederländischen Windparks (Egmond aan Zee und Princess Amalia), dass Basstölpel und Trottellumme den Windpark Princess Amalia v.a. auf Grund einer höheren WEA-Anlagendichte mieden. Für Möwen wiesen Petersen et al. (2006) eine hohe Vermeidungsrate (76 % ) nach, wohingegen Krijgsveld et al. (2011) eine geringe Rate (18 % ) in Distanzen von 12km feststellten. Perrow et al. (2015, Sheringham Shoal) untersuchten im Zuge einer fünfjährigen Studie vor der britischen Küste das Avoidance-Verhalten der Brandseeschwalbe vor und während der Windparkerrichtung sowie während des Betriebs. Dabei wurde deutlich, dass vor Inbetriebnahme des Windparks die überwiegende Mehrheit der anfliegenden Tiere in den Windpark einflog. Nach Inbetriebnahme war dies nur noch bei einem sehr geringen Prozentanteil nachweisbar sie zeigten also aktives Macro-avoidance-Verhalten.
Meso-Avoidance-Verhalten, d.h. die direkte Anpassung des Flugverhaltens und Konzentration des Flugwegs zwischen einzelnen Anlagenreihen, bestätigten diverse Autorinnen und Autoren u.a. für die Eiderente (Christensen et al. 2004, Desholm 2006, Desholm & Kahlert 2005, Kahlert et al. 2004; entgegen Petersen & Fox 2013 und Petersson 2005). Aufgrund ihres Ausweichvermögens wird die Eiderente auf Populationsebene als wenig empfindlich gegenüber Offshore-Windparks eingestuft (Desholm 2006, Desholm & Kahlert 2005, Masden et al. 2009). Auch für die Brandseeschwalbe gibt es Belege von aktiver Meso-Avoidance (Leopold et al. 2011, Perrow et al. 2015). Perrow et al. (2015) wiesen das Verhalten vor der Inbetriebnahme des Windparks für etwa die Hälfte der Vögel in 50 m Distanz zu den WEA nach, wobei sich die durchschnittliche Flughöhe der Tiere nach unten verlagerte und die Wahrscheinlichkeit von kollisionsgefährdeten Vögeln somit um mehr als die Hälfte sank.
Zum Micro-avoidance-Verhalten und der damit einhergehenden sog. Last-second oder auch Last-minute avoidance finden sich insgesamt nur sehr wenige Beobachtungen von Individuen, die die Rotorblätter so dicht passieren, dass eine derartige Ausweich-Aktion überhaupt erforderlich wird. Vereinzelte Nachweise gelangen ansatzweise bisher z.B. über:
visuelle Beobachtungen von Interaktionen mit WEA (Wild Frontier Ecology 2013: acht Individuen, davon z.B. zwei Heringsmöwen mit erfasster Last minute avoidance in 5m Entfernung zu den Rotorblättern),
visuelle und radartechnische Beobachtungen (Krijgsveld et al. 2011: 36 Individuen innerhalb von 50 m um WEA und im Rotorbereich, davon 28 Feldlerchen und nur zwei bis drei Individuen mit erfasster Ausweichbewegung) oder
Aufzeichnungen durch Kameras (Desholm 2005, Petersen et al. 2006: 14 Aufzeichnungen von Vögeln, jedoch nicht in unmittelbarer Nähe, d.h. unter 20 m, zu den WEA).
Desholm (2005) und Krijgsveld et al. (2011) führten dabei die Reaktionen in letzter Sekunde auf schlechte Witterungs- bzw. Sichtbedingungen zurück. Solche Micro-Avoidance Reaktionen erfolgen jedoch selten, da die meisten Vermeidungs- und Ausweichbewegungen bereits vorab auf der Meso- oder Makro-Skala ablaufen (Cook et al. 2014). Die Wahrscheinlichkeit einer Ausweichreaktion in letzter Sekunde für im unmittelbaren Rotorbereich gefährdete Seevögel wird von einigen Autorinnen und Autoren mit mindestens 95 % als hoch eingestuft (Chamberlain et al. 2006, Everaert 2014, Krijgsveld et al. 2011). Die jeweils spezifisch berechneten Raten fließen auch in Kollisionsrisikomodelle ein (Masden & Cook 2016). Cook et al. (2014) ermittelten so über eine Kombination von Schätzungen der Macro-avoidance- und Micro-avoidance-Raten artspezifische Vermeidungsraten (total avoidance rates in Anlehnung an Band 2012). Im Ergebnis lagen die gesamten Vermeidungsraten für die fünf auf Grund ihres Verhaltens und Vorkommens als prioritär untersuchungsrelevant erachteten Arten Silber-, Herings-, Mantel-, Dreizehenmöwe und Basstölpel bei rund 99 % (Cook et al. 2014).
Daher stellt die Kollision an bestimmten Standorten mit regelmäßig durch Windparkareale fliegenden oder ziehenden (See-)Vögeln ein Risiko dar (Cook et al. 2014). Nach wie vor handelt es sich dabei um eine Reihe von Annahmen und Schätzungen, da sämtliche Modellierungen des Kollisionsrisikos bislang auf geschätzten Parametern beruhen, die empirisch sehr schwer zu ermitteln sind. Bereits kleine Unterschiede im Ausweichverhalten können zu deutlich unterschiedlichen Kollisionsrisiken führen. Grundsätzlich ist die Ermittlung von artspezifischen Vermeidungsraten in den räumlichen Skalen sehr komplex und bedarf umfassender Datenerhebungen über mehrere Jahre hinweg (Perrow 07.12.2016, schriftl.).
3.1.2 Habitatverlust durch Störung und Verdrängung aus Nahrungsgebieten
Zu baubedingten Wirkungen auf Seevögel finden sich wenige Erkenntnisse. Indirekte Auswirkungen bestehen beispielsweise im Zuge von Rammarbeiten (Pile-driving) und/oder anderen baubedingten Störungen, die zu artspezifischem Meidungsverhalten von Fischen und rückläufigen lokalen Abundanzen führen können (z.B. Drewitt & Langston 2006, Thomsen et al. 2006). Eine solche Reduktion des Nahrungsangebots kann sich in der Konsequenz auf fischfressende Seevögel wie Seeschwalben und deren Nahrungssuche auswirken (Perrow et al. 2011).
Für Offshore-Windparks wurde früh eine störende Wirkung auf (See-) Vögel angenommen und vermutet, dass sie aus ihren Habitaten verdrängt werden können (z.B. Drewitt & Langston 2006, Langston & Pullan 2003, Stewart et al. 2005; mit Belegen durch Petersen et al. 2006). Meiden (See-) Vögel großräumig und längerfristig sowohl die Windparks als auch das Umfeld, wird nicht nur von Störung und Verdrängung (Displacement), sondern auch von Habitatverlust oder bei eingeschränktem Meidungsverhalten von Habitatverschlechterung gesprochen (Leopold et al. 2013). Oft werden vergleichende Vorher-Nachher-(BACI-) Studien (z.B. Masden et al. 2010a) durchgeführt, um anlage- und betriebsbedingte Wirkungen ermitteln zu können. Einige Studien zeigen anlagebedingte Störungs- oder Verdrängungseffekte bzw. rückläufige Abundanzen für bestimmte Familien oder Arten und Habitatverluste auf. So nahmen Stern- und Prachttaucher (Petersen et al. 2006) sowie Eisente (Petersen et al. 2011) nach Beendigung der Bauphase der dänischen Offshore-Windparks Horns Rev und Nysted signifikant in ihrer Häufigkeit ab. Darüber hinaus werden rückläufige Abundanzen für Trottellumme und Basstölpel (Leopold et al. 2013; Mendel et al. 2014; Petersen et al. 2006, 2011; Vanermen et al. 2012) sowie für Heringsmöwe und Dreizehenmöwe (Leopold et al. 2013, Mendel et al. 2014) beschrieben (Tab. 3). Die Konfiguration eines Windparks kann sich dabei auf die Art und das Ausmaß einer Verdrängung auswirken (z.B. Vanermen et al. 2013a).
Infolge anlagebedingter Barrierewirkungen von Windparks kann es zu einem erhöhten Zeit- und Energieaufwand für Seevögel infolge des großräumigen Meidungsverhaltens kommen (z.B. Fox et al. 2006, Hill 2015, Masden et al. 2010b, Poot et al. 2011, Wilson et al. 2015). Hervorzuheben sind auch hier Stern- und Prachttaucher, die im Falle des Verlustes von wichtigen Ruhe- und Nahrungsgebieten lange Umwege bewältigen müssen. Folglich könnten sie in ihrer Kondition, Fortpflanzungsrate und Populationsgröße beeinträchtigt werden (Mendel & Garthe 2010). Wilson et al. (2015) konnten art-, kolonie- und windparkspezifische Unterschiede auch für weniger untersuchte Arten wie Papageitaucher aufzeigen.
Betriebsbedingte Wirkungen wie z.B. die Rotorbewegung der WEA (z.B. Krijgsveld et al. 2011, Vanermen et al. 2015) können in Abhängigkeit vom Standort (z.B. Braasch et al. 2015) eine andersartige räumliche Verteilung und/oder die Verdrängung bestimmter (fliegender und/oder schwimmender) Seevogelarten aus ihren (Nahrungs-) Habitaten zur Folge haben (z.B. Braasch et al. 2015, Bradbury et al. 2014; Tab. 3). Hill (2015) bestätigte über visuelle Beobachtung das großräumige Meidungsverhalten des Windparks nahe FINO1 für die dort vorkommenden Vogelarten am Tage. So besteht ein Verlust von Ruhe- und Nahrungsgebieten beispielsweise für Basstölpel und Alken (Vanermen et al. 2015). Seetaucher und Entenvögel (Furness et al. 2013, Garthe & Hüppop 2004, Mendel & Garthe 2010) gelten als am stärksten durch Störung und/oder Verdrängung aus Habitaten gefährdet, darunter v.a. Sterntaucher, Eis- und Trauerente (Braasch et al. 2015, Dierschke & Garthe 2006, Leopold et al. 2011, Petersen et al. 2004, 2006; Petersen et al. 2011, Schwemmer et al. 2011). Auch Alken wie die Trottellumme (Mendel et al. 2014, Petersen et al. 2004, Vanermen et al. 2013b) und der Tordalk (Petersen et al. 2004, Vanermen et al. 2013b) zählen hierzu. Für Sterntaucher wiesen Rexstad & Buckland (2012) allerdings nur teilweises Meidungsverhalten im Zuge einer Datenerhebung per Schiff zwischen 2001-2010 vor der englischen Küste nach (Kentish Flats). In Einzelfällen können sich betriebsbedingte Wirkungen auch über große Distanzen bemerkbar machen etwa für Trauerenten zwischen 0,5 bis 6km Entfernung (Petersen et al. 2006).
Zusammenfassend können Störung(en) und/oder die Verdrängung aus Habitaten zu artspezifisch hoher Vulnerabilität und Abnahme der lokalen Population führen (Brabant et al. 2015: Herings- und Mantelmöwe; Drewitt & Langston 2006; Everaert & Stienen 2007: Seeschwalben; Fox et al. 2006a; Furness et al. 2013: Basstölpel, Möwen, v.a. Herings- und Raubmöwe; Garthe & Hüppop 2004: Stern- und Prachttaucher, Eistaucher und Trauerente; Hötker et al. 2006; Poot et al. 2011). Im Zuge einer kumulativen Abschätzung für die südliche Nordsee gehen Platteuw et al. (2015) mit einem DPSIR (Drivers, Pressures, Status, Impact und Response) Modellansatz jedoch von keinem signifikanten Habitatverlust für Seevögel bis 2023 aus.
3.1.3 Indirekte Habitat- und resultierende Verhaltensänderung
Einerseits werden bestimmte Arten durch die Anlagenstrukturen angezogen und nutzen sie als Trittstein oder Ruheplatz auf dem offenen Meer (z.B. Krijgsveld et al. 2011; Attraction nach Vanermen et al. 2013c; Tab. 4). Insbesondere Möwen (Lindeboom et al. 2011, v.a. im Frühjahr und Herbst beim Zug) und Kormorane treten so mit zunehmenden Abundanzen in oder im nahen Umfeld von Offshore-Windparks auf (Leopold et al. 2011, 2013; Nelson et al. 2015), z.T. auch vermehrt während der Brutperiode (Krijgsveld et al. 2011). Vanermen et al. (2013c) beobachteten bereits Brutversuche durch Kormorane und Dreizehenmöwen auf Offshore-Plattformen in der Nordsee. Trotz hoher Individuenzahlen, regelmäßiger Pendelflüge und Nahrungssuche innerhalb und außerhalb von Offshore-Windparks wurde z.B. für den Kormoran offenbar noch kein Nachweis für ein kurzfristig erhöhtes Kollisionsrisiko erbracht (z.B. Lindeboom et al. 2011).
Andererseits reagieren einige Seevogelarten auf das höhere bzw. anders verteilte Nahrungsangebot (Drewitt & Langston 2006, Vaissière et al. 2014; Tab. 4) unter Wasser. Pelagische Fischarten wie Makrele, Bastardmakrele, Atlantischer Hering und Europäische Sprotte (Krägefsky 2014, Abschnitt 3.2.2) nehmen dabei die wichtigste Rolle als Beute ein (Vanermen et al. 2013c). Möwen und Seeschwalben, die bei der Nahrungssuche eine hohe räumliche und zeitliche Variabilität in ihrem Vorkommen aufweisen (Markones et al. 2008), sowie Kormorane und nicht etwa pelagische Seevögel nutzen das erhöhte Nahrungsangebot regelmäßig (Lindeboom et al. 2011). Nachgewiesen wurde dies z.B. für die Dreizehenmöwe (Vanermen et al. 2013c, Bligh Bank) und Heringsmöwe (Vanermen et al. 2015, Thorntonbank). Es wird angenommen, dass diese Familien durch Windparks auf offener See angezogen werden und so (entgegen anderer Einschätzungen, vgl. Abschnitt 3.1.1) in ihrer Häufigkeit zunehmen (Dierschke & Garthe 2006; Vanermen et al. 2012, auch Alken). Dabei spielen einige Umfeld-Faktoren eine Rolle, wie die Häufigkeit von Schleppnetzfischerei, hydrographische Parameter und Gezeiten, Windgeschwindigkeit und Tageszeit, d.h. die so resultierende Nahrungsverfügbarkeit (Markones et al. 2008). Raumzeitliche Variationen in den Interaktionen zwischen Brutgebieten und Offshore-Windparks wurden beispielsweise für die Heringsmöwe im jahreszeitlichen Verlauf (erhöhte saisonale Offshore-Habitatnutzung während der frühen Jungenaufzucht), über mehrere Jahre sowie zwischen Individuen und Geschlechtern durch Thaxter et al. (2015) per GPS-Telemetrie nachgewiesen. Dabei scheint eine Abhängigkeit in den saisonalen Nutzungsmustern von Offshore-Habitaten (männliche Heringsmöwen nutzen diese während der Jungenaufzucht in größerem räumlichen und zeitlichen Maße als weibliche), Ernährungsumstellungen und Veränderungen der räumlichen Verteilung von z.B. marinen Beutetieren, etwa durch die Fischerei denkbar (Thaxter et al. 2015).
Im Zuge der stetigen Expansion von Offshore-Windparks stellt sich die Frage nach Habituation (Gewöhnungseffekten). Aktive Nahrungssuche, Pendelflüge oder die Nutzung von Ruheplätzen (z.B. Markierungsbojen, Abb. 4) werden für einige Seevogelfamilien als attraktivitätssteigernd im Zuge der Errichtung von Windparks angesehen (Tab. 4) jedoch oft nicht empirisch nachgewiesen. Dazu zählen Möwen, Seeschwalben und Entenvögel (Dierschke & Garthe 2006; Leonhard et al. 2013, Trauerente; Thaxter et al. 2015, Heringsmöwe), Kormorane (Dierschke & Garthe 2006, Krijgsveld et al. 2011) und Alken (Vanermen et al. 2012). Hingegen konnten für die relevanten Taucherarten selbst fünf bis sechs Jahre nach Abschluss der Bauphase keine Gewöhnungseffekte, aber noch immer aktives Macro-avoidance-Verhalten nachgewiesen werden (Petersen et al. 2008, Petersen & Fox 2007).
3.2 Auswirkungen auf Fische
Auch für Auswirkungen von Offshore-Windparks auf Fische wurden bau-, anlage- und betriebsbedingte Effekte untersucht.
3.2.1 Baubedingte Wirkungen
Störungen während der Bauphase können Fische und ihre Lebensstadien wie Eier oder Larven offenbar über Dutzende von Kilometern beeinflussen (Thomsen et al. 2006). Baubedingte Unterwassergeräusche und Vibrationen können zu Störungen und Habitatverlust bzw. artspezifischem Meidungsverhalten und rückläufigen lokalen Abundanzen oder Populationen führen (z.B. Bailey et al. 2014, Drewitt & Langston 2006, Thomsen et al. 2006). Der Großteil der ausgewerteten Publikationen konzentriert sich auf Auswirkungen durch Rammarbeiten (Abb. 5), die Installation von Schwerkraftfundamenten und/oder die Kabelverlegung. Rammarbeiten und andere impulsive Geräuschquellen können Fische stören oder vertreiben (z.B. Haelters et al. 2013, Vaissière et al. 2014). Fische und ihre Lebensstadien können temporäre oder permanente Hörschäden erleiden sowie je nach Entfernung und Lärmintensität schwerwiegend verletzt oder getötet werden (z.B. Halvorsen et al. 2012, Krägefsky 2014).
Bau- und betriebsbedingter Lärm beläuft sich zum Großteil auf Frequenzen von unter 1kHz und liegt somit deutlich innerhalb der Hörweite von Fischen (Haelters et al. 2013, z.B. Kabeljau). Die Lärmexposition variiert dabei mit der Windgeschwindigkeit und wird von der Schallausbreitung der jeweiligen WEA zum umgebenden Wasser beeinflusst (Hammar et al. 2014). Auch die Größe, das Lebensstadium und artspezifische Gehör eines Fisches spielen für die Empfindlichkeit eine Rolle (z.B. Haelters et al. 2013). Dabei gilt es als schwierig zu beurteilen, wie sich geringe Geräuschpegel und deren Exposition auf das natürliche Verhalten von Fischen auswirken oder ihre Kommunikation und Orientierungssignale beeinträchtigen (z.B. Müller-Blenkle et al. 2010, Thomsen et al. 2006, Wahlberg & Westerberg 2005).
Für den Kabeljau (im Kattegat) besteht während der Laichzeit ein hohes Risiko durch Rammarbeiten, da diese eine hemmende Wirkung auf den Laichvorgang bis in max. 50km Entfernung entfalten können (Müller-Blenkle et al. 2010, Popper & Hastings 2009, Thomsen et al. 2012). Nach Krägefsky (2014) können Lärmemissionen durch Rammarbeiten von Hering und Makrele bis in mehr als 80km Entfernung wahrgenommen werden. Darüber hinaus gelten Seezunge (Bolle et al. 2012, v.a. Larven; Thomsen et al. 2012) und Europäischer Wolfsbarsch (Debusschere et al. 2015, v.a. Larven oder Juvenile) als vornehmlich durch Rammarbeiten beeinträchtigbar. Nach Elliott (2002) sind insbesondere Sandaale von Störung und Habitatverlust betroffen.
Für bestimmte Arten scheint klar, dass die temporäre Exposition baubedingten Lärms (vergleichbar mit Rammarbeiten) keine unmittelbare oder mittelbare Mortalität bedingen (Bolle et al. 2012, Larven der Seezunge; Casper et al. 2012 und 2013, u.a. Streifenbarsch; Debusschere et al. 2015, Europäischer Wolfsbarsch). Genaue Schwellenwerte für Lärmemissionen mit Bezug auf die Verletzung, beeinträchtigte Entwicklung oder die Mortalität von Individuen sind unbestimmt (Haelters et al. 2013). Nach Hammar et al. (2014) spielen Auswirkungen infolge der Kabelverlegung und Sedimentumlagerung/-erosion eine eher untergeordnete Rolle, da sie einen geringen Einfluss auf adulte Fische (hier Kabeljau) bzw. ihren Laichvorgang haben.
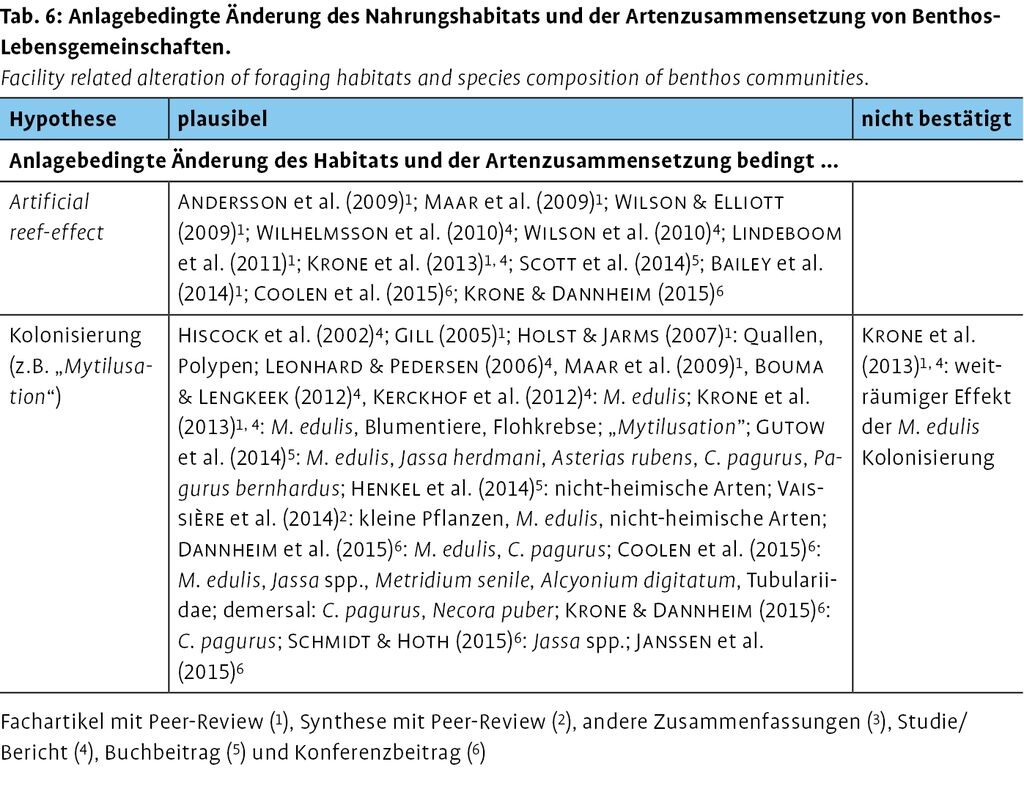
3.2.2 Anlagebedingte Änderung des Habitats und der Artenzusammensetzung (Reef-effect)
Der am häufigsten angeführte und untersuchte Effekt von Offshore-Windparks geht auf den künstlichen Riff-Effekt (Reef-effect) zurück (z.B. Bailey et al. 2014, De Troch et al. 2013, Kerckhof et al. 2010, Schmidt & Hoth 2015, Vaissière et al. 2014). Daneben spielt auch der Reserve-effect eine Rolle, d.h. ein Schutzeffekt des Windparks auf Grund eingeschränkter Fischerei (s. Abschnitt 2). Zusammen begünstigen die genannten Effekte Änderungen in der Artenzusammensetzung (Bailey et al. 2014, Krägefsky 2014, Vaissière et al. 2014), ohne dass jedoch großräumige und langfristige Effekte bislang nachgewiesen sind (Vandendriessche et al. 2013a).
Durch den Reef-effect bzw. das sog. Biofouling (z.B. Beiersdorf 2014, Abschnitt 2; Abb. 6) werden neue Habitate und somit neue Möglichkeiten zur Nahrungssuche für verschiedenste Lebewesen in der marinen Nahrungskette geschaffen (Bailey et al. 2014, Drewitt & Langston 2006, Schmidt & Hoth 2015, Scott et al. 2014, Vaissière et al. 2014). Änderungen in der Zusammensetzung von Arten-/Beute-Gemeinschaften, die mit der Kolonisierung von auf hartes Substrat spezialisierte Arten einhergehen, spiegeln sich u.a. in der lokalen Häufigkeit wider (Bailey et al. 2014; Drewitt & Langston 2006; Krägefsky 2014, demersale und pelagische Arten; Schmidt & Hoth 2015; Vandendriessche et al. 2013b, u.a. mehr demersale, strukturgebundene Fische bis in 180 m Entfernung; Tab. 5). Fische werden durch die neuen Strukturen angezogen, die Rückzugsorte oder Nahrung bereitstellen (Reubens et al. 2013; Vandendriessche et al. 2013b: Kabeljau im 40-m-Radius um WEA). Zum Beispiel fanden Krone et al. (2013, Deutsche Bucht) und Reubens et al. (2011a, Thorntonbank) erhöhte Abundanzen von Amphipoden wie Flohkrebse (Jassa spec.), eine für Franzosendorsche (Trisopterus luscus) leicht zugängliche Beute. Dannheim et al. (2015, alpha ventus) konnten ebenfalls lokal erhöhte Abundanzen des Franzosendorschs sowie der Bastardmakrele bestätigen. Winter et al. (2015, OWEZ) stellten höhere Abundanzen verschiedener Arten, darunter Kabeljau und Franzosendorsch fest, fanden jedoch keinen statistisch signifikanten Effekt im Vergleich zu lokalen Abundanzen in Referenzgebieten ohne Windparks.
Dabei nehmen die Jahreszeiten Einfluss auf die Verteilung bzw. Migration einzelner Fischarten zum Laichen (Reubens et al. 2013, Kabeljau Höchstwerte zwischen Mai bis November, Franzosendorsch Höchstwerte zwischen September bis November; Winter et al. 2015, Sommer). Weiterhin werden Änderungen in der vertikalen Verteilung von Arten beschrieben (Chimienti et al. 2014; v.a. in der näheren Umgebung der Fundamente, Winter et al. 2015; Tab. 5). Krone et al. (2013, Deutsche Bucht) fanden zwischen 5 bis 20 m Wassertiefe vorwiegend die Bastardmakrele, in 28m Tiefe hingegen Kabeljau und Franzosendorsch als dominierende Arten. Degraer et al. (2013a) konnten beim Vergleich eines WEA-Areals mit Schwerkraftfundamenten und zwei sandigen Referenzgebieten (u.a. Gootebank, belgische Nordsee) keine Einschränkungen in der Nahrungsverfügbarkeit für die untersuchten Fischarten feststellen. Vielmehr war der körperliche Status der Tiere geringfügig besser (increased length and enhanced fullness index, S. 179). De Troch et al. (2013, belgische Nordsee) bestätigten ebenfalls die Eignung von Windparks als Nahrungsgründe für Kabeljau und Dorsch, ohne Anzeichen interspezifischer Konkurrenz zu finden.
3.2.3 Betriebsbedingte Wirkungen
Zu betriebsbedingten Wirkungen können vorrangig (artspezifische) Verhaltensänderungen wie die Meidung von Windparkgebieten gezählt werden. Dieser Effekt kann zum einen der Empfindlichkeit gegenüber elektromagnetischen Feldern von Netzkabeln geschuldet sein, da diese die Fortbewegung und Navigation von Fischen (in direkter Umgebung signifikant) beeinträchtigen können (Gill et al. 2009, Krägefsky 2014, Normandeau Associates, Inc. et al. 2011). Als empfindlich gelten Knochenfische und Aale (Westerberg & Lagenfelt 2008), Plattenkiemer (Normandeau Associates, Inc. et al. 2011) und ziehende Arten (Vaissière et al. 2014).
Zum zweiten beeinflussen betriebsbedingte Unterwassergeräusche und Vibrationen durch Generatoren und Getriebe bestimmte Fischarten (nicht so den Kabeljau gemäß Kastelein et al. 2008 und Müller 2007; Krägefsky 2014, Vassière et al. 2014). Diese sind allerdings weitaus niedriger als baubedingte Geräusche und führen nach Krägefsky (2014) zu keiner unmittelbaren Beeinträchtigung. Die Wahrnehmung von Geräuschen oder Lärm hängt hier ebenfalls von der Entfernung zur Quelle, dem Klangbild und artspezifischen Hörvermögen ab so ergeben sich beispielsweise signifikante Variationen zwischen Makrele und Hering (Krägefsky 2014). Zur Wahrnehmung betriebsbedingter Geräusche durch Fische ohne Schwimmblase gilt eine Entfernung von unter 10km, für Fische mit normalem Hörvermögen unter 1km (Krägefsky 2014). In Abhängigkeit von der Schallstärke des Lärms kann die Maskierung von Kommunikationslauten und Orientierungssignalen eine Folge sein (Andersson et al. 2011, Codarin et al. 2009, Krägefsky 2014). Die Empfindlichkeit gegenüber den Schalldruckpegeln und der Teilchenbewegung durch den WEA-Betrieb wird von einigen Publikationen gestützt (Andersson 2011, Engås et al. 1995, Sigray & Andersson 2011, Wahlberg & Westerberg 2005: mit Bezug auf Kabeljau; Müller-Blenkle et al. 2012, Popper & Hastings 2009). Werden Frequenzen von 50Hz überschritten, kann ggf. das Laichen des Kabeljaus beeinträchtigt oder gehemmt werden (Hammar et al. 2014, Slabbekoorn et al. 2010, Wahlberg & Westerberg 2005, Abschnitt 3.2.1). Andere Studien widerlegen jedoch die artspezifische Empfindlichkeit des Kabeljaus (Bergström et al. 2013b, Leonhard et al. 2011, Lindeboom et al. 2011, Reubens et al. 2011b, Winter et al. 2010).
3.3 Auswirkungen auf die Benthos-Lebensgemeinschaft
Neben baubedingten Auswirkungen von Offshore-Windparks spielen Habitatänderungen bzw. eine veränderte Benthos-Zusammensetzung infolge des Reef- und Reserve-effects sowie ihre Wechselwirkungen mit anderen Tiergruppen eine große Rolle.
3.3.1 Baubedingte Wirkungen
Vergleichbar mit den baubedingten Wirkungen auf Fische (Abschnitt 3.2.1), können Störungen von Sedimenten oder Verteilungsmustern der Arten und Individuen durch die Kabelverlegung, die Bewegung von Maschinen sowie die Installation der Fundamente und Netzkabel auftreten. Die baulichen Maßnahmen ziehen Sediment-Verwirbelungen sowie eine entsprechende Wassertrübung nach sich. Baubedingte Effekte auf die Wassertrübung waren in Belgien im Umfeld eines untersuchten Windparks lokal und temporär, aber ohne signifikante Unterschiede im Vorher-Nachher-Vergleich nachweisbar (Van den Eynde et al. 2013). Am Eingriffsort ist zunächst mit einem Verlust an ggf. lokal sesshaften Benthos-Gemeinschaften (Henkel et al. 2014), Wirbellosen (Würmer und Quallen), Seegräsern, Seesternen, Krustentieren u.ä. zu rechnen (Vassière et al. 2014). Sessile und mobile Wirbellose können im Zuge der Auskolkung für die Gründungsstrukturen begraben oder vertrieben werden (Henkel et al. 2014). Die erhöhte Versandung oder Verschlickung und reduzierte Lichtdurchlässigkeit durch Bauarbeiten wirkt vorwiegend beeinträchtigend auf Meeresalgen (Henkel et al. 2014), soweit diese in Offshore-Windparks vorkommen. Lärmemissionen während der Rammarbeiten können nach Einschätzung von Gill (2005) zu einer temporären Verdrängung führen, u.a. von mobilen benthischen Tieren.
3.3.2 Anlagebedingte Änderung des Habitats und der Artenzusammensetzung (Reef-effect)
Viele Publikationen stellen die zunehmende Habitatqualität und -komplexität sowie lokale Artenvielfalt auf Grund des Reef-effects (z.B. De Mesel et al. 2013) in den Mittelpunkt.
Dabei ändern sich die Verhältnisse in der lokalen Nahrungskette durch den Reef-effect (Andersson et al. 2009, Bailey et al. 2014, Coolen et al. 2015, Lindeboom et al. 2011, Maar et al. 2009, Scott et al. 2014, Wilhelmsson et al. 2010, Wilson et al. 2010, Wilson & Elliott 2009; Tab. 6). So steigt beispielsweise für die mobile, demersale Megafauna (d.h. Krustentiere und Fische, z.B. Krone & Dannheim 2015) das Nahrungsangebot, die wiederum pelagische Seevögel anlocken (Abschnitt 3.1.3). Die Fundamente der Anlagen gelten gar als Biomasse-Hotspot (Krone et al. 2013). So wiesen Degraer et al. (2013b) für den Windpark Thorntonbank eine Zunahme der Biomasse um das 14-Fache im Vorher-Nachher-Vergleich nach. Einige Autorinnen und Autoren setzen die gesteigerte Habitatqualität und -komplexität auch in direkte Beziehung mit dem Reserve-effect (Bailey et al. 2014, Maar et al. 2009, Krone et al. 2013, Scott et al. 2014; Abschnitt 2).
Bestätigt wird insbesondere die Kolonisierung durch die Miesmuschel (Mytilus edulis, kurz M. edulis) (Maar et al. 2009, Krone et al. 2013, Gutow et al. 2014, Vaissière et al. 2014, Coolen et al. 2015, Dannheim et al. 2015; Abb. 4 und Tab. 6), mit der Ausbildung dichter Gürtel an Fundamenten (Hiscock et al. 2002, Kerckhof et al. 2012) oder zwischen einzelnen Anlagen (Bouma & Lengkeek 2012). So wiesen Krone et al. (2013) 35-mal mehr Biomasse (Makrozoobenthos) auf Gründungsstrukturen (1024m²) durch die Biofouling-Kolonisierung von M. edulis im Vergleich zu einer gleichgroßen weichen Substratfläche in der AWZ der Deutschen Bucht nach. Sie bezeichnen dieses Phänomen als Mytilusation.
Darüber hinaus werden durch die neu eingeführten harten Substrate auch zunehmende Abundanzen von Quallen (Jellification) in küstennahen Gebieten als wahrscheinlich erachtet (Holst & Jarms 2007). Janssen et al. (2015) rechnen mit einer Zunahme der Ohrenqualle (Aurelia aurita) um 1580 % , gefolgt von einer Kolonisierung durch weit verbreitete Blumentiere (Anthozoa) (Krone et al. 2013, Richardson et al. 2009). Eine weitere, oft im Umfeld der Fundamente nachgewiesene und die demersale Megafauna dominierende Art ist der Taschenkrebs (Cancer pagurus) (Coolen et al. 2015, Dannheim et al. 2015, Gutow et al. 2014, Krone & Dannheim 2015). Auch die um ein Vielfaches kleineren Flohkrebse (Jassa spec.) weisen zunehmende Abundanzen auf (Coolen et al. 2015, Krone et al. 2013; Schmidt & Hoth 2015: teilweise mit Deckungsgraden von 65 100 % ).
Zusammenfassend betrachten Lindeboom et al. (2011) die neu entstehenden, künstlichen Riffe als Trittsteine für die Ausbreitung oder Vergrößerung der Artbestände, die an hartes Substrat gebunden sind. Kritisch sehen Degraer et al. (2013a) und De Mesel et al. (2015) derartige Effekte im Hinblick auf eine möglicherweise begünstigte Expansion nicht heimischer Arten etwa in der Nordsee.
In einigen der genannten Quellen werden Unterschiede in der Kolonisierung bzw. Zusammensetzung von Benthos-Gemeinschaften in Abhängigkeit von bestimmten Faktoren herausgearbeitet:
Zeit nach der Installation von WEA (Dannheim et al. 2015, De Mesel et al. 2013, Leonhard & Pedersen 2006, Kerckhof et al. 2012; Krone & Dannheim 2015: größter Anstieg 2-3 Jahre später),
jahreszeitliche Dynamiken (Dannheim et al. 2015 und Gutow et al. 2014: größte Zunahme der Megafauna im Herbst; De Mesel et al. 2013: höchste Biofouling-Dichte im Frühjahr und Sommer; Krone et al. 2013: Epifauna) und
Wassertiefe bzw. vertikale Zonierung, je nach Lichteinfall und Nahrungsverfügbarkeit (De Mesel et al. 2013: Spritzwasserzone 05 m ü.d.M., intertidale Zone 1 m ü.d.M. bis 3m Tiefe, subtidale Zone 328 m Tiefe; Dannheim et al. 2015, Gutow et al. 2014, Krone et al. 2013, Schmidt & Hoth 2015: Variationen in 1, 5 und 10 m Tiefe; Bouma & Lengkeek 2012, Coolen et al. 2015, Dannheim et al. 2015, Krone et al. 2013: Variationen in 1520 m und 2028 m Tiefe bzw. am Tripod).
4 Bilanz und Ausblick
Die Nutzung der Windenergie auf dem offenen Meer ist noch jung. Dabei sind Meeresgebiete vielfach bereits stark von Aktivitäten durch den Schiffsverkehr und die Fischerei geprägt. Durch den Bau und Betrieb von Offshore-Windparks kann mit zusätzlichen Auswirkungen auf marine Ökosysteme gerechnet werden, die wiederum einer zeitlichen Dynamik unterliegen können (Damian & Merck 2014). Innerhalb des letzten Jahrzehnts wurden die Auswirkungen der Windenergie auf dem Meer verstärkt erforscht. Während einige der Wirkungszusammenhänge bereits recht gut nachvollzogen werden können, wird um andere noch sehr gerungen oder es besteht gänzliche Unklarheit. Vor allem in Europa konnten über die letzten Jahre nennenswerte Erkenntnisgewinne erzielt werden.
Viele der hier dargelegten Hypothesen sind an spezifische Arten oder Familien, Standorte, Windparks oder Regionen sowie Jahreszeiten gebunden. Schließlich existieren bislang nur wenige vergleichende Studien und auch nur wenige, die kumulative Wirkungen einbeziehen (z.B. Brabant et al. 2015, Damian & Merck 2014), nicht zuletzt da auch die meisten Forschungsvorhaben nicht staatenübergreifend realisiert werden konnten. Zudem ist die (empirische) Datenerhebung über dem offenen Meer im Vergleich zum Land um ein Vielfaches erschwert; dies betrifft allein schon das Beobachten von Vogel-Kollisionen und Auffinden von Kollisionsopfern (Hill 2015). Dennoch lassen sich einige Thesen ableiten. So kann bereits besser eingeschätzt werden, welche Tierarten zu den vulnerablen gegenüber Offshore-Windparks zählen u.a. in Abhängigkeit von ihrer Morphologie, der Phänologie, räumlichen oder baulichen Besonderheiten von Offshore-Windparks sowie der Wahrnehmung von Störungen bzw. entsprechenden Reaktionen durch Verhaltensänderungen.
Dabei sind vielfach Ursachen von spezifischen Verhaltensweisen, etwa von Seevögeln, grundsätzlich noch ungeklärt. Es bleibt nach wie vor sehr kompliziert, kausale Effekte zum Kollisionsrisiko, Avoidance-Verhalten oder zur Anlockung zu ermitteln und nachzuvollziehen. Einige Arten oder Familien sind von einem (erhöhten) Kollisionsrisiko betroffen. Tendenziell neigen Seevögel ohne die in Abschnitt 3.1.1 explizit genannten offenbar dazu, eine niedrige Flughöhe unterhalb des Rotorbereichs in Windparks einzunehmen (< 20 m ü.d.M.) (nach Krijgsveld et al. 2011). Darüber hinaus werden Offshore-Windparks durch viele der hier relevanten Arten in hohem Maße gemieden (v.a. während der Zugzeiten nach Skov & Heinänen 2015 und während der Betriebszeiten nach Coppack et al. 2015). Für Heringsmöwe und Eiderente konnte die gezielte Meidung des Rotorbereichs bereits nachgewiesen werden. Derartige Untersuchungsergebnisse bedürfen jedoch weiterer gezielter Forschung (May 2015).
Hingegen sind andere Arten, z.T. auch Fische und Benthos, von (temporären) Störungen, Verdrängungseffekten oder einem Habitatverlust gekennzeichnet (z.B. Furness et al. 2013, Garthe & Hüppop 2004). Der langfristige Habitatverlust für Seetaucher sowie bestimmte Meeresenten und Alken infolge ihrer hohen Störungsempfindlichkeit und ihres großräumigen Meidungsverhaltens (Furness et al. 2013) dienen dafür als Beispiele. Dennoch fehlen auch hier weitergehende Erkenntnisse für artspezifische Reaktionen und langfristige Folgen für vertriebene Individuen bzw. deren Populationen. Busch & Garthe (2016) diskutieren in diesem Kontext einen Ansatz zur Bewertung von Verdrängungseffekten, um Rückschlüsse auf (erhebliche) Beeinträchtigungen auf die lokale Population am Beispiel der Trottellumme zu ermöglichen.
Demgegenüber können die hinzukommenden baulichen Elemente im marinen Biom neue Habitate für verschiedene Tiergruppen schaffen. Verbesserte Nahrungsgründe hätten das Potenzial, die bisher durch Avoidance charakterisierten Arten an Windparks zu gewöhnen und anzulocken (Vanermen et al. 2015). Gleichzeitig kann dies jedoch (Mortalitäts-)Risiken (z.B. Vanermen et al. 2013b) beeinflussen.
Eine Reaktion auf solche keineswegs unüblichen und stets verbleibenden Unsicherheiten stellen Adaptive-Management-Ansätze dar (Köppel et al. 2014, Williams & Brown 2014). Vermeidungsmaßnahmen (Gartman et al. 2016) und, sofern möglich, auch Kompensationsmaßnahmen müssen hier Abhilfe schaffen (Lüdeke et al. 2014). Dies setzt allerdings voraus, dass langfristige, kumulative Wirkungen sowohl auf Populationsebene als auch in Verbindung mit marinen Schutzgebieten berücksichtigt und beurteilt werden können. Gemäß modellhaften Berechnungen zu kumulativen Wirkungen von Poot et al. (2011, 16, OWEZ) seien die nachgewiesenen Auswirkungen multipler Offshore-Windparks in der niederländischen Nordsee noch weit entfernt von Werten, die zu rückläufigen Trends auf Populationsebene für eine Reihe an Seevogel- und ziehenden Singvogelarten führen würden darunter z.B. Eiderente, Trottellumme, Tordalk, Papageitaucher, Zwergseeschwalbe, Star, Feldlerche u.v.m. Derartige Ergebnisse gilt es einzuordnen, im großräumigen Verbund weitergehend zu fokussieren und zu untersuchen.
Insgesamt bleiben zahlreiche Forschungsfragen offen, insbesondere zu Seevögeln. Kollisionsrisikomodelle schaffen in Hinblick auf den zunehmenden Ausbau der Offshore-Windparks keine alleinige Abhilfe, um Kollisionen von (See-)Vögeln auf offener See pro WEA und Jahr hinreichend bestimmen zu können. So sollten beispielsweise das artspezifische Micro-avoidance-Verhalten mit Hilfe von visuellen und radartechnischen Beobachtungen (Abb. 7) besser untersucht sowie artspezifische Populationsmodelle weiterentwickelt werden, um kumulative Risiken bewerten zu können (Brabant et al. 2015).
Sofern sich Attraction-Effekte in den kommenden Jahren bestätigen, können in der Folge des so beeinflussten Kollisionsrisikos neuartige Problemfelder entstehen (Vanermen et al. 2013a). Überdies können bisher vernachlässigbare oder wenig berücksichtigte Arten wie z.B. Papageitaucher und Eissturmvogel von zunehmenden Offshore-Windpark-Planungen in ihren Verbreitungsarealen betroffen sein (Leopold et al. 2013). Gleichzeitig sollten hinreichende Korridore frei von Offshore-Windparks gehalten werden (Damian & Merck 2014). Aktuell können zur möglichen Habituation kaum Aussagen getroffen werden (Mendel et al. 2014). Nicht zuletzt bedarf es eines Windpark-übergreifenden und längerfristigen Monitorings, um auch zeitliche und räumliche Variationen besser abschätzen (Vanermen et al. 2015) und in Umweltprüfungen berücksichtigen zu können (Thaxter et al. 2015).
In Bezug auf Fische und Benthos verbleibt zu validieren, welchen längerfristigen Veränderungen die Biomasse und Artenzusammensetzung an den Anlagen- und Gründungsstrukturen unterliegen (Degraer 2013b) und welche Rolle dabei dem Fischereiausschluss zukommt. Auch ist z.B. das Wissen über Auswirkungen im Falle einer Sedimenterwärmung durch die verlegten Kabel noch nicht hinreichend. Veränderungen in der Nahrungsverfügbarkeit sowie ggf. verhaltensbiologische Anpassungen von Seevögeln gelten es ebenfalls noch näher zu untersuchen und zu verstehen.
Dank
Die Autoren bedanken sich bei Dr. Georg Nehls für die kritische Durchsicht des Manuskripts, bei Dr. Martin Perrow für wertvolle Hinweise zu den Untersuchungen an Brandseeschwalben sowie bei Dr. Aonghais Cook (und Co-Autoren) für die Überlassung von Abb. 3 (mit freundlicher Genehmigung durch Marine Scotland, Dr. Finlay Bennet).
Literatur
siehe beitragsübergreifende Bibliographie in diesem Heft
Fazit für die Praxis
Innerhalb des letzten Jahrzehnts wurden die Auswirkungen der Offshore-Windenergie, v.a. in Europa, zunehmend erforscht.
So lassen sich bereits erste Wirkungshypothesen z.B. zum Kollisionsrisiko bzw. Meidungsverhalten von Seevögeln und zu Habitatveränderungen von Benthos-Lebensgemeinschaften sowie für die Fischfauna beschreiben.
Dennoch mangelt es bis heute an empirischen Daten, es fehlen Meta-Studien (v.a. Auswertungen von Monitoring-Ergebnissen) und längerfristige Effekte sind weitgehend unbekannt.
So besteht noch erheblicher Erkenntnisbedarf etwa bezüglich Verhaltensänderungen und Mortalität von lokalen und regionalen Populationen von Seevögeln.
Bislang gibt es nur wenige vergleichende Studien, die mehrere Offshore-Windparks oder gar kumulative Wirkungen auf Arten(gruppen) untersuchen und bewerten, um etwa auch ggf. erforderliche Korridore von Windenergie frei zu halten.
-
Abb. 1: Vogelzählung auf See.
Bird counting at sea.Graeme Pegram -
Abb. 2: Festrumpfschlauchboot zur Verfolgung von Brandseeschwalben-Flugbewegungen an einem britischen Offshore Windpark.
Rigid inflatable dinghy to trace the flight movements of Sandwich terns at a British offshore wind farm.© Seafari Marine Services Ltd. -
Abb. 3: Schema räumlicher Skalen für Micro-, Meso- und Macro-avoidance/-response; Punkte kennzeichnen die Lokalisierung der einzelnen WEA (unmaßstäblich).
Scheme of spatial scales for micro-, meso- and macro-avoidance/-response; points indicate the locations of the individual wind power plants (without scale). Cook et al. (2014) -
Abb. 4: Brandseeschwalben (Thalasseus sandvicensis) auf einer Markierungsboje zur Errichtung des Sheringham Shoal Offshore Windparks (Perrow et al. 2015).
Sandwich terns (Thalasseus sandvicensis) on a marking buoy established for the Sheringham Shoal Offshore Wind Farm (Perrow et al. 2015). Martin Perrow -
Abb. 5: Rammarbeiten beim Offshore-Windpark Butendiek.
Pile driving for the offshore wind farm Butendiek.Andreas Buchheim -
Abb. 6: Der Biofouling-Effekt an Pfahlgründungen von Offshore WEA, hier: Miesmuschel-Besiedlung (Mytilus edulis) im Windpark alpha ventus.
The biofouling effect on pillar foundations of offshore wind farms, here: settlement of Blue mussels (Mytilus edulis) in the wind farm alpha ventus.Philipp Kastinger -
Abb. 7: Vogelradarerfassung auf See.
Radar detection of birds at sea.Christian Vlasak -
Auswirkungen der Offshore-Windenergie auf Seevögel, Fische und Benthos -
Auswirkungen der Offshore-Windenergie auf Seevögel, Fische und Benthos -
Auswirkungen der Offshore-Windenergie auf Seevögel, Fische und Benthos -
Auswirkungen der Offshore-Windenergie auf Seevögel, Fische und Benthos -
Auswirkungen der Offshore-Windenergie auf Seevögel, Fische und Benthos -
Auswirkungen der Offshore-Windenergie auf Seevögel, Fische und Benthos







Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.