Quo vadis Moorfalter?
Abstracts
Schmetterlinge nährstoffarmer Moore sind an kühlere Lebensräume angepasst. Deshalb ist zu befürchten, dass es im Zuge des Klimawandels wegen der ansteigenden Temperaturen zu einem lokalen Aussterben dieser Arten kommen kann. Seit 1990 sind in Mecklenburg-Vorpommern (M-V) bereits Vorkommen von sechs Arten erloschen. Um nachhaltige, naturschutzfachliche Konzepte für den Schutz dieser Arten ausarbeiten zu können, ist es wichtig, ihre ökologische Amplitude gegenüber wesentlichen Klimafaktoren zu kennen. Durch eine Verschneidung der Verbreitungsgebiete von Hochmoor-Arten in Europa mit Datensätzen zu den Isohypsen und Isothermen wurden Erkenntnisse zu einzelnen klimatischen Toleranzbereichen ermittelt.
Die Arten weisen eine relativ einheitliche Toleranz gegenüber den analysierten Faktoren auf. Es besteht somit die Gefahr des gleichzeitigen Erlöschens verschiedener Arten im Zuge des Klimawandels. Vergleicht man die in der Arbeit ermittelten Toleranzbereiche der Taxa mit den Temperaturwerten der Periode 1950 2000 sowie der Klimaprojektion bis 2070, so wird für Mecklenburg-Vorpommern deutlich, dass besonders bei der Jahresmittel- und Minimaltemperatur das Erreichen bzw. Überschreiten der Toleranzgrenzen kurz bevorsteht bzw. sich bereits vollzogen hat.
Abschließend wird ein Beispiel für eine Restaurierung eines Kesselmoores beschrieben, durch die das Überleben des Hochmoor-Scheckenfalters gesichert werden konnte.
Quo vadis butterflies and moths of bogs? Climatic preferences of tyrphobiontic and tyrphophilic species of nutrient-poor bogs in terms of temperature and precipitation in the context of climate change
Lepidoptera of bogs and swamps are adapted to cooler habitats. It is to be feared that these species may face local extinction because of rising temperatures due to climate change. Since 1990, extinction has been reported for 6 species in Mecklenburg-Western Pomerania (M-V).
In order to develop sustainable nature conservation concepts for the protection of these species, it is important to know their ecological amplitude regarding essential climate factors. This amplitude has been determined by linking their distribution areas in Europe to recorded isohypse and isotherm data.
The analysis revealed the following findings: The species have a relatively coherent tolerance range regarding their (median) response towards all factors analyzed. As a consequence, local extinction of various species may occur simultaneously in the course of climate change. Comparing the measured data to those of 1950 - 2000 and to the climate projections until 2070, it becomes clear that especially in terms of annual mean temperatures and minimum temperatures, the tolerance limits of the analyzed species are almost reached in M-V.
As a last point, the paper describes the example of a restored kettlehole which ensured the survival of the Cranberry Fritillary.
- Veröffentlicht am

1 Einführung
Die Existenz stenotoper Schmetterlinge in nährstoffarmen Mooren (Hoch- und Zwischenmoore) ist in besonderer Art und Weise gebunden an
- das Vorhandensein von moortypischen Pflanzengesellschaften (u.a. Zwergstrauch-Torfmoosrasen, Torfmoos-Seggenriede) und Fraßpflanzen (v.a. Moosbeere Vaccinium oxycoccos , Rauschbeere V. uliginosum , Rosmarinheide Andromeda polifolia , Sumpf-Porst Rhododendron tomentosum ),
- ein eher kontinentales" Eigenklima (kühler als Umland, Frühjahr kommt spät, Sommer mit hohen Temperaturen, hohe Verdunstung) und
- spezifische Mikroklimate (Schlenken: hohe Luftfeuchte und ausgeglichene Temperatur; Bulten: trockener, bezüglich der Temperaturen Extremklima).
Solche Moore sind heute als Palaeorefugien für viele stark angepasste Schmetterlingsarten hoch bedeutsam (Sommeret al. 2015,Thieleet al. 2016). Insbesondere unter dem Blickwinkel der prognostizierten weiteren Temperaturerhöhungen (Klimawandel) gilt das für feucht- und kaltstenotope Moorbesiedler, die durch diese Veränderungen zunehmend in Bedrängnis" geraten (Devictoret al. 2012;Thieleet al. 2013, 2014a;Van Dycket al. 2015).
Ist zu den Pflanzengesellschaften und Fraßpflanzen tyrphobionter/-philer Schmetterlingsarten relativ viel bekannt (vgl. Literaturauswertung inThiele & Luttmann2015), so findet man zu den kleinklimatischen Ansprüchen der ektothermen Taxa in der Literatur nur wenig. Einzelne Angaben wie Itame brunneata : kühle Habitate werden bevorzugt" (Ebert2003) lassen sich bezüglich der Abschätzung von Folgen des Klimawandels nur schwer verwerten. Günstiger ist schon die Datenlage für Colias palaeno . Bei dieser Art gebenBräuet al. (2013) einen Super-Cooling-Point der Raupen von 26 °C an und legen den Beginn der kritischen" winterlichen Temperaturen für die Raupen um den Gefrierpunkt fest. Liegen diese darüber, führen Schimmelbefall und Fäulnis unter Umständen zum Absterben der Larven.
Radchuket al. (2013a) nutzten Modelle, um vorauszusagen, wie sich steigende Temperaturen auf die Populationsdynamik von Boloria eunomia auswirken. Sie fanden heraus, dass diese grundsätzlich einen positiven Effekt auf die Fertilität und die Überlebensrate der Tiere hatten. Es gab eine gravierende Ausnahme: Die Larven starben bei höheren winterlichen Temperaturen verstärkt ab. Dies hatte einen überproportional negativen Effekt auf das Populationswachstum. In einer weiteren Studie vonRadchuket al. (2013b) führten die Autoren Experimente durch, um das Verhalten der Raupen unter den unterschiedlichen klimatischen Zukunftsprojektionen zu beobachten. Im Ergebnis zeigte sich, dass die Anzahl gelegter Eier sowie die Überlebensrate von Eiern und Puppen wenig beeinflusst waren. Wegen des wärmeren Winters wurden aber die Raupen deutlich stärker von Krankheiten und Pilzinfektionen betroffen. Das kann eine Population je nach klimatischer Projektion zu 88 bis 97 % reduzieren. Die überwinternden Raupen sind somit das klimasensibelste Stadium.
Systematische Untersuchungen zu den Wirkungen des Klimawandels auf Tagfalter sind durchSetteleet al. (2008) vorgenommen worden. Im Rahmen des Climatic Risk Atlas of European Butterflies" wurden unter Zugrundelegung der klimatischen Projektionen potenzielle, zukünftige Verbreitungsgebiete für die Arten abgeleitet. Dabei sind auch die tyrphobionten/-philen Taxa behandelt worden, soweit sie zu den Tagfaltern gehören. Bezüglich der Nachtfalter sind den Autoren keine vergleichbar konkreten Analysen bekannt.
In den vorliegenden Analysen wurden nun für alle stenotopen Arten nährstoffarmer Moore klimatische Parameter auf Basis ihrer europäischen Verbreitungsgebiete ermittelt. Dabei sind ähnliche Methoden wie beiSetteleet al. (2008) angewandt worden. Die Ergebnisse werden mit klimatischen Toleranzen von Arten unterschiedlicher Verbreitungstypen verglichen. Dadurch lassen sich tyrphobionte/-phile Taxa bezüglich dieses Merkmals einordnen. Zudem werden auf Grundlage von Klimaprojektionen Aussagen getroffen, wie hoch das Risiko ist, dass Arten nährstoffarmer Moore in Mecklenburg-Vorpommern lokal aussterben. Abschließend wird ein Beispiel für die positiven Wirkungen von Moorrestaurierungen auf die Art Boloria aquilonaris vorgestellt. Die Ergebnisse sollen nachfolgend beschrieben und diskutiert werden.
2 Methode
Um die autökologischen Anspruchskomplexe der tyrphobionten/-philen Taxa abzuleiten, wurde adäquat zur Methode vonSetteleet al. (2008) vorgegangen. Dabei wird vorausgesetzt, dass die Verbreitung der Arten nicht zufällig ist. Somit spielen auch artspezifisch determinierte, klimatische Toleranzen eine wesentliche Rolle. Durch Verschneidung der europäischen Verbreitungsgebiete mit aus der Literatur bekannten Isohypsen und Isothermen können klimatische Toleranzbereiche der jeweiligen Taxa abgeleitet werden. Einschränkend muss zu dieser Methode bemerkt werden, dass Temperatur und Niederschlag nur zwei Faktoren für die Verbreitung von Arten darstellen und diese wesentlich über die Menge und Qualität der Nahrung sowie der physiologischen Zeit wirken (Begonet al. 1998). Zudem sind Moore azonale Lebensräume, die ein Eigenklima besitzen, was die Aussagekraft der Toleranzen eingrenzt. Die Abweichungen von Mooreigenklimaten und Umgebungsklimaten sind derzeit wenig untersucht. Vermutlich interagieren aber beide Klimate, so dass die Umgebungstemperatur als Proxy für die Mooreigenklimate dienen kann. Somit muss bei dieser Vorgehensweise immer eine schwer einzuschätzende Unschärfe unterstellt werden. Diese kann solange nicht ausgeräumt werden, wie genauere Untersuchungen zu den klimatischen Präferenzen der Arten für das Freiland fehlen.
Um diese objektiv nicht vermeidbaren Unschärfen in den nachfolgenden Betrachtungen möglichst klein zu halten, bedarf es genauer Verbreitungskarten. Diese existieren aber für die betrachteten Arten nicht in hinreichender Qualität. Deshalb wurden im ersten Schritt die europäischen Verbreitungsgebiete aus Literaturquellen unterschiedlicher europäischer Gebiete miteinander abgeglichen, wobei besondere Beachtung den jeweiligen Gebieten galt, in denen die Bearbeiter vornehmlich tätig waren. Dabei ist von einer relativ ungenauen, aber für alle Arten vorhandenen Grundkarte (Funet2015) ausgegangen worden, in die sukzessive Verbreitungsinformationen u.a. vonHenriksen & Kreutzer(1982),Skou(1991),Marttilaet al. (1992),Inman-Tjørve & Trolle(1999),Saarinen & Jantunen(2003),Trolle & Rubæk(2003),Lafranchis(2004),Leraut(2006),Maceket al. (2007,2008,2012), Aarviket al. (2009),Bühler-Cortesi(2012),Paolucci (2013) undKudnraet al. (2015) eingearbeitet wurden. Hinzu kamen noch zahlreiche Informationen aus einschlägigen Internetquellen (u.a. www.ukbutterflies.co.uk ), auf die an dieser Stelle nicht dezidiert eingegangen werden soll. Diese Karte wurde mit dem Geographischen Informationssystem ArcGIS georeferenziert und in das Koordinatensystem ETRS LAEA (Lambert Azimuthal Equal Area) transformiert.
In einem zweiten Schritt wurden die Verbreitungskarten mit Klimadaten der Worldclim-Datenbank (Hijmanset al. 2005) verschnitten. Über eine Ermittlung der statistischen Flächenverteilung der Klimawerte in den Verbreitungsgebieten sind die Spannweiten und Quantile (inkl. Median) der artspezifischen Toleranzen gegenüber der
- Jahresmitteltemperatur,
- Minimaltemperatur: mittlere Temperatur des kältesten Monats (syn. winterliche Temperatur),
- Maximaltemperatur: mittlere Temperatur des wärmsten Monats (syn. sommerliche Temperatur),
- Temperaturtagessumme > 5° als Parameter, der die Länge und Wärmeverfügbarkeit der Vegetationsperiode bestimmt (syn. Wachstumsgradtage, WGT) sowie
- mittleren Jahresniederschläge
analysiert worden.
Zur Abgrenzung der Klimatoleranzbereiche wurden Interquantilsintervalle (Q0,05;Q0,95) für die jeweiligen Parameter festgelegt. Damit sind unschärfebedingte Extremwerte abgetrennt worden, die durch Unsicherheiten bei der Verbreitungsgebietsermittlung aufgetreten sein könnten und mit hoher Wahrscheinlichkeit außerhalb der Toleranzgrenzen liegen.
Ein dritter Schritt bezog sich auf die Einordnung der klimatischen Toleranzbereiche tyrphobionter/-philer Arten in das Spektrum von Taxa unterschiedlicher Verbreitungstypen (arktisch, arkto-alpin, boreo-montan, boreal, montan, euro-orientalisch, mediterran, tropisch). Da beispielhaft nur Tagfalter betrachtet wurden, konnten die Verbreitungsinformationen ausKudnraet al. (2015) entnommen werden. Die Artauswahl erfolgte innerhalb eines Verbreitungstyps bewusst zufällig. Mit dieser Vorgehensweise lässt sich einerseits überprüfen, ob die erhaltenen Indikatorwerte zu Temperatur- und Niederschlagstoleranzen untereinander plausibel sind. Andererseits können die Werte für die Taxa nährstoffarmer Moore eingeordnet werden, so dass ein Rückschluss auf den Verbreitungstyp gezogen werden kann.
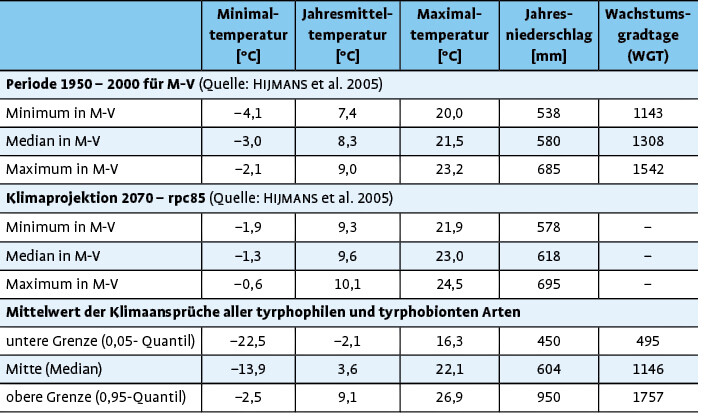
Als Quintessenz wurden die Temperatur- und Niederschlagstoleranzindikatoren mit den einzelnen klimatischen Projektionen für das Gebiet von Mecklenburg-Vorpommern verglichen. Basis waren regionalisierte IPCC5-Szenarien des globalen Klimamodells MPI-ESM des Max-Planck-Instituts für Meteorologie aus dem Coupled Model Incomparison, Project Phase 5 (CMIP5). Es standen eine milde" (rcp45) und eine gravierende" (rcp85) Projektion zur Auswahl, die jeweils unterschiedliche Entwicklungen der Konzentration der klimarelevanten CO2-Äquivalente (Representative Concentration Pathways RCPs) beinhalten. Da die derzeitige Entwicklung mehr dem rcp85-Szenario entspricht (Anstieg auf 8,5 ppm CO2-eq. bis 2070), wurde dieses für die weiteren Betrachtungen zugrunde gelegt. Somit kann festgestellt werden, welche Arten einem besonders hohen klimatischen Risiko bis 2070 (Durchschnitt für 2061-2080) unterliegen. Naturschutzfachlich sollte das gezielte Reaktionen nach sich ziehen, die beschrieben werden.
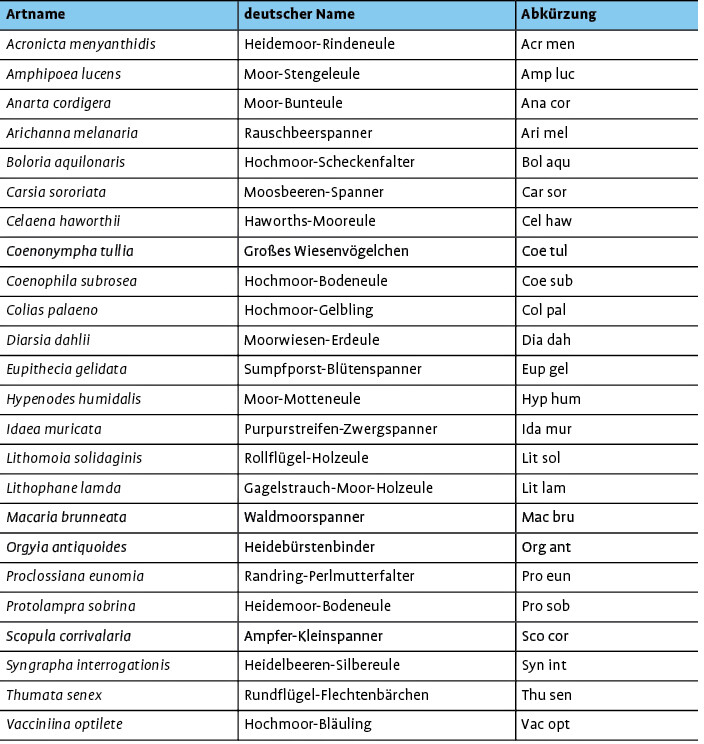
In die Analyse wurde 24 tyrphobionte/-phile Arten einbezogen. Die Artabkürzungen und deutschen Trivialnamen werden in Tab. 1 den wissenschaftlichen Namen gegenübergestellt.
Weiter wurden folgende Arten verschiedener Verbreitungstypen zu Vergleichszwecken herangezogen: Nördliches Posthörnchen ( Colias hecla, arktisch) , Arktischer Rundaugenmohrenfalter ( Erebia polaris, arktisch) , Polarperlmutterfalter ( Boloria polaris, arktisch) , Graumelierter Alpen-Würfel-Dickkopffalter ( Pyrgus andromedae, arkto-alpin) , Ähnlicher Perlmutterfalter ( Boloria napaea, arkto-alpin) , Graubrauner Mohrenfalter ( Erebia pandrose, arkto-alpin) , Freijas Perlmutterfalter ( Boloria freija, boreal) , Friggas Perlmutterfalter ( Boloria frigga, boreal) , Nicias-Bläuling ( Aricia nicias, boreo-montan) , Heller Alpenbläuling ( Agriades orbitulus, boreo-montan) , Bergweißling ( Pieris bryoniae, montan) , Tagpfauenauge ( Aglais io, ubiquitär), Kleiner Fuchs (Aglais urticae, ubiquitär), Kleiner Kohlweißling (Pieris rapae, ubiquitär), Kleiner Alpenbläuling ( Cupido osiris , euro-oriental) , Gelber C-Falter ( Polygonia egea , euro-oriental), Osterluzeifalter ( Zerynthia polyxena , euro-oriental), Schwarzaugenbläuling ( Glaucopsyche melanops , mediterran), Kleopatrafalter ( Gonepteryx cleopatra , mediterran), Pelargonien-Bläuling ( Cacyreus marshalli , tropisch), Gras-Juwel ( Chilades trochylus , tropisch) und Großer Wanderbläuling ( Lampides boeticus , tropisch).
3 Ergebnisse und Diskussion
3.1 Klimatische Toleranzbereiche von Moorarten und Vergleich zur Periode 1950 bis 2000 bzw. Klimaprojektion 2070
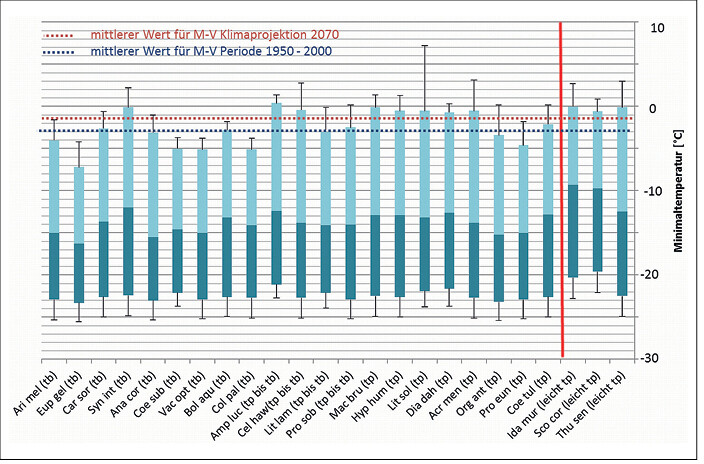
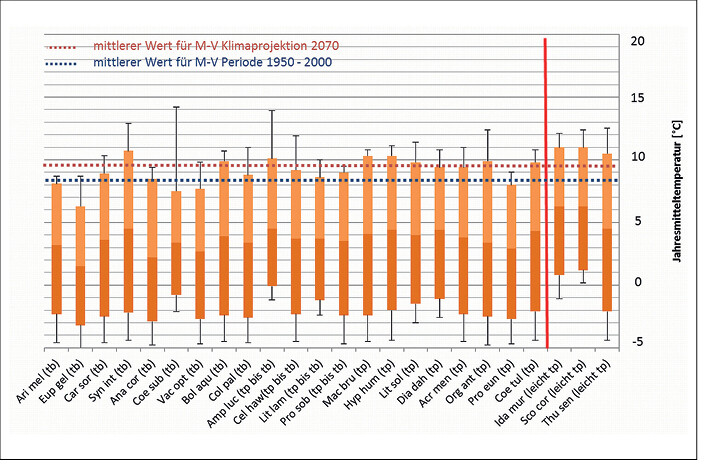
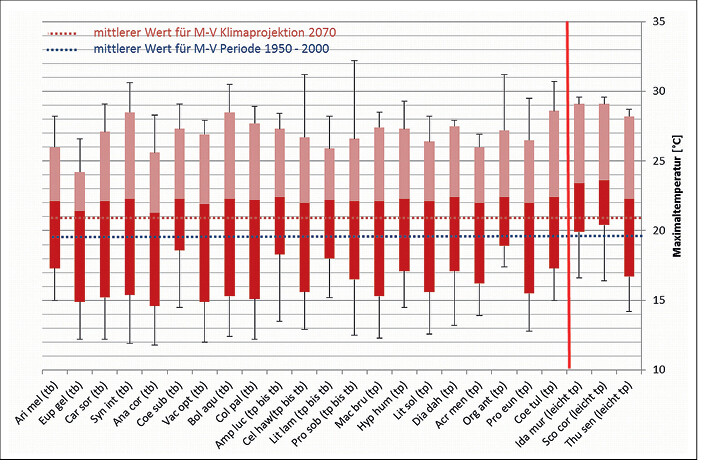
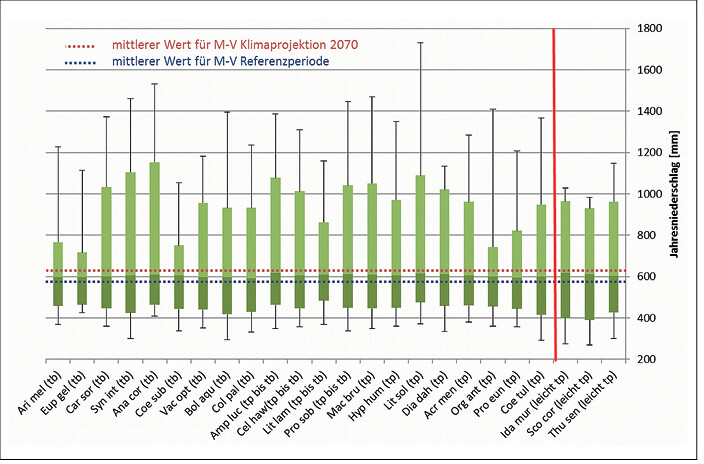
In Abb. 1 bis 4 sind die Toleranzbereiche für die tyrphobionten/-philen Arten bezüglich der untersuchten Klimaindikatoren dargestellt. Um den Übergang zu den Taxa des Moorumlands zu demonstrieren, wurden leicht tyrphophile Arten hinzugenommen. Zudem sind die Mediane für die klimatologische Periode 1950 2000 und die Klimaprojektion 2070 bei starkem Anstieg der Konzentration der CO2-Äqivalente (rcp85) als gestrichelte Linie eingefügt.
Es zeigt sich, dass die tyrphobionten/-philen Arten relativ einheitliche Toleranzbereiche aufweisen. Die Mediane der Werte variieren nur unwesentlich, die Streuungen partiell stärker. In Tab. 2 sind die durchschnittlichen (gerundeten) Minimal-, Median- und Maximalwerte für jeden Klimafaktor aufgeführt. Dagegen gespiegelt wurden die Werte für die klimatologische Periode 1950 2000 und die Klimaprojektion bis 2070. Es zeigen sich folgende Tendenzen:
- Durch die einheitliche Reaktion der Arten gegenüber wesentlichen klimatischen Faktoren besteht die Gefahr, dass mehr oder weniger stark angepasste Schmetterlingspopulationen nährstoffarmer Moorbereiche relativ gleichzeitig lokal aussterben. Dieses Szenario ist umso wahrscheinlicher, da im Tiefland die Arten heute in relativ isoliert liegenden Mooren leben und nicht in höhere und damit kühlere Bereiche ausweichen können.
- Zwischen tyrphophilen und tyrphobionten Arten sind nur geringe Unterschiede in der Toleranz gegenüber klimatischen Faktoren zu konstatieren, d.h. auch diese Taxa unterliegen bei deutlichen klimatischen Änderungen der Gefahr des regionalen Erlöschens der Populationen.
- Leicht tyrphophile Arten weichen in ihrer Temperaturtoleranz nach oben ab, sind somit deutlich besser an den klimatischen Wandel angepasst. Ubiquitärere Arten mit zumeist deutlich größerer ökologischer Amplitude können somit aus dem umgebenden, vielfach landwirtschaftlich genutzten Umland in das Moor einwandern und auch bei zukünftig höheren Temperaturen diese Habitate besiedeln.
- Bezogen auf die Temperaturen werden die Wirkungen des Klimawandels besonders bei den Toleranzen der Arten gegenüber von Jahresmitteltemperatur und Minimaltemperaturen deutlich. Das steht in Übereinstimmung mit den Literaturangaben (vgl.Radchuket al. 2013a, b). Aber auch bei der Maximaltemperatur ist eine leichte bis mäßige Abweichung zwischen den ermittelten Toleranzbereichen der Arten und der Projektionen zu erkennen.
- Beim mittleren Jahresniederschlag stimmen die ermittelten Klimatoleranzen der Arten mit denen der klimatologischen Periode 1950 2000 bzw. Klimaprojektion bis 2070 weitestgehend überein. Da selbst degradierte Moore in Mecklenburg-Vorpommern noch mäßig wassergesättigt sind, kommt dieser Faktor beim Klimawandel wahrscheinlich erst deutlich später zum Tragen.
- Nährstoffarme Moore sind azonale Lebensräume. Das konnte bei der Ermittlung der Toleranzbereiche der Arten nur unvollständig Berücksichtigung finden. Da aber die Temperaturverhältnisse in den Mooren extremer als im Umland sind, müssen die Toleranzen eher enger gesehen werden.
3.2 Einordnung der Klimatoleranzen von Moorarten bezogen auf den Verbreitungstypus
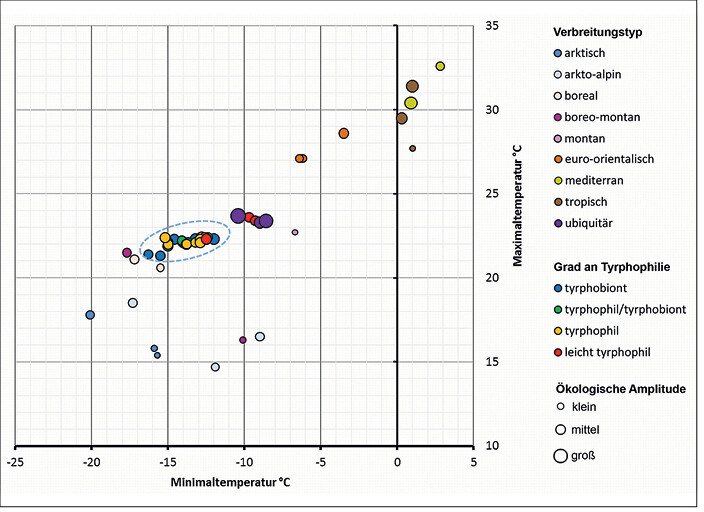
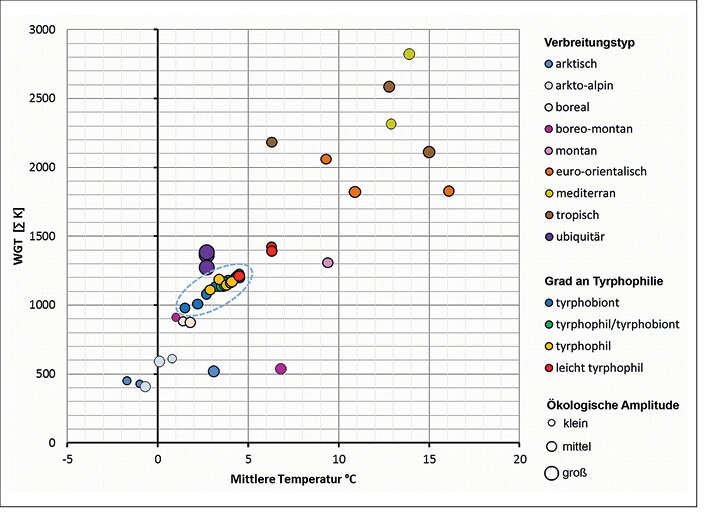
Um zu analysieren, welchem Verbreitungstypus die tyrphobionten/tyrphophilen Arten angehören, wurden die Klimatoleranzen von jeweils drei Arten mit bekanntem Verbreitungstypus (vgl.Kudnraet al. 2015) auf Basis ihres Verbreitungsgebietes errechnet. In diese Reihe sind die Taxa der nährstoffarmen Moore eingeordnet worden. In Abb. 5 und 6 werden die Zentren und Spannweiten der Toleranzbereiche als Indikatoren der ökologischen Klimaoptima und -amplituden für die jeweiligen Arten in Diagrammen dargestellt. Dabei wurden die funktionalen Zusammenhänge bezüglich Minimal- und Maximaltemperatur sowie mittlerer Temperatur und Wachstumsgradtagen abgebildet. Es wird deutlich, dass
- die Zentren der Toleranzbereiche der tyrphobionten und -philen Arten dicht zusammenliegen,
- sie sich zwischen denen der arkto-alpinen/borealen und euro-sibirischen Arten einordnen und
- diese den Toleranzbereichen der ubiquitären Arten nahe liegen (wenn auch größere Spannweiten auftreten). Ubiquisten sind damit ideale" Sekundärbesiedler von Mooren.
4 Beispiel für die Optimierung von Lebensräumen in einem mäßig degradierten Hochmoor zur Schaffung von Zeit für die Klimaanpassung
Die Untersuchungen haben gezeigt, dass tyrphobionte und tyrphophile Lepidopteren sehr ähnliche ökologische Toleranzen bezüglich der betrachteten klimatischen Parameter aufweisen. Von den Anspruchskomplexen an Temperatur, Länge der Vegetationsperiode und Niederschlag her sind sie mit Taxa vergleichbar, die heute in der borealen Zone gefunden werden (boreo-montaner Verbreitungstyp). Sie sind an kühlere und kontinentalere Verhältnisse angepasst. Somit wird offensichtlich, dass nicht nur Strukturen und Ressourcen des Lebensraumes Hochmoor das Vorkommen der Taxa bestimmen, sondern auch ganz entscheidend mesoklimatische Faktoren.
Weiter indizieren die durchgeführten Analysen, dass es notwendig wird, den Arten durch optimale Habitatbedingungen Zeit für die Anpassung zu verschaffen. Dazu soll als praktisches Beispiel die partielle ökologische Sanierung des FND Großes Rahmannsmoor" in Krakow am See dienen. Dieses Kesselmoor liegt in der kuppigen Endmoränenlandschaft und ist vegetationsökologisch durch Torfmoos-Seggen-Wollgrasriede gekennzeichnet. Im Bereich des Laggs stehen Bruch- und Grauweidengebüsche, die mit Birken durchmischt sind. Durch einen gestörten Landschaftswasserhaushalt konnten sich bis in die 1990er-Jahre hinein auf größeren Flächen Kiefern und Birken ausbreiten. Es gab aber immer noch genügend Offenland, auf dem der Hochmoor-Scheckenfalter ( Boloria aquilonaris ) lebte. Der letzte Nachweis gelang 1998, dann verschlechterte sich durch den forcierten Aufwuchs von Gehölzen die Lage dermaßen, dass nur noch wenige baumfreie Flächen existierten. Die Fraßpflanze des Schmetterlings (Moosbeere) gab es weiterhin reichlich, doch das Moor trocknete stärker aus. Ursache dafür war die Kombination von Verdunstung der Gehölze und sich verstärkendem Klimawandel. Im Frühjahr 2016 gelang es, 0,5 ha vornehmlich Kiefernwald zu fällen. Prognostiziert war ein Anstieg des Moorwasserspiegels um 10 20 cm (Abb. 7).
Bei Untersuchungen, die Ende Juni/Anfang Juli durchgeführt wurden, konnte festgestellt werden, dass das Wasser im Moor trotz vorausgegangener Hitzeperiode flurgleich stand. Es wurden gut entwickelte Bestände an Rundblättrigem Sonnentau ( Drosera rotundifolia ), Moosbeere ( Vaccinium oxycoccos ) und Schmalblättrigem Wollgras ( Eriophorum angustifolium ) gefunden. Die Sphagnumdecke ( S. fallax, S. squarrosum, S. palustre ) war vital. Nach fast 20 Jahren konnte auch der tyrphobionte Hochmoor-Scheckenfalter in wenigen Exemplaren auf der Renaturierungsfläche wiedergefunden werden. Es kann davon ausgegangen werden, dass diese Art in kleinen, waldfreien Bereichen das Pessimum überleben konnte. Somit besteht die berechtigte Hoffnung, dass dieser Bestand auf Grund der größeren Flächenverfügbarkeit sich erholt und deutlich individuenstärker wird. Es ist somit Zeit für die Anpassung geschaffen worden.
Dank
Für die sachgerechte Aufarbeitung der Daten zu den Verbreitungsgebieten der Arten und die Berechnung der statistischen Größen sowie ihre Visualisierung sei Carola Höfs und Torsten Foy herzlich gedankt. Die ökologische Sanierung im FND Großes Rahmannsmoor" wurde von der Unteren Naturschutzbehörde des Landkreises Rostock finanziert und begleitet. Dafür sei insbesondere Frank Vökler vielmals gedankt.
Literatur
Aarvik, L., Hansen, L.O., Kononenko, V. (2009):Norges Sommerfugler. Håndbok over Norges dagsommerfugler og nattsvermere. Norsk Entomologisk forening, Naturhistorisk museum, Universitetet i Oslo.
Begon, M.E., Harper, J.L., Townsend, C.R.(1998): Ökologie. Spektrum, Heidelberg, Berlin.
Bräu, M., Bolz, R., Kolbeck, H., Nummer, A., Voith, J., Wolf, W.(2013): Tagfalter in Bayern. Ulmer, Stuttgart.
Bühler-Cortesi, T.(2012): Schmetterlinge. Tagfalter der Schweiz. Haupt, Bern.
Devictor, V., van Swaay, C., Brereton, T., Brotons, L., Chamberlain, D., Heliölä, J., Herrando, S., Julliard, R., Kuussaari, M., Lindström, Å., Reif, J., Roy, D., van Strien, A, Settele, J., Schweiger, O., Stefanescu, C., Vermouzek, Z., van Turnhout, C., Wallis de Vries, M., Wynhoff, I., Jiguet, F.(2012): Differences in the climatic debts of birds and butterflies at a continental scale. Nature Climate Change 2, 121-124.
Ebert, G.(Hrsg., 2003): Die Schmetterlinge Baden-Württembergs. Ulmer, Stuttgart, Band 9: Nachtfalter VII.
Funet(2015): Lepidoptera. www.nic.funet.fi/pub/sci/bio/life/insecta/lepidoptera/ , abgerufen im Mai 2016.
Henriksen, H.J., Kreuzer, I.(1982): The butterflies of Scandinavia in nature. Skandinavisk Bogforlag, Odense.
Hijmans, R.J., Cameron, S.E., Parra, J.L., Jones, P.G., Jarvis, A. (2005): Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology 25, 1965-1978.
Inman-Tjørve, I., Trolle, L.(1999): Sommerfugler i Norge. Aschehougs Naturbøker, Oslo.
Kudnra, O., Pennersdorfer, J., Lux, K.(2015): Distribution atlas of European butterflies and skippers. PEKS, Schwanfeld.
Lafranchis, T.(2004): Butterflies of Europe. New field guide and key. Diatheo, Paris.
Leraut, P.(2006): Moths of Europe. Volume 1. Saturnids, Lasiocampids, Hawkmoths, Tiger Moths. N.A.P Editions, Verrières le Busisson.
Macek, J., Dvor ák, J., Traxler, L.,C ervenka, V.(2007): Motýli a housenky str ední Evropy. Noc ní motýli I. Academia, Praha.
, Dvor ák, J., Traxler, L.,C ervenka, V.(2008): Motýli a housenky str ední Evropy. Noc ní motýli II. Academia, Praha.
, Procházka, J., Traxler, L.(2012): Motýli a housenky str ední Evropy. Noc ní motýli III. Academia, Praha.
Marttila, O., Haahtela, T., Aarnio, H., Ojalainen, P.(1992): Päiväperhos opas. Kirjayhtymä, Helsinki.
Paolucci, P. (2013): Butterflies and Burnets of the Alps and their Larvae, Pupae and Cocoons. WBA Handbooks 4, Verona.
Radchuk, V., Turlure, C., Schtickzelle, N.(2013a): Predicting responses to climatic change requires all life-history stages. J. Anim. Ecol. 82, 3 - 5.
, Turlure, C., Schtickzelle, N.(2013b): Each life stage matters: the importance of assessing the response to climatic change over the complete life cycle in butterflies. J. Anim. Ecol. 82, 275-285.
Saarinen, K., Jantunen, J.(2003): Perhoset 2. Päivällä lentävät yön perhoset. Werner Söderström Osakeyhtiö, Helsinki.
Settele, J., Kudnra, O., Harpke, A., Kühn, I., van Swaay, C., Verovnik, R., Warren, M., Wiemers, M., Hanspach, J., Hickler, T., Kühn, E., van Halder, I., Veling, K., Vliegenthart, A., Wynhoff, I., Schweiger, O.(2008): Climatic Risk Atlas of European Butterflies. Pensoft, Sofia, Moscow.
Skou, P. (1991):Nordens Ugler. Danmarks Dyreliv. Apollo Books, Stenstrup.
Sommer, R., Thiele, V., Seppä, H.(2015): Use and misuse of the term "glacial relics in the Central European biogeography and Conservation ecology of insects. Insect Conservation and Diversity, DOI: 10.1111/icad.12109.
Thiele, V., Luttmann, A.(2015): Tyrphobionte Schmetterlingsarten nährstoffarmer Moore. Eine parametergestützte Analyse zum Artenspektrum als Grundlage für Schutzstrategien mit Hinblick auf den Klimawandel. Naturschutz und Landschaftsplanung 47 (4), 101-108.
, Luttmann, A., Hoffmann, T., Röper, C. (2014a):Ökologische Auswirkungen von Klimaänderungen und Maßnahmenstrategien für europäisch geschützte Arten. Fallstudie zweier FFH-Gebiete in Sachsen-Anhalt. Naturschutz und Landschaftsplanung 46 (6), 169-176.
, Luttmann, A., Hoffmann, T., Schuhmacher, S., Blumrich, B.(2016): Zur Bestandsdynamik tyrphobionter/-philer Schmetterlinge in nährstoffarmen Mooren Mecklenburg-Vorpommerns über die letzten 125 Jahre und anthropogen wie klimatisch bedingte Ursachen der Bestandsschwankungen. Naturschutz und Landschaftsplanung 48 (7), 227-233.
, Luttmann, A., Liebe Edle von Kreutzner, K., Degen, B., Berlin, A., Lipinski, A., Niederstraßer, J., Koch, R., von dem Bussche, J. (2013):Durchführung einer Untersuchung zu den Folgen des Klimawandels in Sachsen-Anhalt. Wirkungen des Klimawandels auf europäisch geschützte Arten und Lebensräume. Berichte des Landesamtes für Umweltschutz Sachsen-Anhalt 6.
Trolle, L., Rubæk, B.(2003): Gads Håndbog om Sommerfugle. Gad & Natur, København.
Van Dyck, H., Bonte, D., Puls, R., Gotthard, K., Maes, D. (2015):The lost generation hypothesis: could climate change drive ectotherms into a developmental trap? Oikos 124 (1), 54-61.
Fazit für die Praxis
- Tyrphobionte/-phile Schmetterlingsarten der nährstoffarmen Moore gehören dem boreo-montanen Verbreitungstyp an und haben eine relativ einheitliche Temperatur- und Niederschlagstoleranz. In Mecklenburg-Vorpommern wird die Toleranz der Arten bezüglich der Jahresmittel- und Minimaltemperaturen bereits heute vielfach überschritten.
- Somit ist das langfristige Überleben der stenotopen Arten in Frage gestellt. Um den Arten die Chance zu geben, sich an den Klimawandel anzupassen, müssen die Habitatverhältnisse in vielen Mooren verbessert werden:
- Über die Fällung von Bäumen kann deutlich die Wasserbilanz optimiert werden (Senkung der Verdunstung); wichtig ist das Ringeln bei Birkenaufwuchs. Zudem sind zahlreiche Moorfalter schattenfliehend.
- Dem Erfahrungsaustausch zwischen den regionstragenden Akteuren kommt besondere Bedeutung zu, da es zu wenige Erkenntnisse in diesem Zusammenhang gibt.
Kontakt
Dr. Volker Thiele ist Geschäftsführer des Institutes biota Institut für ökologische Forschung und Planung GmbH in Bützow und arbeitet vornehmlich im Bereich des Gewässer- und Moorschutzes. Forschung zur Bioindikation, zur Entwicklung ökologischer Bewertungsverfahren und zu Klimafolgen. Planung für Natur- und Artenschutz. Spezialist für Lepidopteren und Autor zahlreicher wissenschaftlicher Artikel und Bücher. Von der IHK Rostock öffentlich als Sachverständiger für Naturschutz und Landschaftspflege sowie Gewässerschutz bestellt und vereidigt.
> volker.thiele@institut-biota.de
Dr. Tim Hoffmann arbeitet seit 2009 als Arbeitsgruppenleiter im Institut biota überwiegend in den Fachbereichen Geoinformatik, Statistik und Hydrologie. Promotion 2005 an der Universität Greifswald im Fach Geoinformatik. Forschungen zu anthropogenen Eingriffen in den Wasserhaushalt, Klimawandel und seine Auswirkungen auf das Hochwassergeschehen sowie räumliche Datenanalysen. Mitarbeiter in mehreren Bundesforschungsprojekten zum Thema regionales Wasserressourcen-Management.
-
Abb. 1: Toleranzbereiche von Klimaindikatoren für tyrphobionte und tyrphophile Arten bezüglich Minimaltemperatur und Angabe des mittleren Werts für die Periode 1950 – 2000 sowie für die Klimaprojektion des Jahres 2070 (rcp85) in Mecklenburg-Vorpommern (Legende: Median und Streuung der Werte, tp = tyrphophil, tb = tyrphobiont, rote Linie: Abgrenzung leicht tyrphophiler Arten). Tolerance ranges of climate indicators of tyrphobiontic and tyrphophilic species regarding minimum temperature and average values for the period 1950 - 2000 and the climate projections for the year 2070 (rcp85) in Mecklenburg-Western Pomerania (Key: median and value dispersal, tp = tyrphophilic, tb = tyrphobiontic, red line: delimitation of weak tyrphophilic species).Von Volker Thiele und Tim Hoffmann -
Abb. 2: Toleranzbereiche von Klimaindikatoren für tyrphobionte und tyrphophile Arten bezüglich der Jahresmitteltemperatur und Angabe des mittleren Werts für die Periode 1950 – 2000 sowie für die Klimaprojektion des Jahres 2070 (rcp85) in Mecklenburg-Vorpommern (Legende: Median und Streuung der Werte, tp = tyrphophil, tb = tyrphobiont, rote Linie: Abgrenzung leicht tyrphophiler Arten). Tolerance ranges of climate indicators of tyrphobiontic and tyrphophilic species regarding relative annual mean temperature and average values for the period 1950 - 2000 and the climate projections for the year 2070 (rcp85) in Mecklenburg-Western Pomerania (Key: median and value dispersal, tp = tyrphophilic, tb = tyrphobiontic, red line: delimitation of weak tyrphophilic species).Von Volker Thiele und Tim Hoffmann -
Abb. 3: Toleranzbereiche von Klimaindikatoren für tyrphobionte und tyrphophile Arten bezüglich der Maximaltemperatur und Angabe des mittleren Werts für die Periode 1950 – 2000 sowie für die Klimaprojektion des Jahres 2070 (rcp85) in Mecklenburg-Vorpommern (Legende: Median und Streuung der Werte, tp = tyrphophil, tb = tyrphobiont, rote Linie: Abgrenzung leicht tyrphophiler Arten). Tolerance ranges of climate indicators of tyrphobiontic and tyrphophilic species regarding maximum temperature and average values for the period 1950 - 2000 and the climate projections for the year 2070 (rcp85) in Mecklenburg-Western Pomerania (Key: median and value dispersal, tp = tyrphophilic, tb = tyrphobiontic, red line: delimitation of weak tyrphophilic species).Von Volker Thiele und Tim Hoffmann -
Abb. 4: Toleranzbereiche von Klimaindikatoren für tyrphobionte und tyrphophile Arten bezüglich des Jahresniederschlages und Angabe des mittleren Werts für die Periode 1950 – 2000 sowie für die Klimaprojektion des Jahres 2070 (rcp85) in Mecklenburg-Vorpommern (Legende: Median und Streuung der Werte, tp = tyrphophil, tb = tyrphobiont, rote Linie: Abgrenzung leicht tyrphophiler Arten). Tolerance ranges of climate indicators of tyrphobiontic and tyrphophilic species regarding annual precipitation and average values for the period 1950 - 2000 and the climate projections for the year 2070 (rcp85) in Mecklenburg-Western Pomerania (Key: median and value dispersal, tp = tyrphophilic, tb = tyrphobiontic, red line: delimitation of weak tyrphophilic species).Von Volker Thiele und Tim Hoffmann -
Abb. 5: Auf Basis der Verbreitungsgebiete ermittelte Temperaturtoleranzen des wärmsten und kältesten Monats von tyrphobionten, tyrphophilen und ausgewählten Vergleichsarten verschiedener Verbreitungstypen (blau umgrenzt: Parameterbereich der tyrphobionten bis tyrphophilen Arten); der Durchmesser der Kreise symbolisiert relativ zueinander die Größe ihrer ökologischen Amplitude. Temperature range of the warmest and coldest month of tyrphobiontic, tyrphophilic and selected comparative species of different distribution types, based on distribution areas (encircled area: parameter area of tyrphobiontic to tyrphophilic species); the diameter of the circles symbolizes their ecological amplitude relative to each other.Von Volker Thiele und Tim Hoffmann -
Abb. 6: Auf Basis der Verbreitungsgebiete ermittelte Toleranzen bezüglich der mittleren Temperaturen und Wachstumsgradtage (WGT) von tyrphobionten, tyrphophilen und ausgewählter Vergleichsarten (blau umgrenzt: Parameterbereich der tyrphobionten bis leicht tyrphophilen Arten); der Durchmesser der Kreise symbolisiert relativ zueinander die Größe ihrer ökologischen Amplitude. Tolerances with respect to the range of mean temperatures and days of growth degree (WGT) of tyrphobiontic, tyrphophilic and selected comparative species, calculated on the basis of the distribution areas (encircled area: parameter area of tyrphobiontic to slightly tyrphophilic species); the diameter of the circles symbolizes the size of their ecological amplitude relative to each other.Von Volker Thiele und Tim Hoffmann -
Abb. 7: Restaurierte Fläche im Großen Rahmannsmoor" (großes Bild) und typische Arten (Moosbeere, Hochmoorscheckenfalter, Wollgrasbulten kleine Bilder). Restored area in the bog Großes Rahmannsmoor" (large picture) and typical species (small pictures: Cranberry, Cranberry Fritillary, mounts of Cotton Grass).Volker Thiele, Tim Hoffmann -
-
-
Von Volker Thiele und Tim Hoffmann -
Von Volker Thiele und Tim Hoffmann

Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.