Die Bachmuschel (Unio crassus) überlebt noch in Metapopulationen
Abstracts
Die Verbreitung der Bachmuschel ist natürlicherweise diskontinuierlich, und zwar sowohl auf lokaler Ebene innerhalb einer Gewässerstrecke oder im Verlauf eines Gewässers als auch im regionalen Maßstab in einem Gewässereinzugsgebiet oder einer Landschaft. Diese Situation wurde im Laufe der Zeit durch anthropogene Einflüsse stark verschärft, was zu einer zunehmenden Isolation der Restbestände, stark rückläufigen Individuenzahlen und häufig sogar zum regionalen Aussterben der Art führte.
Begrenzte Ressourcen erfordern unter diesen Umständen sachgerechte und zielführende Ansätze zu ihrem Schutz. Zwei Konzepte stehen dabei im Vordergrund: die kleinste überlebensfähige Population und die Metapopulation. In Gebieten, in denen die Bachmuschel noch eine gewisse Verbreitung besitzt, ist das Metapopulationskonzept mit Stamm- oder Quellpopulationen, Nebenpopulationen und Latenzpopulationen geeignet, um die Situation und die Perspektiven zu analysieren. Das Verständnis funktionaler Zusammenhänge ist die Voraussetzung für sinnvollen und nachhaltigen Mitteleinsatz im Artenschutz. Zwei Beispiele aus Baden-Württemberg (Bachsysteme in der Oberrheinniederung, Neckarzufluss Eschach) zeigen die konkrete Anwendung.
The Thick Shelled River Mussel (Unio crassus) still survives in metapopulations. Protection concepts for an endangered species and prioritisation of measures
The distribution of the thick shelled river mussel is naturally discontinuous, both locally, within the course of a stream/river as well as on a regional scale in the catchment area of a river or in a landscape. The situation has been significantly exacerbated by anthropogenic influences, causing an increasing isolation of remnant populations, strongly declining numbers of individuals, and frequently even the regional extinction of the species. Since the resources for the protection of the species are limited appropriate and target-oriented approaches are required. In terms of protection measures two concepts are to the fore: the minimum viable population and the metapopulation.
In areas where the thick shelled river mussel still has a certain distribution the concept of metapopulations (considering stock populations, source populations, side populations and latency populations) is suited to analyse the current situation and further perspectives. The understanding of functional correlations provides the base for a sensitive and sustainable usage of means in species conservation. Two examples in the federal state of Baden-Württemberg show the detailed application.
- Veröffentlicht am

Thick shelled river mussels in a transect of 5 m in the Eschach River. Michael Pfeiffer
1 Einleitung
Wildlebende Tiere und Pflanzen sind typischerweise diskontinuierlich im Raum verbreitet und das umso ausgeprägter, je enger ihre Bindung an ganz bestimmte geologische, geographische und klimatische Faktoren ist. Die Nutzung der Landschaft durch den Menschen und ihre Ausstattung mit technischer Infrastruktur führt zu einer zunehmenden Fragmentierung der Lebensräume. Habitatfragmentierung, ein aus der Inselbiogeographie (McArthur & Wilson 1967) abgeleitetes Paradigma im Naturschutz, gilt als eine wesentliche Bedrohung für die Überlebensfähigkeit vieler Arten und Lebensgemeinschaften (z.B. Fahrig & Rytwinski 2009, Harrison & Bruna 1999). Das trifft auch für Gewässerlebensräume zu (Zwick 1992).
Relevante Bedingungen, unter denen Arten in fragmentierten Lebensräumen existieren können, werden im Wesentlichen in zwei Konzepten formuliert. Zum einen ist es das der kleinsten überlebensfähigen Population (minimum viable population, MVP; Shaffer 1981), zum anderen das der Metapopulation (Hanski 1999, Levins 1969). Theoretische und empirische Studien zu beiden Konzepten gibt es vor allem für terrestrische, aber auch für aquatische Arten (z.B. Jacob et al. 2009, Schiel & Buchwald 2001, Settele et al. 2005, Stegner 2006), weit weniger jedoch für Arten der Fließgewässerlebensräume (z.B. Mari et al. 2014, Terui et al. 2014, Wanzenböck 2004).
Die Situation vieler Fließgewässerorganismen lässt sich mit den Eigenschaften einer Metapopulation beschreiben. Kennzeichnend hierfür ist ein Verbund von (Sub-) Populationen, von denen einzelne aussterben und auch neu gebildet werden können und zwischen denen gelegentlich, aber regelmäßig Individuen ausgetauscht werden (Settele 1998: 14). Daraus lassen sich wichtige Gesichtspunkte für Schutzkonzepte für bedrohte Arten und die Priorisierung von Maßnahmen ableiten (Akçakaya et al. 2007). Das soll nachfolgend am Beispiel der Bachmuschel (Unio crassus Philipsson 1788; Abb. 1) gezeigt werden.
2 Gesetzlich verankert: der Schutz von Individuen und Populationen
Gegenstand des Artenschutzes auf europäischer und nationaler Ebene sind Individuen und Populationen bzw. die lokale Population einer Art (z.B. FFH-Richtlinie Art.1 Ziffer i); BNatSchG §44 Abs. 1 Nr. 2). In diesem Sinne werden auch Populationen der Bachmuschel auf der Ebene lokaler Bestände beschrieben, die ihren Namen dann meist einem konkreten Bach- oder Flusslauf verdanken.
Sind Zuflüsse ebenfalls besiedelt, dann werden deren Muscheln als eigenständige Population betrachtet. Die Populationen werden über ihre Verbreitungsgrenzen und die Bestandsgröße charakterisiert. Beide Parameter sind bei der Anwendung der bisher empfohlenen Methode (z.B. Kobialka & Colling 2006) kritisch zu sehen. Pfeiffer & Nagel (2010) zeigten das für die Ableitung der Populationsgröße aus Stichproben auf. Die Nennung von Verbreitungsgrenzen legt zudem den Schluss nahe, dass die Populationsdichte vom Zentrum der Verbreitung zu Ober- und Untergrenze hin abnimmt. Das muss nicht der Fall sein und ist in jedem Fall durch eine hinreichende Probendichte zu untersuchen.
Ließe sich die Mindestgröße einer überlebensfähigen Population angeben, so wäre das ein wichtiger Zeigerwert für den Schutz gefährdeter Arten. In einer Meta-Analyse ermittelten Traill et al. (2007: 164) als Faustzahl den Wert von einigen Tausend Tieren (ca. 5000). Diese Analyse blieb nicht unwidersprochen (Flather et al. 2011), so dass ihre Anwendbarkeit aktuell unklar ist. Bisher wurde auch noch kein Wert für die kleinste überlebensfähige Population einer Muschelart bestimmt. Die vielfache Abhängigkeit der Flussmuscheln von biotischen (z.B. Wirtsfische) und abiotischen (z.B. Wasser- und Substratqualität) Faktoren sowie der Einfluss von Verteilung und lokaler Dichte der Tiere sowie der Strömung auf den Reproduktionserfolg machen diese Bestimmung sehr schwierig (Haag 2012: 394). Im Extremfall könnte ein einziges zwittriges Tier einer langlebigen Art diese Größe repräsentieren (z.B. Flussperlmuschel, Margaritifera margaritifera Linnaeus, 1758), doch angesichts unvorhersehbarer negativer Einflüsse wäre das für praktische Zwecke ohne Belang (vgl. Nagel 2002a: 71, 72; 2009: 30).
3 Die Metapopulation als realistisches Modell für die Verbreitung der Bachmuschel und anderer Flussmuschelarten
3.1 Die Fragmentierung der Bestände
Die ungleiche Verteilung von Flussmuscheln mit lokalen Häufungen (Muschelbänken) und spärlich oder nicht besiedelten Zwischenstrecken ist ein bekanntes Phänomen, welches die Heterogenität des Lebensraums Fließgewässer, in geringerem Ausmaß auch der Stillgewässer, widerspiegelt (Strayer et al. 2004: 434f.). Individuelles Verhalten (aktive Wanderungen) und populationsökologische Prozesse (Verdriftung, Auslöschung, Neu- und Wiederbesiedlung) von Muscheln und ihren Wirtsfischen tragen ebenfalls zu einer räumlichen und zeitlichen Fragmentierung von Flussmuschelpopulationen bei (Pfeiffer & Nagel 2010: 172-174; Zajac & Zajac 2011: 457-459).
3.2 Der genetische Austausch
Der genetische Austausch zwischen lokalen Populationen kann in Fließrichtung durch Spermiendrift oder die Verlagerung von juvenilen und adulten Tieren bei Flutereignissen stattfinden. Gegen die Fließrichtung können sich die temporär an Wirtsfischen parasitierenden Larven (Glochidien) ausbreiten. Die bei diesen Vorgängen überwindbaren Distanzen sind nicht konkret bekannt. Für die Ausbreitung über das Stadium der Glochidien können sie aber aus den Wanderdistanzen der betreffenden Fischarten abgeschätzt werden (z.B. Terui et al. 2014). Ein wichtiger Vorgang sind z.B. die Laichwanderungen der Elritzen, eines bedeutenden Wirtsfisches, die zeitgleich mit der Glochidienabgabe der Bachmuschel stattfinden. Hierbei ist eine Größenordnung von mehreren hundert Metern realistisch, für andere Arten (z.B. Döbel) auch mehrere Kilometer.
3.3 Aussterben, Neubesiedlung und Wiederansiedlung
Das Verschwinden und Entstehen von Habitatflächen ist die unmittelbare Konsequenz einer natürlichen Fließgewässerdynamik, die Sedimentumlagerungen, Uferabbrüche und selbst das Trockenfallen und Verlanden von Fließstrecken mit sich bringt. Diese hydrologischen Eigenschaften von Bächen und Flüssen zwingen ihre Bewohner zu regelmäßigen Standortwechseln. Dabei sind Fließgewässer zwar offene, dynamische und sehr komplexe Systeme auf der zeitlichen Skala betrachtet sind die Verhältnisse jedoch sehr stabil (Vannote et al. 1980). Auch in Stillgewässern entstehen und vergehen geeignete Habitate, dort aber tendenziell in längeren Zeiträumen. Das Verschwinden oder Absterben von lokalen Muschelvorkommen dürfte daher in den historischen Fließgewässerlandschaften nicht selten gewesen sein. Um solche Verluste kompensieren und neu entstehende Habitate besiedeln zu können, verfügen Flussmuscheln über angepasste Überlebensstrategien. Durch sie ist sowohl lokal das kurzfristige Überdauern als auch regional die Gründung neuer Populationen möglich (Nagel 2002: 264; 2009: 29f.). Besondere Bedeutung kommt dabei den jeweils verfügbaren Wirtsfischen zu, deren Wanderverhalten die Ausbreitung der Flussmuscheln und die Besiedlung neuer Habitate durch Jungmuscheln bestimmt (Verzeichnis von Wirtsfischen in Nagel 2009: 26).
In vielen Gewässern sind Döbel und Elritze die wichtigsten Wirtsarten für Unio crassus. Besonders die Elritze ist auch in Laborversuchen und als Träger von Glochidien in Schutzprojekten für die Bachmuschel eine sehr geeignete und häufig verwendete Art. Der klein bleibende Schwarmfisch besiedelt alle Typen von Fließgewässern, bevorzugt aber überströmte Areale mit einem heterogenen Gemisch aus Feinsedimenten. Die Elritze dringt bis in die Oberläufe von Bächen vor und war früher weit verbreitet und häufig. Damit stimmt die Beobachtung überein, dass junge Bachmuscheln oft in den bevorzugten Aufenthaltsbereichen der Elritze gefunden werden (eigene Beobachtungen; vgl. Abb. 2).
4 Die Elemente einer Metapopulation: Beispiele aus Baden-Württemberg
Populationen unterscheiden sich in Größe und Altersstruktur. Große (Sub-) Populationen mit natürlicher Reproduktion können dabei als Stamm- oder Quellpopulation für die Besiedlung entfernterer Habitate dienen (source). Von kleinen (Sub-) Populationen mit unausgeglichener Altersstruktur oder ohne Nachwuchs kann vermutet werden, dass sie evtl. nicht stabil sind und aussterben (Latenzpopulation, sink).
4.1 Stamm- oder Quellpopulation
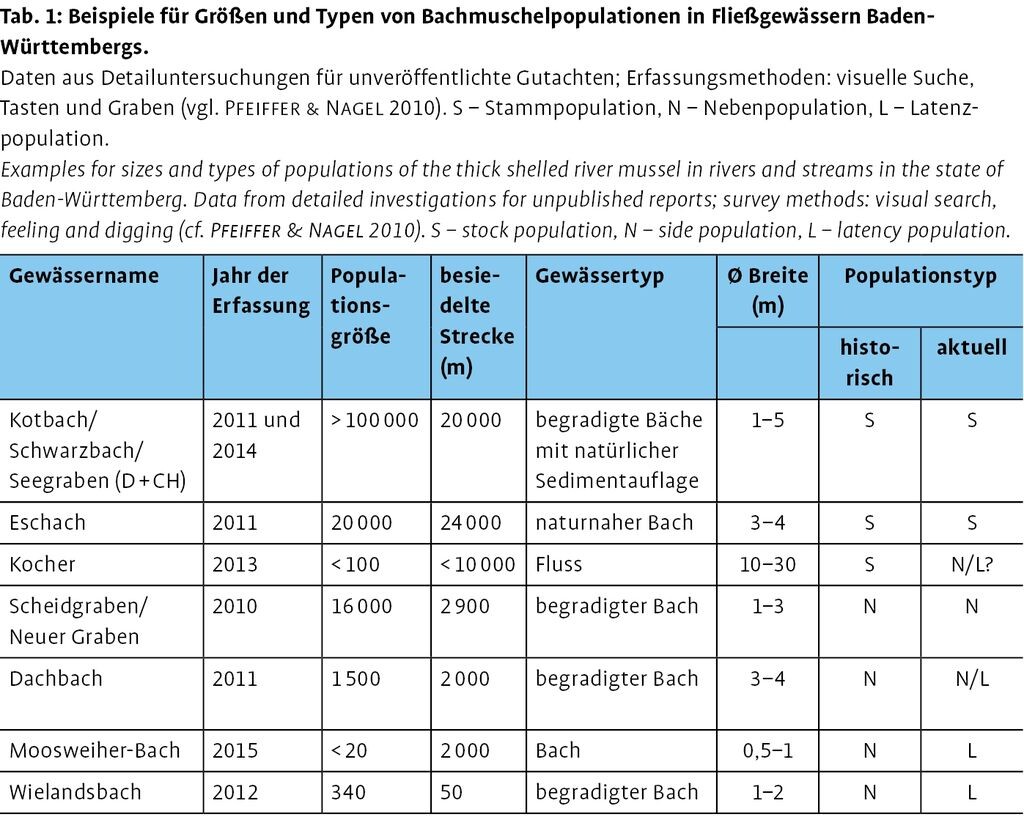
Charakteristisch für eine Stammpopulation sind eine hohe Abundanz (in unbeeinträchtigten Gewässern bis weit über 100000 Tiere) und ein ausgedehnter Lebensraum (mit >10km besiedelter Gewässerstrecke), wobei die Populationsdichte lokal auch gering sein kann (Tab.1). Außerdem sind Stammpopulationen über lange Zeiträume hinweg (Jahrzehnte) stabil. Lokal auftretende Verluste z.B. nach Hochwasser werden durch interne Neuansiedlungen kompensiert. Aktuell sind solche als Stammpopulationen zu bezeichnenden Bestände noch in einigen der großen Zuflüsse des Oberrheins bekannt (Alte Rench, Alte Elz, Schutter/Unditz, Mühlbach/Sulzbach), im großen Neckarzufluss Eschach sowie grenzüberschreitend mit der Schweiz in einem 20km langen Bachsystem im Klettgau.
Viele Stammpopulationen sind inzwischen erloschen. In Baden-Württemberg betrifft das die großen Ströme und Flüsse Donau, Rhein und Neckar. Nur selten gibt es noch Hinweise auf Restbestände (unbekannter Größe), beispielsweise im großen Neckarzufluss Kocher (Tab.1).
4.2 Nebenpopulation
Die meisten der aktuell bekannten Bachmuschelpopulationen leben in kleineren Fließgewässern. Ihre Bestandszahlen bewegen sich in einem mittleren Bereich (500 bis 5000 Individuen, kleine bis mittelgroße Populationen nach Nagel 2002: 71). Die Habitate der Muscheln sind in diesen Gewässern weniger gepuffert und daher anfälliger gegenüber natürlichen (Temperaturschwankungen, Sedimentumlagerungen, Wasserführung, zeitliche und räumliche Verfügbarkeit von Nahrung und Wirtsfischen) oder vom Menschen verursachten (Verschmutzung, Eingriffe in Bodensubstrat und Ufer) Einflüssen. Dementsprechend muss hier mit größeren Abundanzschwankungen gerechnet werden.
In der Kulturlandschaft bieten oft kanalisierte Gewässerstrecken und Gräben für Flussmuscheln günstige Habitatbedingungen. Stabile Substrate, eine von Extremereignissen abgekoppelte Wasserführung und eine bei entsprechender Wasserqualität individuenreiche Fischfauna lassen große Populationen auf klar abgrenzbaren, oft kurzen Strecken entstehen. Katastrophale Ereignisse wie z.B. Trockenperioden, Vergiftungen, Gewässerräumungen oder Bisamfraß können unter diesen Umständen leicht bestandsgefährdend werden, da Rückzugsräume wie in dynamischen Gewässersystemen fehlen. Solche als Nebenpopulationen charakterisierbaren Population sind vor allem in den Einzugsgebieten der oben genannten Stammpopulationen und mittlerweile auch vollkommen isoliert in zahlreichen anderen Gewässersystem anzutreffen. Hat eine solche Nebenpopulation noch eine Verbindung zu einer Stamm- oder anderen Nebenpopulation, so kann sie sich selbst nach katastrophalen Ereignissen immer wieder regenerieren. So profitiert etwa der Bestand in Scheidgraben und Neuem Graben in der Oberrheinebene bei Offenburg (vgl. Tab.1), der regelmäßigen Grabenräumungen ausgesetzt ist, von möglichen Zuwanderungen mittels glochidieninfizierter Fische aus der Population in der Unditz. Im nur unwesentlich kleineren Dachbach bei Heilbronn ist die Population hingegen sehr stark gefährdet. Die ehemalige Nebenpopulation (eine Stammpopulation besiedelte historisch den Lein) ist inzwischen vollkommen isoliert, ein weiterer Bestandseinbruch wäre nicht mehr zu kompensieren.
4.3 Latenzpopulation
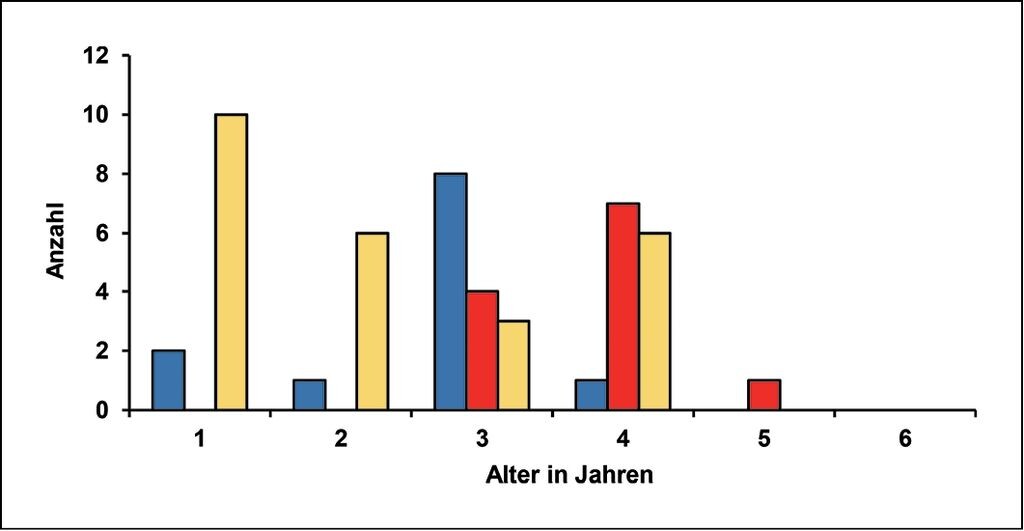
Zahlreiche kleine Zuläufe (Bäche, Gräben) in der Peripherie eines von Bachmuscheln besiedelten Gewässersystems beherbergen nur noch sehr kleine Populationen (<100 Tiere), gelegentlich sogar nur Einzeltiere (Tab.1). Solche Bestände sind vermutlich auf ständige Neuansiedlungen aus Stamm- oder Nebenpopulationen angewiesen. Sind diese nicht (mehr) vorhanden, dann finden sich manchmal noch Leerschalen als letzte Hinweise auf eine frühere Besiedlung. Die Gefahr, dass solche Latenzpopulationen verschwinden, ist groß. Beispiele liefern der Moosweiherbach und der Wielandsbach (Tab.1). Dort leben verstreut auf längerer Strecke nur noch wenige Tiere (Moosweiherbach) bzw. ein kleiner Bestand konzentriert sich auf die letzte kurze Strecke mit ausreichender Habitatqualität (Wielandsbach). Am Oberrhein konnte im verzweigten Gewässersystem der Renchniederung an einigen Stellen auch das Entstehen solcher Latenzpopulationen dokumentiert werden (Rupp 1996-2001 und eigene Beobachtungen, Abb. 3). Die blauen Säulen in Abb. 3 stehen für eine Muschelgruppe, die auf ca. 1 m² Fläche und vollkommen isoliert angetroffen wurde. Exakt in diesem Bereich standen zahlreiche Wirtsfische (einjährige Döbel) in der Strömung. Gelbe Säulen symbolisieren die Besiedlung einer 4m² großen, stabilen Schlammbank, rote Säulen die Besiedlung einer ca. 200 m langen Bachstrecke, die offenbar innerhalb von drei Jahren erfolgte. Die meisten Tiere wurden in der Nähe eines Kolks (Fischeinstand!) eng beieinander sitzend angetroffen.
Drei Wiederansiedlungen teils mehrere hundert Meter entfernt von bekannten Beständen gehören ebenfalls hierher. Gemeinsam ist diesen Fundstellen, dass es bevorzugte Aufenthaltsorte des Döbels waren, eines hier offensichtlich wichtigen Wirtsfisches der Bachmuschel.
5 Die Entwicklung von Metapopulationen: Beispiele aus Baden-Württemberg
5.1 Der Oberlauf eines naturnahen und verzweigten Bachsystems (Eschach) (Abb. 4 und 5)
Im karbonatreichen Einzugsgebiet der Eschach, einem Zufluss zum oberen Neckar, bleiben die Leerschalen von Großmuscheln sehr lange erhalten. Dadurch kann das ehemalige Verbreitungsbild noch heute nachvollzogen werden. Eine große Stammpopulation war von mehreren Neben- und Latenzpopulationen umgeben. Der Hauptlauf der Eschach war vermutlich noch bis in den 1960er-Jahren von der Quellregion bis zur Mündung in den Neckar besiedelt. Mittlerweile bricht die Besiedlung im Bereich eines früher besiedelten großen Bachs (2) abrupt ab. Eine natürliche Wiederansiedlung der flussabwärts verwaisten Gebiete ist momentan nicht möglich, da der vitale Rest der Stammpopulation (1a) vom Unterlauf fast vollkommen abgeschnitten ist (Abb. 4). Bereits in einem so vergleichsweise einfach strukturierten Gewässersystem spielen Austauschprozesse selbst zwischen Teilen einer Stammpopulation eine große Rolle für die Stabilität eines Bestandes im gesamten potenziellen Verbreitungsgebiet.
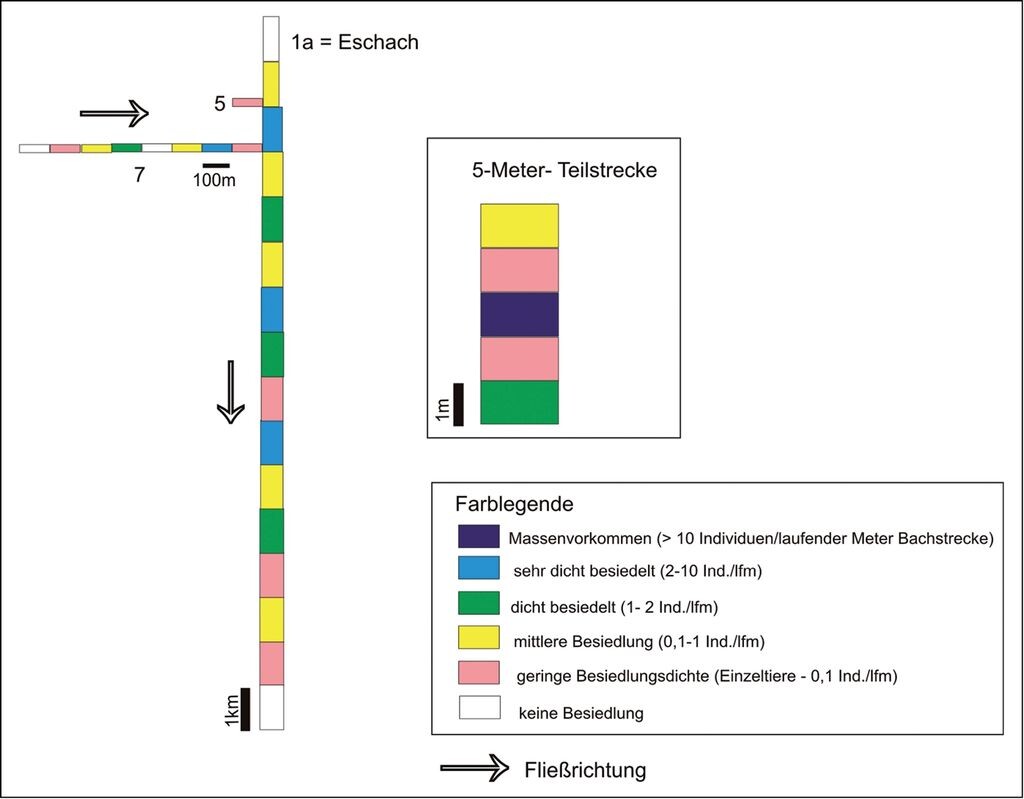
In den aktuell besiedelten Gewässerstrecken (1a, 5, 7) wechseln sich dicht besiedelte mit kaum besiedelten Abschnitten ab (Abb. 5). Die Siedlungsdichten schwanken erheblich (um mehrere Zehnerpotenzen), aber es gibt kaum unbesiedelte Fließstrecken in diesem naturnahen, strukturreichen und dynamischen Teil des Gewässersystems.
5.2 Ein vernetztes Gewässersystem am Oberrhein (Renchniederung) (Abb. 6, 7 und 8)
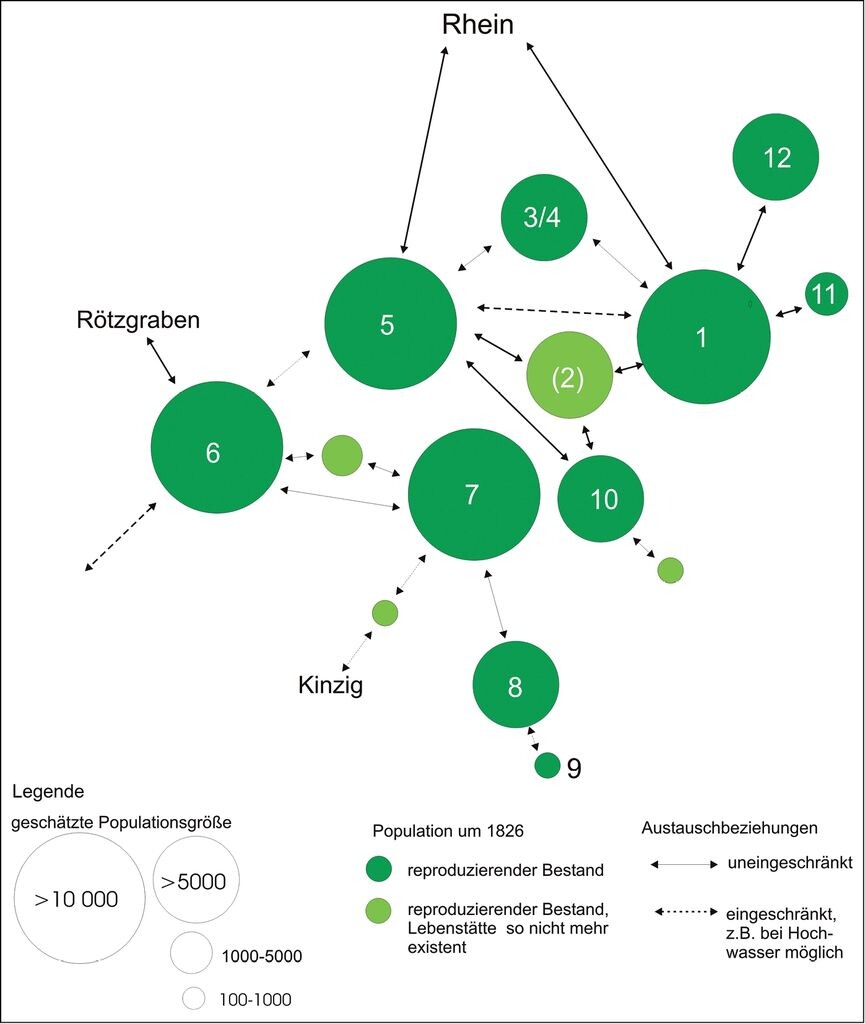
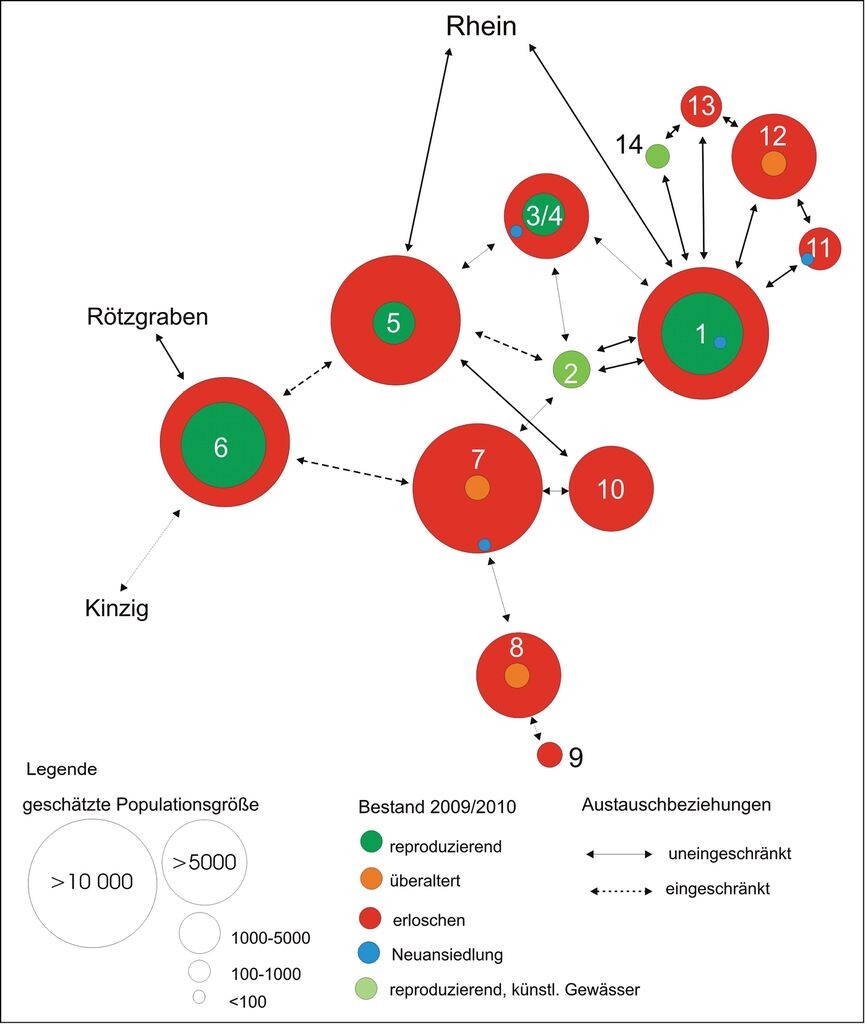
Weitaus komplizierter ist die Situation in den großen Bächen in der Oberrheinniederung wie Alter Rench, Kinzig, Schutter oder Mühlbach/Sulzbach. Für die Rekonstruktion der historischen naturräumlichen Situation im Schwemmfächer der Rench wurde die Topographische Charte von Schwaben aus den Jahren 1825/1826 ausgewertet (vgl. Fischer 1993). Funde von alten Leerschalen (z.B. im Oberlauf von Bach 11) und die Restvorkommen in inzwischen vollständig verbauten Bächen bestätigen dort das erschlossene historische Verbreitungsmuster. Das weitläufige Gewässernetz beherbergte noch vor 150 Jahren vermutlich mehrere Hauptpopulationen (1, 5, 6, 8, 12), die zudem meist miteinander in Verbindung standen (Abb. 6).
Dieser Verbund garantierte ein hohes Maß an Stabilität, so dass auch natürliche Katastrophen (Hochwässer) keine nachhaltigen Bestandseinbußen verursachen konnten, ebenso wie anthropogene Belastungen in der vorindustriellen Kulturlandschaft. Heute sind einige historische Lebensstätten nicht mehr vorhanden und im Zuge der Acher-Rench-Korrektion zwischen 1936 und 1967 wurden einige Gewässer grundlegend verändert: vollständiger Sohl- und Uferverbau (6, 7), Querverbau (z.B. 1, vgl. auch Röck 2006, 2008). In den 1960er-Jahren kam es zur dauerhaften Verschlechterungen der Wasserqualität in einigen Gewässern (z.B. 8, 10), was zu großen Bestandseinbußen führte. Abb. 7 zeigt den Zustand 2010. Lediglich drei der großen Haupt- und Nebenpopulationen sind noch vorhanden (1, 3, 6), andere einst sehr große Bestände (7, 8, 10) reproduzieren nicht mehr oder sind bereits ausgestorben. Mehrere kleinere Nebenpopulation (z.B. 9) sind ebenfalls erloschen. Zwei neu geschaffene Gewässer (2, 14) wurden von Bachmuscheln besiedelt. Vor allem die Population im Rench-Flutkanal (2), der sich an einem historischen Bachlauf orientiert, dürfte momentan einen wichtigen Beitrag zum Überleben der Gesamtpopulation leisten. Mit dem Anschluss des Acher-Flutkanals (13) wurde eine weitere Lebensstätte in das System integriert, die Population ist aber inzwischen wieder erloschen.
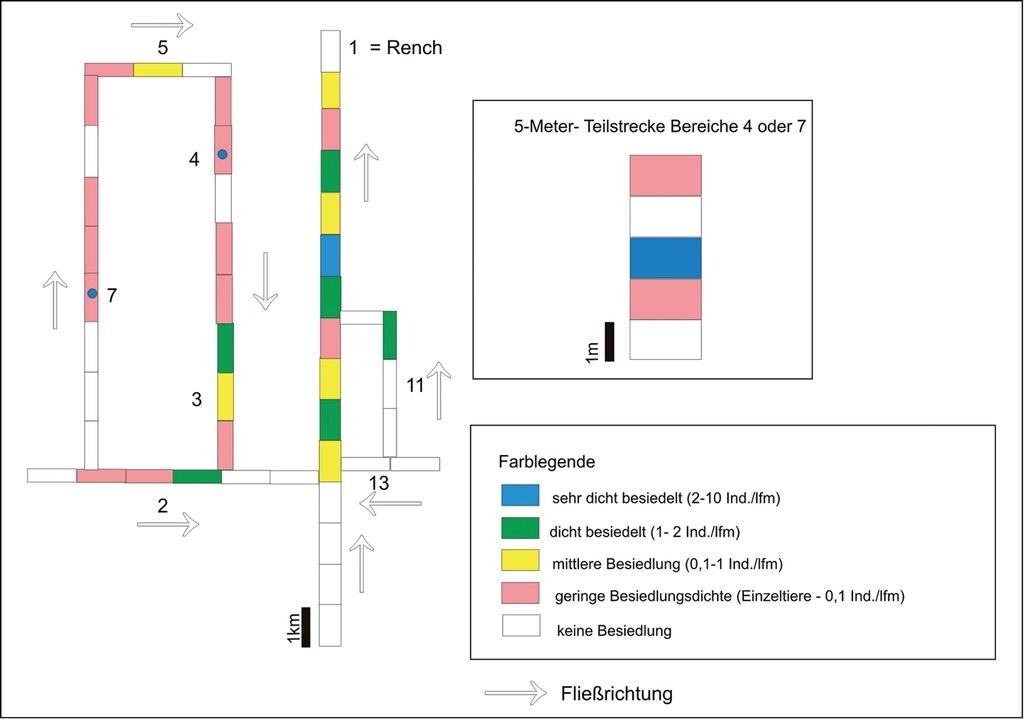
Auch in diesem System wechseln sich Abschnitte mit sehr unterschiedlichen Besiedlungsdichten ab, sowohl im kleinen (5m) als auch im größeren Maßstab (100 m, 1 km) (Abb. 8): Die Stammpopulation in der Rench weist auf einer längeren Strecke wechselnde Dichten auf. Zahlreiche Nebenpopulationen sind miteinander vernetzt, zuweilen liegen aber mehrere hundert Meter oder sogar Kilometer zwischen den Subpopulationen bzw. Muschelgruppen. Wechsel zwischen dicht besiedelten Strecken, Anhäufungen (Neuansiedlungen) und muschelfreien Abschnitten sind erkennbar.
6 Diskussion
Die Bachmuschel lebt heute in weiträumig isolierten Populationen oder Beständen mit wenigen Muscheln bis hin zu nur noch einzelnen Tieren. Beispielhaft gilt dies für einige größere Bachsysteme am Oberrhein in den Niederungen von Rench, Kinzig, im Sulzbachsystem sowie in der Freiburger Bucht (Dreisam-Glotter-System) (AG Mollusken BW 2009, Pfeiffer 2011; Pfeiffer & Nagel 2010: 175; Pfeiffer briefl. 2009). Auch angesichts begrenzter Ressourcen erfordert diese Situation ein Nachdenken über Ansätze und effiziente Maßnahmen zum Schutz der Art. Vereinzelt finden sich bereits Hinweise darauf, dass für die Analyse und Bewertung der Bestandssituation der Bachmuschel der Blick über einzelne (Sub-) Populationen hinausgehen muss. Zettler et al. (1994: 33, 35) bezeichnen lokale Häufungen von mehreren hundert bis tausend Tieren im Verlauf der Warnow als Subpopulationen und regen die Untersuchung der (abiotischen) Dynamik zur Klärung des Verbreitungsmusters an. Auch Pfeiffer & Nagel (2010: 174) und Zajac & Zajac (2011: 459) schlagen vor, die Verteilung von Bachmuscheln im Kontext einer Metapopulation zu untersuchen. Strayer et al. (2004: 435) weisen ausdrücklich auf das Modell der Metapopulation zur Erklärung der räumlichen Interaktion von (Sub-) Populationen hin, das aber bei Flussmuscheln noch wenig angewendet werde.
Das Bundesnaturschutzgesetz (BNat-SchG 2012) kennt den Metapopulationsgedanken (§1 Abs. 2 Nr. 1 BNatSchG), auch Runge et al. (2009: 17) weisen darauf mit Beispielen hin. Für den Schutz einer (Sub-)Population sind die Kenntnis der genauen (aber zeitlich durchaus variablen) Verbreitungsgrenzen sowie das Verständnis von Interaktionen oder gar Abhängigkeiten entscheidend. Ein primäres Schutzziel muss daher die Vernetzung von bestehenden Populationen sein, mit einem besonderen Augenmerk auf mögliche oder zu entwickelnde Stammpopulationen. Hierbei könnte z.B. die Wiederanbindung kleiner Nebengewässer und ihre Reaktivierung als Laichbiotop für die Elritze die Erfolgsaussichten für den Erhalt einer Bachmuschel-Populationen wesentlich verbessern. Es ist kein Zufall, dass Jungmuscheln bevorzugt in typischen Laichstrukturen (Kiesbänke) wichtiger Wirtsfischarten gefunden werden (Abb. 2). Mari et al. (2014: 432) zeigten auch, dass in einem verästelten Netzwerk (Gewässernetz) eine Metapopulation eine höhere Überlebenswahrscheinlichkeit hat.
Literatur
AG Mollusken BW (Arbeitsgruppe Mollusken Baden-Württemberg am staatlichen Museum für Naturkunde Stuttgart, 2009): Ersteinrichtung der Monitoringstrecken und erste Datenerhebung für das FFH-Stichprobenmonitoring der Bachmuschel (Unio crassus, PHIL. 1788). Unveröff. Gutachten i.A. der Landesanstalt für Umwelt, Messungen und Naturschutz (LUBW) Baden-Württemberg.
Akçakaya, H.R., Mills, G., Doncaster, C.P. (2007): The role of metapopulations in conservation. In: Macdonald, D.W., Service, K., eds., Key Topics in Conservation Biology, Blackwell, 64-84.
Bundesnaturschutzgesetz vom 29. Juli 2009 (BGBl. I S. 2542), das zuletzt durch Artikel 5 des Gesetzes vom 6. Februar 2012 (BGBl. I S. 148) geändert worden ist.
Fahrig, L., Rytwinski, T. (2009): Effects of roads on animal abundance: an empirical review and synthesis. Ecology and Society 14 (1), 21 [online].
Fischer, H. (1993): Die Charte von Schwaben 1: 86 400. Cartographica Helvetica 7, 3-10.
Flather, C.H., Hayward, G.D., Beissinger, S.R., Stephens, P.A. (2011): Minimum viable populations: is there a magic number for conservation practitioners? Trends in Ecology and Evolution 26, 307-316.
Hanski, I. (1998): Metapopulation dynamics. Nature 396, 41-49.
(1999): Metapopulation Ecology. Oxford University Press. IX, 313S.
Harrison, S., Bruna, E. (1999): Habitat fragmentation and large-scale conservation: what do we know for sure? Ecography 22, 225-232.
Jacob, A., Scheel, B., Buschmann, H. (2009): Raumnutzung in einer Metapopulation der Gelbbauchunke (Bombina variegata) an ihrer nördlichen Verbreitungsgrenze. Zeitschr. f. Feldherpetologie 16, 85-102.
Kobialka, H., Colling, M. (2006): Weichtiere (Mollusca). In: Schnitter, P., Eichen, C., Ellwanger, G., Neukirchen, M., Schröder, E., Bearb., Empfehlungen für die Erfassung und Bewertung von Arten als Basis für das Monitoring nach Artikel 11 und 17 der FFH-Richtlinie in Deutschland, Ber, LandesamtUmweltschutz Sachsen-Anhalt, Sonderh. 2, 100-111.
Levins, R. (1969): Some demographic and genetic consequences of environmental heterogeneity for biological control. Bulletin of the Entomological Society of America 15, 237-240.
MacArthur, R.H., Wilson, E.O. (1967): The theory of island biogeography. Princeton University Press, Princeton, NJ, 203 pp.
Mari, L., Casagrandi, R., Bertuzzo, E., Rinaldo, A., Gatto, M. (2014): Metapopulation persistence and species spread in river networks. Ecology Letters 17, 426-434.
Nagel, K.-O. (2002a): Die Flussperlmuschel (Margaritifera margaritifera L. 1758) in Hessen: Schutzstrategien aus Erkenntnissen über sehr kleine Populationen. Jahrb. Naturschutz in Hessen 6 (2001), 70-82.
(2002b): Muschel, Mensch und Landschaft Zusammenhänge zwischen Landnutzung und Bestandsentwicklung bei Flussmuscheln. Naturschutz und Landschaftsplanung 34 (9), 261-269.
Pfeiffer, M. (2011): Großmuschelbestände im Regierungsbezirk Freiburg. Dokumentation von aktuellen Funden und kurze Einschätzung der Situation in verschiedenen Regionen. Unveröff. Gutachten i.A. des Regierungspräsidiums Freiburg.
, Nagel, K.-O. (2010): Schauen, tasten, graben Strategien und Methoden für die Erfassung von Bachmuscheln (Unio crassus). Naturschutz und Landschaftsplanung 42 (6), 171-179.
Röck, S. (2005): Der Rench-Flutkanal. Die Natur eines künstlichen Gewässers. In: Ohlig, C., Hrsg., Gewässerentwicklung in der Kulturlandschaft, Schr. Deutsche Wasserhistor. Ges. (DWhG) 7, 374S.
(2008): Naturqualität und Bewertung künstlicher Gewässer am Beispiel zweier Flutkanäle in der Oberrheinebene. Culterra 53, 179 S. + 38S. Anhang.
Runge, H., Simon, M., Widdig, T. (2009): Rahmenbedingungen für die Wirksamkeit von Maßnahmen des Artenschutzes bei Infrastrukturvorhaben. FuE-Vorhaben im Rahmen des Umweltforschungsplanes des Bundesministeriums für Umwelt, Naturschutz und Reaktorsicherheit im Auftrag des Bundesamtes für Naturschutz FKZ 3507 82 080 (unter Mitarbeit von: Louis, H.W., Reich, M., Bernotat, D., Mayer, F., Dohm, P., Köstermeyer, H., Smit-Viergutz, J., Szeder, K.). Hannover, Marburg.
Rupp, L. (1996-2001): Programm zur Erfassung, Erhaltung und Wiederansiedlung der Bachmuschel (Unio crassus Phil. 1788) im Regierungsbezirk Freiburg. Unveröff. Ber. i.A. des Regierungspräsidiums Freiburg und der Bezirksstelle für Naturschutz.
Schiel, F.-J., Buchwald, R. (2001): Die Große Moosjungfer in Südwest-Deutschland: Konzeption, Durchführung und Ergebnisse des LIFE-Natur-Projekts für gefährdete Libellenarten am Beispiel von Leucorrhinia pectoralis. Naturschutz und Landschaftsplanung 33, 274-280.
Schnitter, P., Eichen, C., Ellwanger, G., Neukirchen, M., Schröder, E. (Bearb., 2006): Empfehlungen für die Erfassung und Bewertung von Arten als Basis für das Monitoring nach Artikel 11 und 17 der FFH-Richtlinie in Deutschland. Ber. Landesamt f. Umweltschutz Sachsen-Anhalt, Sonderh. 2, 370S.
Settele, J. (1998): Metapopulationsanalyse auf Rasterdatenbasis: Möglichkeiten des Modelleinsatzes und der Ergebnisumsetzung im Landschaftsmaßstab am Beispiel von Tagfaltern. Teubner, Stuttgart, Leipzig, 130S.
, Kühn, E., Thomas, J. A. (2005): Studies on the ecology and conservation of butterflies in Europe. Vol. 2: Species Ecology along a European Gradient: Maculinea Butterflies as a Model. Pensoft, Sofia, 289S.
Shaffer, M.L. (1981): Minimum population sizes for species conservation. BioScience 31, 131-134.
Stegner, J. (2006): Kriterien zur Bewertung des Erhaltungszustandes der Populationen des Eremiten Osmoderma eremita (Scopoli, 1763). In: Schnitter, P., Eichen, C., Ellwanger, G., Neukirchen, M., Schröder, E., Bearb., Empfehlungen für die Erfassung und Bewertung von Arten als Basis für das Monitoring nach Artikel 11 und 17 der FFH-Richtlinie in Deutschland, Ber. Landesamt f. Umweltschutz Sachsen-Anhalt, Sonderh. 2.
Strayer, D.L., Downing, J.A., Haag, W.R., King, T.L., Layzer, J.B., Newton, T.J., Nichols, S.J. (2004): Changing perspectives on pearly mussels, North Americas most imperiled animals. BioScience 54, 429-439.
Terui, A., Miyazaki, Y., Yoshioka, A., Kaifu, K., Matsuzaki, S.S., Washitani, I. (2014): Asymmetric dispersal structures a riverine metapopulation of the freshwater pearl mussel Margaritifera laevis. Ecology and Evolution 4, 3004-3014.
Topographische Charte von Schwaben, Blatt Strassburg (1826), Baden (1825), Offenburg (1826) ( http://idb.ub.uni-tuebingen.de/diglit/FhXIV1a _qt/0011/...12/...20)
Traill, L.T., Bradshaw, C.J.A., Brook, B.W. (2007): Minimum viable population size: A meta-analysis of 30 years of published estimates. Biol. Conserv. 139, 159-166.
Vannote, R.L., Minshall, G.W., Cummings, K.W., Sedelland, J.R., Cushing, C.E. (1980): The river continuum concept. Canadian Journal Fisheries and Aquatic Sciences 37, 130-137.
Wanzenböck, J. (2004): European Mudminnow (Umbra krameri) in the Austrian floodplain of the river Danube: conservation of an indicator species for endangered wetland ecosystems in Europe. In: Akçakaya, H.R., Burgman, M.A., Kindvall, O., Wood, C.C., Sjögren-Gulve, P., Hatfield, J.S. & McCarthy, M.A., eds., Species Conservation and Management: Case Studies, Oxford University Press, 200-207.
Zajac, K., Zajac, T. (2011): The role of active individual movement in habitat selection in the endangered freshwater mussel Unio crassus Philipsson. 1788. Journal of Conchology 40, 446-461.
Zettler, M.L., Jueg, U. (2001) Die Bachmuschel (Unio crassus) in Mecklenburg-Vorpommern. Naturschutzarbeit in Mecklenburg-Vorpommern 44, 9-16.
, Kolbow, D., Gosselck, F. (1994): Die Unioniden im Warnow-Einzugsgebiet unter besonderer Berücksichtigung der Bachmuschel (Unio crassus PHILIPSSON 1788). Naturschutzarbeit in Mecklenburg-Vorpommern 37, 30-39.
Zwick, P. (1992): Stream habitat fragmentation a threat to biodiversity. Biodiversity and Conservation 1, 80-97.
-
Abb. 1: Bachmuscheln aus einem 5-m-Transekt in der Eschach.© Michael Pfeiffer
Thick shelled river mussels in a transect of 5 m in the Eschach River. Michael Pfeiffer -
Abb. 2: Mikrohabitat der Jungmuschel in einem typischen Elritzenstandort.
Microhabitat of the young mussel in a typical habitat of the common minnow (Phoxinus phoxinus).Michael Pfeiffer -
Abb. 3: Beispiele für punktuelle Neuansiedlungen von Bachmuscheln und die Verteilung der Tiere in Altersklassen.
Examples for isolated new settlement of thick shelled river mussels and the age distribution of specimens. -
Abb. 4: Bestandsgrößen und Austauschbeziehungen (Spermiendrift und wandernder Wirtsfisch Elritze), historisch (links) und aktuell (rechts) in der Eschach. Die Ziffern stehen für ein Gewässer bzw. für eine lokale Population. Die Fließrichtung ist jeweils von der höheren zur kleineren Ziffer (z.B. 4 2 1).1a, 1b Eschach (Stammpopulation, 1b ausgestorben), 2 Fischbach (Nebenpopulation, ausgestorben), 3 Teufenbach (Nebenpopulation, ausgestorben), 4 Badische Eschach (Nebenpopulation, ausgestorben), 5 Bannmoosgraben (Latenzpopulation), 6 Kimmichgraben (Nebenpopulation, ausgestorben), 7 Seltenbach (Nebenpopulation). Stock sizes and exchange conditions (drift of sperm and migration of the host fish common minnow), historically (on the left) and currently (on the right) in the Eschach River. Each number stands for a water body or for a local population. The flow direction points from the larger to the smaller number (e.g. 4 2 1).1a, 1b Eschach (stock population, 1b extinct), 2 Fischbach (side population, extinct), 3 Teufenbach (side population, extinct), 4 Badische Eschach (side population, extinct), 5 Bannmoosgraben (latency population), 6 Kimmichgraben (side population, extinct), 7 Seltenbach (side population). -
Abb. 5: Siedlungsdichten der Bachmuschel in der Stammpopulation der Eschach (1a in Abb. 3) und in zwei Nebengewässern (5 und 7 in Abb. 3). Länge der Abschnitte: 1a 1 km, 5, 7 100 m.
Settlement densities of the thick shelled river mussel in the stock population of the River Eschach (1a in Fig. 3) and in two side streams (5 and 7 in Fig. 3). Length of the sections: 1 a 1km, 5, 7 100 m. -
Abb. 6: Rekonstruktion der Metapopulation von Unio crassus in den Niederungen von Rench und Holchenbach im Jahr 1826 (Oberrhein).Die Ziffern stehen für ein Gewässer bzw. für eine lokale Population. Die Fließrichtung ist jeweils von der höheren zur kleineren Ziffer.1 Rench, 2 Hoschgraben, 3 Schwiebergraben, 4 Rehlach, 5 Holchenbach, 6 Fischgießen, 7 Kammbach, 8 Wannenbach, 9 Durbach, 10 Stangenbach, 11 Dorfbach, 12 Plauelbach.
Reconstruction of the metapopulation of Unio crassus in the lowlands of the River Rench and the Holchenbach in 1826 (Upper Rhine River)Each number stands for a water body or for a local population. The flow direction points from the larger to the smaller number.1 Rench, 2 Hoschgraben, 3 Schwiebergraben, 4 Rehlach, 5 Holchenbach, 6 Fischgießen, 7 Kammbach, 8 Wannenbach, 9 Durbach, 10 Stangenbach, 11 Dorfbach, 12 Plauelbach. -
Abb. 7: Die Metapopulation von Unio crassus in der Renchniederung im Jahr 2010 (ca. 20000 Tiere).1 Rench, 2 Rench-Flutkanal, 3 Schwiebergraben, 4 Rehlach, 5 Holchenbach, 6 Fischgießen, 7 Kammbach, 8 Wannenbach, 9 Durbach, 10 Stangenbach, 11 Dorfbach, 12 Plauelbach, 13 Acher-Flutkanal, 14 Schwellengraben.
The metapopulation of Unio crassus in the lowlands of the River Rench in 2010 (about 20.000 individuals). -
Abb. 8: Siedlungsdichten der Bachmuschel in einem Teilbereich des Gewässernetzes in der Renchniederung (Oberrhein; Ziffern wie in Abb. 5). Weitere Erläuterung im Text.
Settlement densities of the thick shelled river mussel in a part of the stream network in the lowlands of the River Rench (Upper Rhine; numbers as in. Fig. 5). Further explanations in the text (only in German). -
Die Bachmuschel (Unio crassus) überlebt noch in Metapopulationen -
Die Bachmuschel (Unio crassus) überlebt noch in Metapopulationen



Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.