Erhaltungskulturen zur Wiederansiedlung des Karpaten-Enzians im Osterzgebirge
Abstracts
Um das einzige deutsche Vorkommen des Karpaten-Enzians (Gentianella lutescens (Velen.) Holub) am Geisingberg im Osterzgebirge zu erhalten, wurden zwischen 2010 und 2015 im Rahmen der Richtlinie Natürliches Erbe des Freistaats Sachsen Projekte zur Stützung der Population durchgeführt. Es wurden Erhaltungskulturen zunächst in den botanischen Gärten Altenberg-Schellerhau und Marburg aufgebaut. Auf vorbereiteten Flächen in der Nähe des einzigen verbliebenen Alt-Standortes und an weiteren ehemaligen Standorten in der Umgebung wurden insgesamt 42 300 überwiegend in den Erhaltungskulturen geerntete Samen und 159 angezogene Jungpflanzen ausgebracht.
Trotz schwieriger Bedingungen (u.a. zu trockene Sommer, Schädlinge) und erheblichem Mehraufwand (Gießen während Dürrephasen) wurden bis Juli 2015 auf sechs Flächen Teilpopulationen mit insgesamt 175 einjährigen und 464 blühenden Pflanzen wieder oder neu begründet. Auch der Alt-Bestand hat sich positiv entwickelt: In der Projektlaufzeit ist dieser von 191 blühenden Pflanzen in 2009 und vier in 2010 auf 1660 blühende Pflanzen in 2015 gewachsen. Dies hängt vermutlich mit einer späten Nachmahd bzw. -beweidung zusammen. Der Nutzen von Erhaltungskulturen und Wiederansiedlungen als Maßnahmen eines technischen Artenschutzes werden vor dem Hintergrund der Auswirkungen großklimatischer Veränderungen auf Lebensräume diskutiert.
Cultivation of Gentianella lutescens for its re-establishment in the Osterzgebirge, Saxony Successful combination of ex-situ and in-situ conservation measures
Between 2010 and 2015 several management projects funded by the Saxon state guideline Richtlinie Natürliches Erbe were carried out to save the only population of Gentianella lutescens in Germany in the Osterzgebirge (Eastern Ore Mountains) near the town of Altenberg (Saxonia). Ex-situ conservation cultures were established in the botanical gardens of Altenberg-Schellerhau and Marburg. Close to the last remaining occurrence of the species and on further former areas of its dispersal in the surrounding several sites were prepared, and altogether 159 juvenile plants and 42.300 seeds harvested predominantly from the ex-situ cultures were deployed.
In spite of unfavourable conditions (such as dry summers, pests) and considerable additional efforts (watering during drought) stands could be established on six sites with altogether 175 juvenile plants and 464 adult flowering plants until July 2015. Even the initial population increased from 191 flowering individuals in 2009 and only 4 in 2010 to 1.660 flowering plants in 2015. This has probably been caused by a late date of after-mowing or sheep-grazing. The benefit of ex-situ conservation cultures and re-establishment as highly technical measures of species conservation is discussed against the background of macroclimatic changes of their habitats.
- Veröffentlicht am

Carpathian gentian (Gentianella lutescens).Randolf Manderbach
1 Einleitung
Der Karpaten-Enzian (Gentianella lutescens) (Abb. 1) besitzt nur noch ein Vorkommen in Deutschland am Geisingberg bei Altenberg (Sachsen) (Abb. 2). Die Art ist damit in Sachsen und Deutschland vom Aussterben bedroht. Die nächstgelegenen aktuellen Vorkommen in Europa liegen im Biosphärenreservat Bile Karpaty (Weiße Karpaten) in Südost-Tschechien, ca. 500km Luftlinie vom Vorkommen am Geisingberg entfernt. Weitere finden sich in Kärnten, der Steiermark und den slowenischen Alpen (Aeschimann et al. 2004, Amt der Steiermärkischen Landesregierung 2007).
Trotz intensiver Schutzbemühungen unterlag die Population im NSG Geisingberg über viele Jahre starken Bestandsschwankungen. Nach einem Tiefstand Anfang der 1990er-Jahre erhöhten sich die Bestandszahlen in den Folgejahren (Uhlig & Müller 2001), um dann bis 2003 erneut stark abzunehmen. Ab 2005 schwankte der Bestand der Art in einem zweijährigen Zyklus extrem, in geraden Jahren sank die Anzahl blühender Pflanzen auf 6 in 2008 und 4 in 2010.
Dieser dramatischen Entwicklung sollte ein erstes Projekt ab Mai 2010, das im Rahmen der damals neu aufgelegten Richtlinie Natürliches Erbe des Freistaats Sachsen gefördert wurde, durch Aufbau von Ex-situ-Erhaltungskulturen und (Wieder-)Ansiedlungsmaßnahmen entgegenwirken. Ex-situ-Erhaltungskulturen als Basis für (Wieder-)Ansiedlungsmaßnahmen sind mittlerweile ein erprobtes Mittel, um Populationen vor allem hochgradig gefährdeter Arten, für deren Schutz Deutschland beispielsweise eine besondere Verantwortung hat oder die auf den Anhängen II oder IV der FFH-Richtlinie stehen, in situ zu stützen oder zu re-etablieren oder ex situ für die Zukunft zu sichern. Eine sehr gute Übersicht liefern hierzu Diekmann et al. (2015).
Das dieser Studie zugrunde liegende Projekt zum Karpaten-Enzian ist in Deutschland hinsichtlich der hohen Gefährdungskategorie der Art, des relativ hohen Aufwands und der Langfristigkeit der Maßnahmen sowie der wissenschaftlichen Dokumentation mit Ansiedlungsmaßnahmen z.B. bei der Finger-Küchenschelle (Pulsatilla patens) in der Garchinger Heide in Bayern (Röder & Kiehl 2008), dem Stengellosen Tragant (Astragalus excapus) in Thüringen (Becker 2012), dem Böhmischen Enzian (Gentianella bohemica) in Bayern (Dolek et al. 2009, Koeniger 2012, Zillig et al. 2010) und der Hirschzunge (Asplenium scolopendrium) (Becker & Becker 2010) zu vergleichen.
Erhaltungskulturen gefährdeter Arten bedürfen häufig einer aufwendigen Kulturführung (vgl. Brunzel 2010, Lauterbach et al. 2015, Zillig et al. 2010). Insbesondere Keimung und Jungpflanzenaufzucht sind bei Vertretern der zweijährigen Gattung Gentianella häufig schwierig, was u.a. an der zur Keimung benötigten Kältestratifikation, der Empfindlichkeit insbesondere der Jungpflanzen gegenüber Hitze und Trockenheit sowie einer häufig notwendigen Infizierung mit vesikulär-arbuskulärer Mykorrhiza (VAM) liegt (Brabec 2005, Brunzel 2010, Dolek 2008, Deno 1996, Grzes 2007, Milberg 1994, Zillig et al. 2010). So war es auch bei Gentianella lutescens bis 2007 trotz intensiver Bemühungen nicht gelungen, Samen in nennenswertem Umfang zur Keimung zu bringen und so eine funktionierende Ex-situ-Erhaltungskultur aufzubauen (Müller & Zoephel 2008). Erst in den Jahren 2007 bis 2009 gelang es im Rahmen eines von der Bundesanstalt für Landwirtschaft und Ernährung (BLE) geförderten Projekts im Botanischen Garten der Universität Marburg, eine funktionierende Kultur der Art zu etablieren (Brunzel 2010). Durch diese Ex-situ-Erhaltungskultur konnte erstmals ausreichend Saatgut produziert werden, um die dieser Studie zu Grunde liegenden Projekte starten zu können. Ziel dieser vom Freistaat Sachsen geförderten Projekte war es, ausgehend von einer aufzubauenden Ex-situ-Erhaltungskultur vor allem im Botanischen Garten Schellerhau bei Altenberg, mit der Produktion von Samen und Jungpflanzen Wiederansiedlungsmaßnahmen auf Flächen in der Nähe des einzig verbliebenen Alt-Standortes auf der sog. Hufeisenwiese am Geisingberg sowie auf ehemals besiedelten Flächen in der Umgebung Altenbergs zu ermöglichen.
2 Methoden
Zunächst wurde Saatgut vom einzig verbliebenen, ursprünglichen Standort auf der Hufeisenwiese am Geisingberg gesammelt, um Ex-situ-Erhaltungskulturen aufzubauen. Diese bestanden zur Risikominimierung von 2010 bis 2012 zunächst im Botanischen Garten Marburg und im Botanischen Garten Schellerhau bei Altenberg. Von 2012 bis 2015 befanden sich zwei Erhaltungskulturen im Botanischen Garten Schellerhau und in einem Privatgarten in Naundorf, ebenfalls im Osterzgebirge. In den Ex-situ-Kulturen wurden ebenso wie in den Teilpopulationen in situ regelmäßig verschiedene Fitness-Parameter der Pflanzen (Blütenzahl, Durchmesser der Grundblattrosette, Länge größtes Laubblatt, Anzahl Samenkörner je Frucht) protokolliert. Das während der Projektlaufzeit geerntete Saatgut sowie die herangezogenen Jungpflanzen wurden benutzt, um sowohl die Erhaltungskulturen weiter auszubauen als auch jedes Jahr die neu begründeten Teilpopulationen in situ aufzufrischen bzw. die Chance der dauerhaften Etablierung durch ausreichend hohe Individuenzahlen zu erhöhen.
Die Ex-situ-Kulturen mussten mit Schneckenzäunen geschützt werden (Abb. 3). Während der Projektlaufzeit mussten die Kulturen einmal gegen Pilzbefall behandelt werden (mit dem Fungizid Ortiva®), einmal im Jahr wurde Schneckenkorn innerhalb des Schneckenzauns gestreut und zusätzlich alle Töpfe wöchentlich auf Schnecken kontrolliert. Gegen den Befall des Kleinschmetterlings Stenoptilia nelorum Gibeaux, dessen Larve die Fruchthülle durchbohrt und die Samenanlagen auffrisst, wurde jede Saison in dem Zeitraum des Erscheinens von großen Knospen bis zum Ausreifen der Samen dreimal ein Insektizid gesprüht.
Das Substrat in den Aussaattöpfen bestand im Wesentlichen aus Drainageschicht, Sand und nährstoffarmen Spezial-Pflanzerden (Lignostrat®, Neudohum®), dem Pikiersubstrat wurde dann zumeist noch Mykorrhiza-Substrat (Mykotown®) zugesetzt. Im Detail sind die Substrate bei Brunzel (2010) und Zillig et al. (2010) beschrieben.
Zur Etablierung von neuen Beständen, im Folgenden Teilpopulationen genannt, wurden Flächen so vorbereitet, dass kleinflächige Bodenverwundungen und Vegetationslücken in einer Größe von ca. 10 x 10 cm entstanden. Die Deckung der vegetationsfreien Lücken sollte insgesamt 25 % der bearbeiteten Fläche nicht überschreiten. Diese Vegetationslücken dienen als Nischen für Keimung und Etablierung von Jungpflanzen, was in einer geschlossenen Vegetationsdecke ansonsten nur ausnahmsweise möglich ist. Größere Bodenverwundungen sind ungeeignet, weil die Gefahr der Austrocknung von Jungpflanzen zu groß ist (vgl. Brabec 2010, Dolek et al. 2009). Es wurden mehrere Verfahren getestet (u.a. maschineller Vertikutierer, Handvertikutierer, manuelles Abharken mit schwerem Eisenrechen). Mahd mit anschließendem zweimaligem tiefgreifendem Abrechen mit einem Eisenrechen erbrachte die besten Ergebnisse (Deckung der vegetationsfreien Flächen von im Mittel 14 % ), weshalb vornehmlich dieses Verfahren im Weiteren zur Vorbereitung der Aussaatflächen genutzt wurde (vgl. auch Brabec 2010, Dolek et al. 2009). In den Projektjahren 2012 bis 2014 wurden die Aussaaten und Auspflanzungen in manuell angelegte Vegetationslücken von 10 x 10 cm vorgenommen.
Zur Neu- bzw. Wiederbegründung von Teilpopulationen wurden zunächst Flächen ausgewählt, die in enger Nachbarschaft zu dem Alt-Standort lagen und augenscheinlich vergleichbare Bedingungen aufwiesen. Dann wurden Flächen in der weiteren Umgebung des Geisingbergs (max. 10km Umkreis) gewählt, die ebenfalls geeignet erschienen und/oder auf denen G. lutescens früher vorkam.
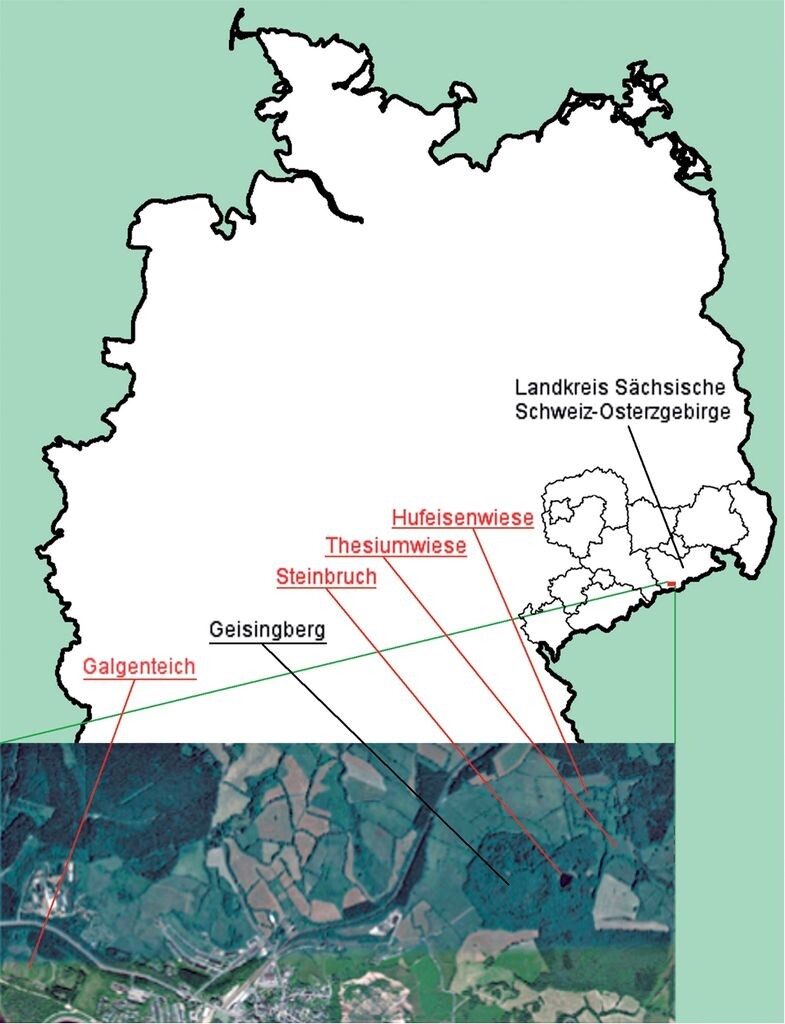
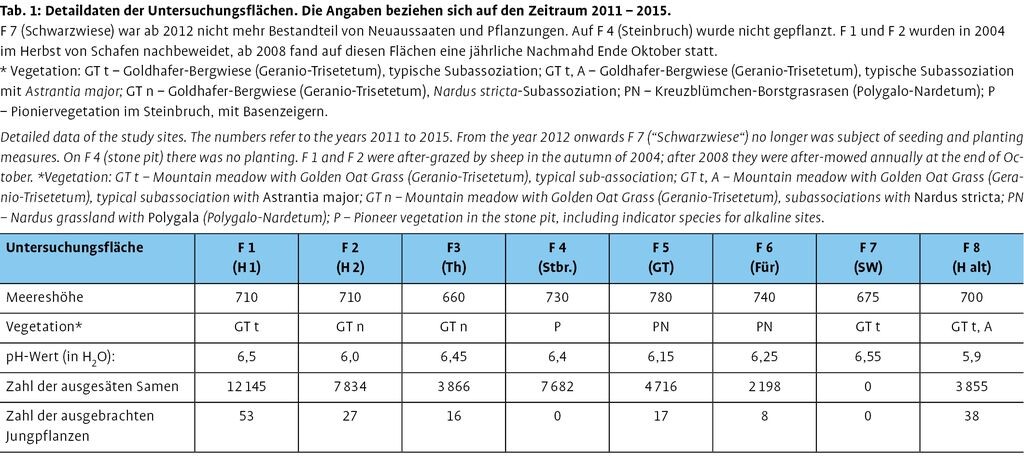
Hinsichtlich der Vegetation der früheren Standorte und des verbliebenen Alt-Standorts erschienen für die insgesamt acht Untersuchungsflächen (F 1: Hufeisenwiese 1, F 2: Hufeisenwiese 2, F 3: Thesiumwiese, F 4: Steinbruch, F 5: Galgenteich, F 6: Fürstenau, F 7: Schwarz´ Wiese, F 8: Hufeisenweise alt) insbesondere Übergänge zwischen der typischen Goldhafer-Bergwiese und deren Borstgras-Subassoziation sowie den Kreuzblümchen-Borstgrasrasen mit einer lockeren Vegetationsdeckung auf leicht basenreichem Substrat am besten geeignet (Tab. 1). Am Geisingberg sind dies besonders vom Basalt beeinflusste Standorte. Aufgrund der Konkurrenzschwäche der Art wurden Rohbodenstandorte wie im Steinbruch Geisingberg sowie am Galgenteich Altenberg in die Auswahl mit einbezogen.
Insgesamt wurden auf den acht Flächen in einer Größe von 1200 m2 Ausbringungsflächen wie oben beschrieben vorbereitet. Zum Zweck des Jungpflanzen-Monitorings wurde jeweils ein 1-m2-Raster darüber gelegt und die gerasterten Ausbringungsflächen zum Schutz gegen Wildschweine gezäunt (Abb. 4). In allen Ausbringungsflächen wurden zur Beurteilung ihrer Eignung die pH-Werte gemessen (vgl. Tab. 1). Diese lagen damit alle im Bereich des Alt-Standortes (pH≈6,0).
In die vorbereiteten Ausbringungsflächen wurden 2012 bis 2014 insgesamt 42300 Samenkörner in Bodenverwundungen ausgebracht sowie 159 Jungpflanzen ausgepflanzt (vgl. Tab. 1). Bedingt durch die zwei Projektphasen 20102012 und 20122015 sowie die räumlichen Gegebenheiten der Flächen waren auch die eigentlichen Ausbringungsflächen unterschiedlich groß. Deshalb schwankte die Anzahl der ausgebrachten Samen auf den Teilflächen zwischen 2198 und 12145 Korn, die der ausgepflanzten Jungpflanzen zwischen 8 und 53. Die ausgebrachten Jungpflanzen stammten alle, das Saatgut bis inklusive 2013 aus den Ex-situ-Kulturen. 2014 wurden 9566 Samenkörner von den neu etablierten Pflanzen geerntet und in die direkt um die Pflanzen herum vorbereiteten Ausbringungsflächen ausgesät. Ausgesät wurde im Oktober, zum Teil und je nach Witterung auch nochmals im darauffolgenden April.
Jungpflanzen wurden überwiegend im Frühling (Ende Mai/Anfang Juni) ausgepflanzt, anfänglich einmal auch im Herbst (Oktober) mit schlechten Überlebensraten. Zur Aussaat wurde ein Salzstreuer umgebaut und das Saatgut mit Sand in ähnlicher Korngröße vermischt, um Verklumpungen bei der Aussaat zu verhindern. Das ausgebrachte Saatgut wurde auf der Erdoberfläche leicht angedrückt, dann mit gesiebter Erde abgedeckt (Abb. 5) und danach mit einem Blumensprüher befeuchtet.
Frisch gekeimte und ausgepflanzte Jungpflanzen in den neu begründeten Teilpopulationen mussten anfänglich zum Teil aufwändig weiter gepflegt oder gegen unterschiedliche Störungen geschützt werden:
Abdeckung gegen Starkregen und Hagel,
intensives Gießen vor allem in den trockenen Frühsommern 2014 und 2015,
manuelles Entfernen von Moos- und Pflanzenkonkurrenz,
künstliche Bedeckung mit Schnee während starker Barfröste in schneearmen Spätwintern.
Das Projekt war sehr arbeitsaufwändig und kann auch nicht als komplett abgeschlossen betrachtet werden, da noch nicht sichergestellt ist, dass die wieder angesiedelten Bestände auf den Ausbringungsflächen mittelfristig stabil sind. Über beide Projektphasen von Mai 2010 bis August 2015 kann der Zeitaufwand auf ca. 3800 Stunden geschätzt werden.
Bei den statistischen Analysen wurden bei den Fitnessparametern der Bestände auf den unterschiedlichen Flächen und in den Erhaltungskulturen Gruppenvergleiche mit einer Kruskall-Wallis-ANOVA durchgeführt, zur Abschätzung der Populationsentwicklung (Anzahl blühender Individuen) wurden lineare Regressionsmodelle genutzt.
3 Ergebnisse
3.1 Bestände in den Erhaltungskulturen und Größenschwankungen der Pflanzen
Über die gesamte Projektlaufzeit wurden in den Erhaltungskulturen ca. 50000 Korn Saatgut gewonnen. Davon wurden ca. 42300 zur Wieder- bzw. Neuansiedlung von Teilpopulationen auf den ausgewählten Flächen ausgesät, der Rest wurde in andere Projekte abgegeben oder für die jedes Jahr erneut notwendige Aussaat in den Ex-situ-Erhaltungskulturen verwendet.
Über die gesamte Projektlaufzeit wurden in den Ex-situ-Erhaltungskulturen insgesamt über 1000 einjährige Jungpflanzen kultiviert, von denen 159 auf den Flächen ausgepflanzt wurden. Von den in den Kulturen verbliebenen Jungpflanzen wurden insgesamt 571 zur Blüte gebracht und beerntet. Von allen Pflanzen wurden bis zur Samenreife zu verschiedenen Zeiten Fitnessparameter aufgezeichnet, um die Kulturführung zu evaluieren und ggf. zu optimieren (vgl. Abschnitt 2).
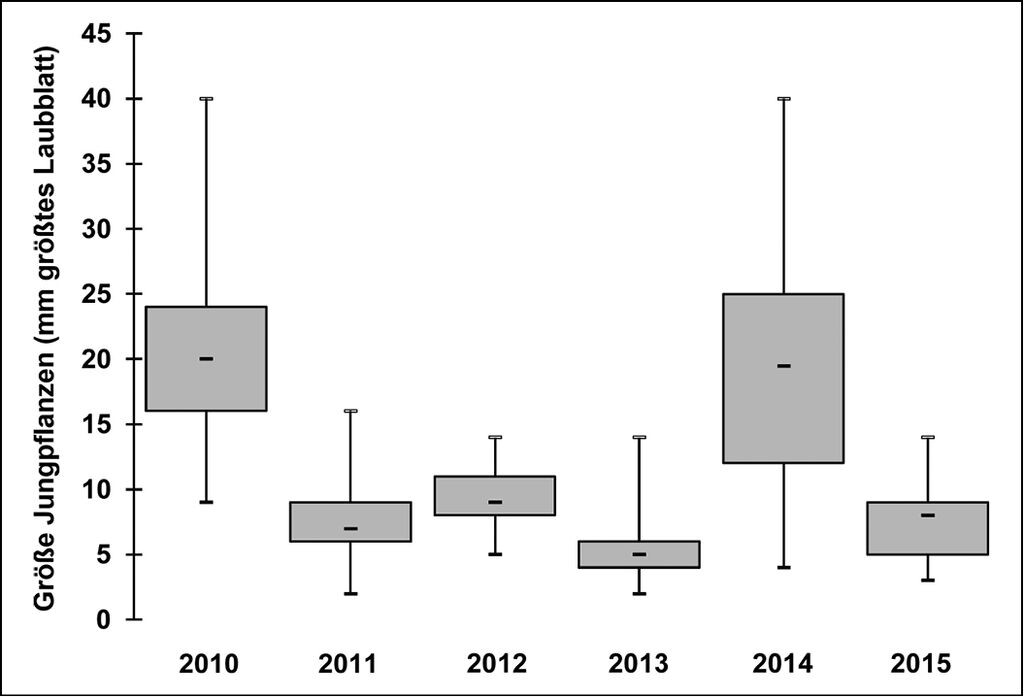
Abb. 6 zeigt die starken Schwankungen der Größen des Fitnessparameters Längstes Laubblatt und damit auch der Pflanzen in den Erhaltungskulturen über die Jahre. So ergab z.B. die Vermessung der Jungpflanzen 2015 für das längste Blatt eine Länge von 7mm im Median, 14mm im Maximum und 2mm im Minimum. Damit waren die Jungpflanzen im Botanischen Garten Schellerhau Ende August 2014 im Mittel dreimal so groß wie 2015 (vgl. Abb. 6). Gemessen wurde aus Gründen der Vergleichbarkeit der Werte jeweils zwischen 31.08. und 08.10. Zu beachten ist, dass die Einjährigen 2010 aus der Erhaltungskultur Schmiedeberg-Naundorf in die Erhaltungskultur im Botanischen Garten Schellerhau überführt wurden, d.h. den größten Teil ihres Wachstums an einem anderen Standort zugebracht haben.
3.2 Bestandsentwicklung ein- und zweijähriger (blühender) Pflanzen auf den Teilflächen 1 bis 8 ab 2013
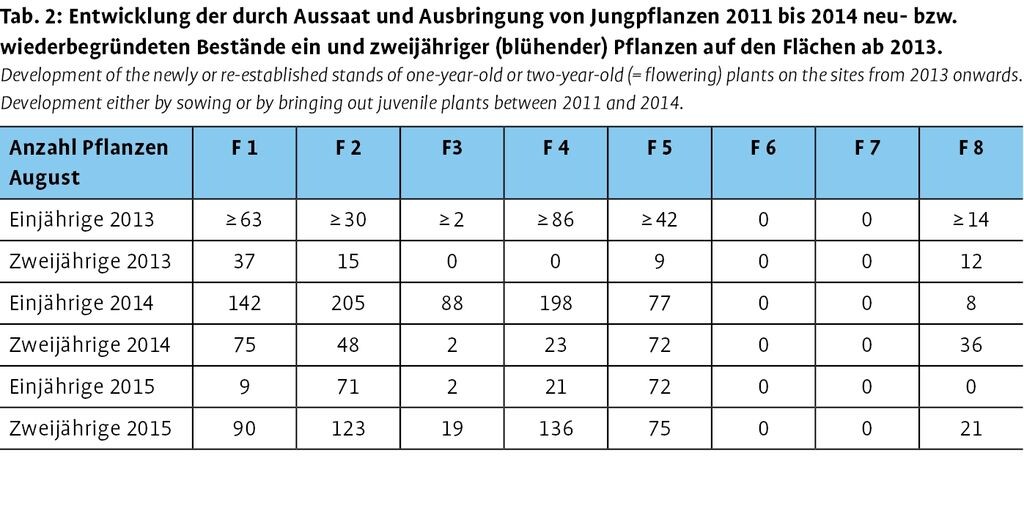
Tab. 2 zeigt die Anzahl der vorhandenen einjährigen Jungpflanzen und zweijährigen blühenden Pflanzen in den letzten drei Projektjahren 2013 bis 2015 sowie die Entwicklung der Bestände in diesem Zeitraum. Die Gesamtzahl aller Pflanzen geht auf Aussaat von 42300 Samenkörnern und Auspflanzung von 159 einjährigen Jungpflanzen über den gesamten Projektzeitraum zurück. Auf den Flächen F 6 und F 7 konnten in keinem Jahr gekeimte Jungpflanzen nachgewiesen werden, ausgepflanzte Jungpflanzen konnten beim Monitoring nach relativ kurzer Zeit auf beiden Flächen ebenfalls nicht mehr festgestellt werden. Zum Teil waren diese abgestorben, zum Teil einfach verschwunden. Unterschiedliche Zahlen der auf den einzelnen Flächen vorhandenen Pflanzen können natürlich prinzipiell auf die unterschiedlichen Mengen ausgebrachter Samen und Jungpflanzen zurückgeführt werden. Dass auf zwei Flächen (TF 6 und 7) über drei Jahre kein einziger Keimling festgestellt wurde, ist damit jedoch nicht zu erklären und dürfte andere Ursachen haben.
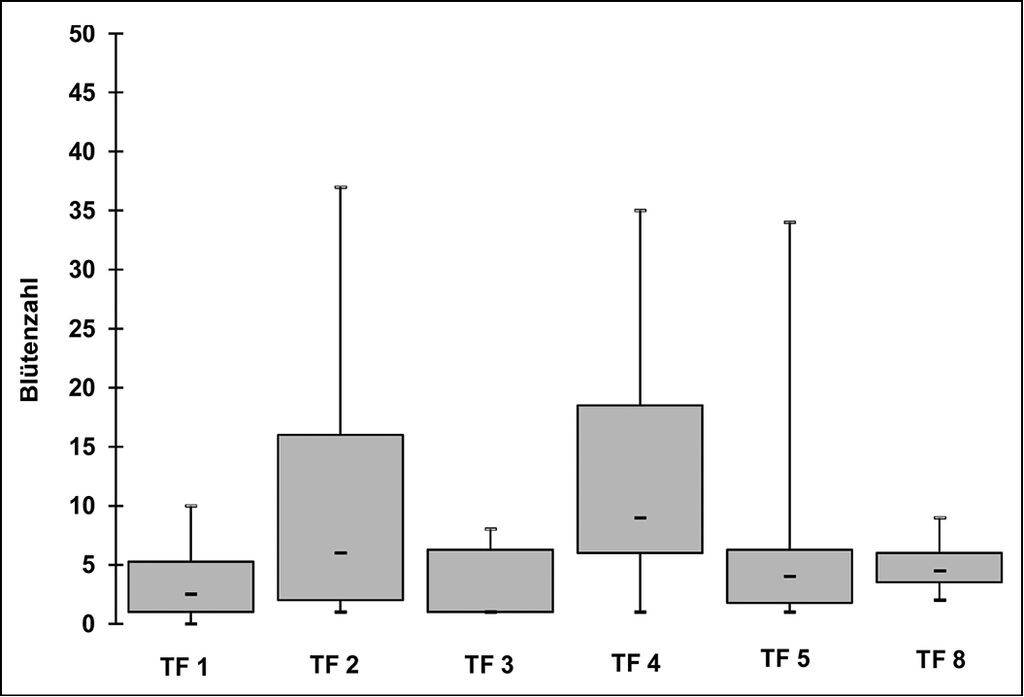
Um Unterschiede in der Eignung der Teilflächen unabhängig von der Anzahl ausgebrachter Samen und Jungpflanzen festzustellen, wurde als Indikator für die Fitness der Pflanzen die Anzahl der Blüten gezählt. Die Blütenzahl korreliert mit dem Durchmesser der Blattrosette und indiziert damit die Größe der Pflanzen (Brunzel 2010). Für den Fitnessparameter kann daher angenommen werden, dass er die Eignung der jeweiligen Teilfläche für die neu oder wieder begründete Teilpopulation abbildet. Abb. 7 zeigt die Anzahl der Blüten in den unterschiedlichen neu bzw. wieder begründeten Teilpopulationen. Die Flächen F 1 (Hufeisenwiese 1), F 2 (Hufeisenwiese 2) und F 8 (Hufeisenwiese alt) wiesen mit 45, 25 und 29 die maximalen Blütenzahlen auf. Hinsichtlich der Blütenzahlen bestehen signifikante Unterschiede (p=0.045) zu F 4 (Steinbruch) und F 1 (Hufeisenwiese 1). Dagegen waren 2014 noch signifikante Unterschiede zwischen mehreren, auch anderen Flächen festzustellen, so dass von einem Angleichen der Flächen hinsichtlich ihrer Blütenzahlen ausgegangen werden kann.
3.3 Populationsdynamik der ursprünglichen Population am Alt-Standort
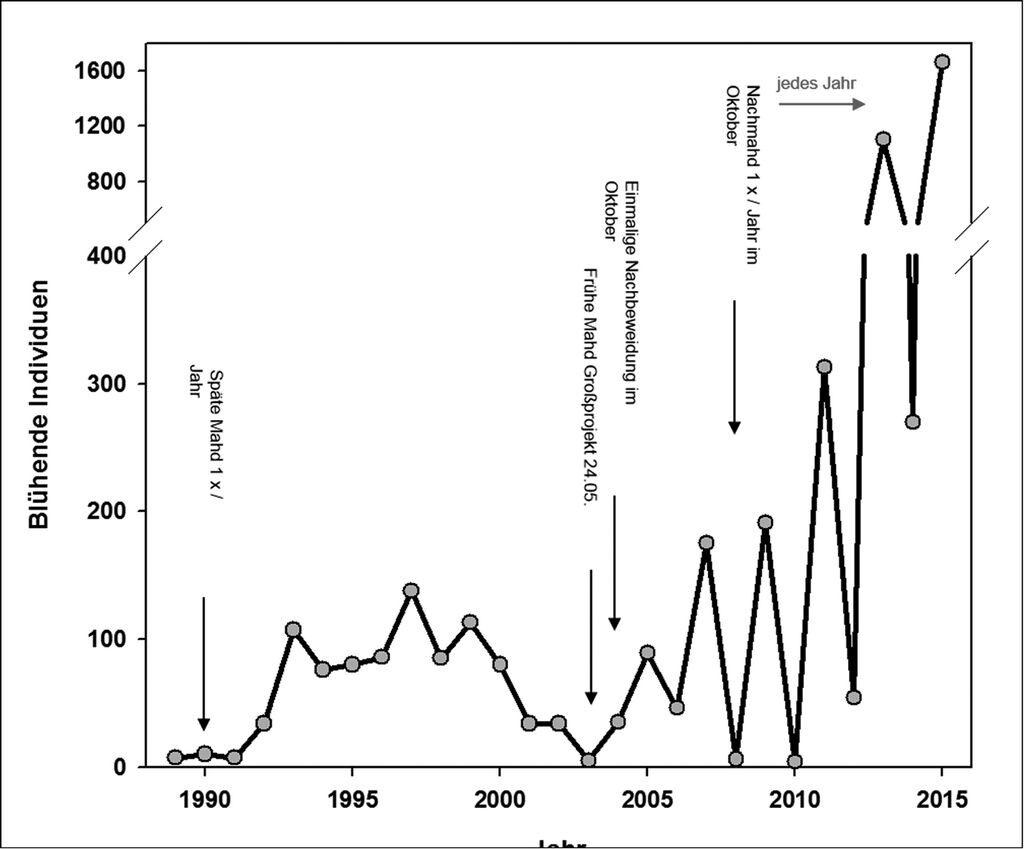
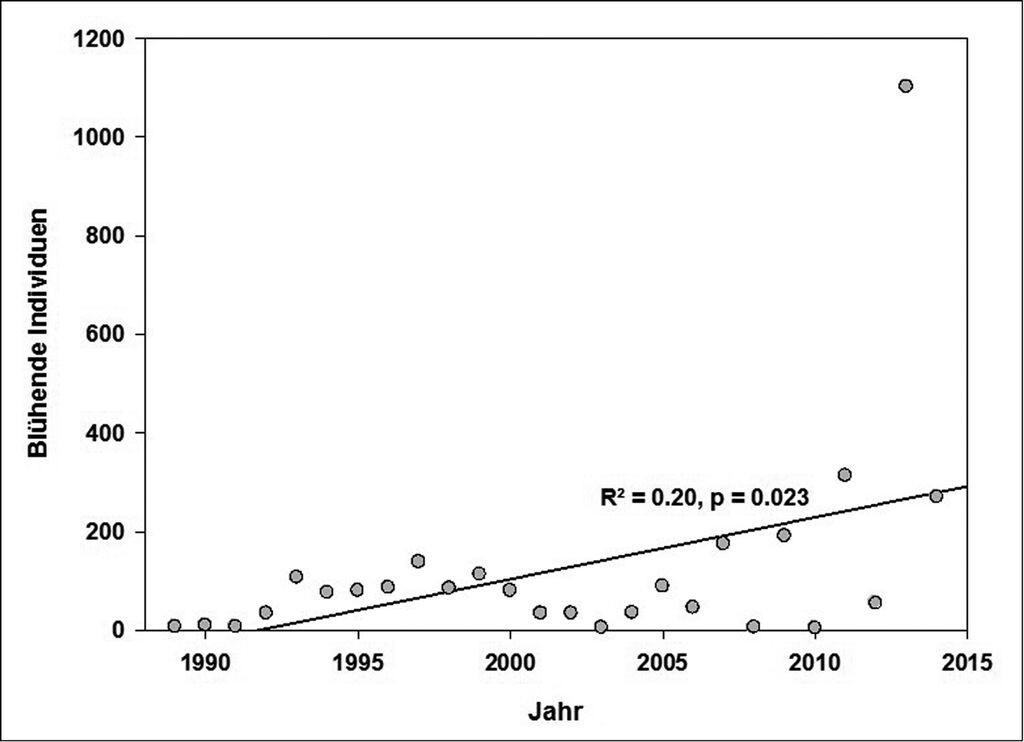
Die Population des Karpaten-Enzians am Geisingberg drohte Anfang der 1990er-Jahre zu erlöschen (Abb. 8). Die Populationsdynamik weist seit 2005 ursächlich kaum zu erklärende starke Bestandsschwankungen zwischen ungeraden und geraden Jahren auf. Insgesamt besteht aber ein signifikant positiver Trend für die Population (Abb. 9). Ende Juli 2015 wurden im ursprünglichen Alt-Bestand des Karpaten-Enzians 1660 blühende Pflanzen gezählt. Damit war 2015 das bisher individuenstärkste Jahr seit Beginn des Populations-Monitorings 1989. Von 2009 bis 2013 erfolgte ein exponentieller Anstieg der Anzahl blühender Individuen in ungeraden Jahren. Dies scheint sich 2015 leicht abzuschwächen und geht möglicherweise in ein typisches Wachstum einer Population an ihrer Umweltkapazitätsgrenze über. Die Individuenzahlen der geraden Jahre sind 2012 und 2014 ebenfalls sehr stark angestiegen. Dieser Trend hat 2016 jedoch nicht angehalten.
4 Diskussion
4.1 Schwankungen der Pflanzengrößen in den Erhaltungskulturen
Im November und Dezember 2013 wurden hinter dem Verwaltungsgebäude des botanischen Gartens in Schellerhau Bäume entnommen, die die Erhaltungskultur des Enzians stark beschatteten. Bei der regelmäßigen Größenvermessung der Einjährigen im September 2014 stellte sich nun heraus, dass die Pflanzen im Vergleich zu den Vorjahren erheblich größer und nun ähnlich groß wie die Enzianpflanzen aus der Erhaltungskultur im Garten Naundorf waren. Gleichzeitig wurde im Herbst 2013 die bisherige Enzian-Kulturlinie des botanischen Gartens durch neues Saatgut, das aus der Wild-Population entnommen wurde, ersetzt. Der sprunghafte Anstieg der Größen der einjährigen Enziane in der Erhaltungskultur Schellerhau von 2013 auf 2014 (vgl. Abb. 6) muss nun entweder mit der Entnahme der beschattenden Bäume und des dadurch verbesserten Mikroklimas zu tun haben oder liegt darin begründet, dass die bisher in der Kultur des Botanischen Gartens Schellerhau befindliche Linie aufgrund genetischer Drift und/oder schon auftretender Inzuchtdepressionen kontinuierlich an Fitness verloren hat, was in Erhaltungskulturen relativ schnell gehen kann (vgl. Cockburn 1995, Lauterbach et al. 2012).
Abb. 6 zeigt nun, dass die Größen der einjährigen Enziane im Garten Schellerhau seit 2010 mehr oder weniger kontinuierlich zurückgegangen sind bzw. ebenfalls in zweijährigem Turnus zu schwanken scheinen: Die Einjährigen 2015 entsprechen mit ihrer geringen Größe nämlich wieder den Größen der Einjährigen 2013. Dies spricht aber dafür, dass das Phänomen der plötzlich gewachsenen Größen 2014 vermutlich weniger auf die Entnahme der beschattenden Gehölze im Winter 2013/2014 zurückzuführen ist, denn dann sollten die Jungpflanzen weiterhin ähnlich groß wie 2014 gewesen sein. Damit kommen die zwei alternativen Erklärungen in Betracht:
(1) Wenn die Größenunterschiede auf unterschiedlich günstige klimatische Bedingungen des jeweiligen Jahres zurückzuführen wären, sollten die Größen der Jungpflanzen im Garten Naundorf ähnlich schwanken und 2015 ebenfalls deutlich kleiner als 2014 sein.
(2) Es lag tatsächlich schon eine Inzuchtdepression in der Kulturlinie des Gartens Schellerhau vor. Dann sollten die 18 Jungpflanzen, die auf frisches Saatgut, das 2014 auf Fläche 1 geerntet wurde, zurückgehen, signifikant größer sein als die sechs Jungpflanzen, die auf Saatgut von der alten Kulturlinie des Botanischen Gartens Schellerhau zurückgehen.
In der Tat sind die Jungpflanzen im Garten Naundorf 2015 ebenfalls nur etwa halb so groß wie 2014 (Median 2015: 11mm, 2014: 22mm; vgl. Abschnitt 7), so dass Hypothese (1) als Erklärung im Moment am wahrscheinlichsten ist. Dies wird noch dadurch unterstützt, dass bzgl. Hypothese (2) im Mittel zwar Unterschiede zwischen den Größen der Herkünfte festzustellen waren (Herkunft F 1: 8mm, Herkunft BG Schellerhau, 7mm), diese aber nicht signifikant sind (U-Test: U=42,00, p=0,336). Damit ist Hypothese (2) als Erklärung des Phänomens eher unwahrscheinlich, auch wenn die Stichprobengröße bei Herkunft BG Schellerhau mit n=6 natürlich recht gering ist.
Zusammenfassend kann damit festgestellt werden, dass auch in den Erhaltungskulturen die Größen der (Jung-)pflanzen stark von den (klimatischen) Bedingungen des jeweiligen Jahres bestimmt werden.
4.2 Entwicklung der Wiederansiedlungsflächen
Auch wenn noch nicht abschließend beurteilt werden kann, ob die Wieder- bzw. Neuansiedlung von Beständen des Karpaten-Enzians auf den ausgewählten Flächen dauerhaft erfolgreich war, gibt die Entwicklung der Bestände Anlass zur Hoffnung auf eine längerfristige Etablierung. Die letzten Aussaaten und Auspflanzungen auf den Ausbringungsflächen wurden im Frühling 2014 vorgenommen. Zwar ist auf einigen Flächen die Anzahl der Pflanzen 2015 leicht zurückgegangen jedoch nicht auf allen (vgl. Tab. 2). Die leichten Rückgänge von 2014 auf 2015 können unterschiedliche Gründe haben. So können die Populationsgrößen von Vertretern der Gattung Gentianella generell stark schwanken (vgl. Brabec 2010, Brunzel 2012, Dolek et al. 2009, Fischer & Matthies 1997) und sogar deutliche genetische Unterschiede zwischen Generationen aufeinanderfolgender Jahre auftreten (Königer et al. 2012). Der bisherige Erfolg der Wiederansiedlungs-Maßnahmen des Karpaten-Enzians im Ost-Erzgebirge ist damit mit anderen erfolgreichen Ansiedlungsmaßnahmen hochgradig gefährdeter Arten vergleichbar (vgl. Diekman et al. 2015).
4.3 Populationsdynamik am Alt-Standort: intensives Management und langfristige Überlebenschancen
Die Wiesen am Geisingberg waren Anfang der 1990er-Jahre völlig verbracht und zum Teil schon verbuscht. Die umgebenden Steinrücken waren komplett mit Bäumen zugewachsen. In dieser Zeit war die Population in einem Jahr auf sieben blühende Individuen geschrumpft. Um den Bestand zu retten, wurde die Fläche wieder in eine regelmäßige Nutzung überführt. Zusätzlich hatte der damalige Eigentümer eines Teils der Geisingberg-Wiesen, der den Enzianbestand ebenfalls bewahren wollte, 1991 mehrfach Samen mit einer Gießkanne ausgesät. Im Rahmen von Erstpflegemaßnahmen wurden dann die Steinrücken freigestellt sowie die Wiesen entbuscht und in eine jährliche späte Mahd überführt. Dadurch stabilisierte sich der Bestand Mitte der 1990er-Jahre auf einem Niveau von 76 bis 138 blühenden Individuen, brach aber im trocken-heißen Jahr 2003 wieder auf fünf blühende Pflanzen ein. Nach einer erneuten, leichten Erholung nahm der Bestand auf dem Alt-Standort aber zum Ende der Dekade wieder dramatisch auf sechs in 2008 und vier blühende Pflanzen in 2010 (niedrigster Wert seit Beginn der Zählungen) ab.
Da also die bis dahin durchgeführten Maßnahmen offensichtlich nicht zu einem dauerhaften Erhalt führten, entschloss man sich in einem vom Bundesamt für Naturschutz (BfN) geförderten Naturschutzgroßprojekt, ab 2003 weitere Maßnahmen durchzuführen. So wurde 2003 eine Frühmahd durchgeführt und in 2004 die Wiesen im Herbst nachbeweidet. Seit 2008 wird im Oktober eine zweite Mahd durchgeführt, um den Aufwuchs und die Streuauflage während des Winters möglichst gering zu halten. Diesbezüglich wäre eine Nachbeweidung mit Schafen aufgrund der Bodenverwundungen und der damit verbundenen Schaffung von Keimbetten vermutlich ebenfalls günstig. Ab 2010 wurden weitere Steinrücken um das ursprüngliche natürliche Vorkommen auf der Hufeisenwiese herum freigestellt, so dass der Bestand in den folgenden Jahren deutlich weniger beschattet wurde und sich im Herbst weniger Laubstreu auf der Wiese ablagern konnte.
Es ist bisher nicht befriedigend möglich, die starken Schwankungen des Bestands auf dem Alt-Standort zu erklären. Neben dem Einfluss extremer Witterungsverhältnisse einzelner Jahre muss davon ausgegangen werden, dass die zwischenzeitlich bis auf wenige Individuen zusammengeschmolzene Population einer starken genetischen Drift unterliegen könnte und Inzuchteffekte nicht auszuschließen sind (vgl. Fischer & Matthies 1997, 1998; Königer et al. 2012, Reisch 2008). Ebenso können lokale Populationen von Gentianella relativ schnell Adaptionen an besondere Standortverhältnisse und damit lokale Besonderheiten zeigen (vgl. Grzes 2007). Für die nahe verwandte, ebenfalls zweijährige Art G. bohemica ist bekannt, dass isoliert liegende Populationen eine starke genetische Verarmung zeigen und sogar schon starke genetische Unterschiede zwischen Generationen aufeinanderfolgender Jahre festzustellen sind (Königer et al. 2012). Es ist daher auch nicht auszuschließen, dass die Population von G. lutescens im Osterzgebirge sich genetisch schon deutlich von den Populationen der nächstgelegenen Vorkommen in den Weißen Karpaten unterscheidet.
Ebenso ist es nicht möglich, die unerwartet deutliche positive Entwicklung des natürlichen Bestands auf dem Alt-Standort befriedigend auf eine oder mehrere Naturschutz-Maßnahmen zurückzuführen. Betrachtet man die Bestandsdynamik, so ist am wahrscheinlichsten, dass die seit 2008 in jedem Jahr durchgeführte Herbstmahd durch Konkurrenzreduktion einen erheblichen Beitrag zur positiven Entwicklung beigetragen hat. Aufgrund der positiven Korrelation zwischen hohen Keimraten und kühlen, schneereichen Wintern (vgl. Brunzel 2010, Brunzel & Sommer 2011) haben die Winter 2009/10 bis 2012/13 sicher ebenfalls zum starken Anstieg beigetragen.
Im ursprünglichen Bestand am Alt-Standort wurden neben der erwähnten Nachmahd keine weiteren, intensiven Management-Maßnahmen, wie sie in den neu begründeten Teil-Populationen nötig waren, durchgeführt. Hierzu zählt vor allem die Wässerung der neu begründeten Teil-Populationen in trockenen heißen Jahren.
Es ist damit in aller Deutlichkeit festzuhalten, dass in diesem ursprünglichen Bestand auf dem Alt-Standort eine positive Entwicklung ohne Intensiv-Maßnahmen nur mit gewöhnlicher Wiesennutzung möglich war. Gentianella lutescens gilt wie andere Vertreter der Gattung auch als trockenheits- und hitzeempfindlich (Brabec 2010, Brunzel 2010, Grzes 2007, Milberg 1994, Zillig et al. 2010). Darüber hinaus ist anzunehmen, dass sich eine zunehmende Atlantisierung der Winter negativ auf den Keimungserfolg der Art auswirkt (Brunzel 2010, Brunzel & Sommer 2011). Sollten die Populationen des Karpaten-Enzians also ohne Intensiv-Maßnahmen wie z.B. Wässerungen nicht überleben können, wäre der Sinn von intensiven Erhaltungsmaßnahmen wie der Etablierung von Erhaltungskulturen und Wiederansiedlungen vor dem Hintergrund prognostizierter klimatischer Entwicklungen fraglich. Aufgrund der geringen Anzahl von Vorkommen der Art in Europa konnte bisher keine Klimanische für G. lutescens mit statistischen Modellen berechnet werden (Ingolf Kühn, mündlich), mit deren Hilfe sich die Überlebenswahrscheinlichkeit der Art für das Osterzgebirge unter verschiedenen Klimawandelszenarien prognostizieren ließe.
Auch wenn aufgrund der Ökologie der Art anzunehmen ist, dass sie negativ auf prognostizierte klimatische Veränderungen reagieren wird, zeigt die Entwicklung des Bestands auf dem ursprünglichen Alt-Standort, dass diese Population trotz eines genetischen Flaschenhalses und der Häufung klimatisch ungünstiger Jahre offenbar anpassungsfähig genug ist, um eine extrem positive Entwicklung zu nehmen. Hieran wird deutlich, dass geeignetes Habitat-Management auch in Kombination mit z.T. aufwendigen Artenschutz-Maßnahmen, wie es Erhaltungskulturen und Wiederansiedlungs-Maßnahmen eben sind (vgl. Becker 2012, Röder & Kiehl 2008), großes Potential besitzen, mögliche negative Auswirkungen großklimatischer Veränderungen zu kompensieren.
Dank
Wir danken dem Förderverein für die Natur des Osterzgebirges und der GmbH Naturbewahrung Osterzgebirge sowie Friedemann Trittmacher für die konstruktive Zusammenarbeit während der Projektlaufzeit.
Zwei Projekte mit dem Titel Ex-Situ und In-Situ-Management des in Sachsen vom Aussterben bedrohten Enzians Gentianella lutescens wurden im Rahmen der Richtlinie Natürliches Erbe des Freistaates Sachsen gefördert. Ein drittes Folgeprojekt wird ab 2016 ebenfalls im Rahmen der Richtlinie Natürliches Erbe gefördert.
Literatur
Aeschimann, D., Lauber, K., Moser, D. M., Theurillat, J.-P. (2004): Flora alpina. Haupt, Bern.
Amt der Steiermärkischen Landesregierung (2007): Naturschutz in der Steiermark Geschützte Pflanzen. http://www.umwelt.steiermark.at/cms/dokumente/10218136_1563917/b8e60aa5/Geschützte-Pflanzen.pdf. Graz.
Becker, T. (2012): Die Steppenreliktart Astragalus exscapus eine Schlüsselart der Steppenreste Mitteleuropas? In: TMUEN, Hrsg., Steppenlebensräume Europas Gefährdung, Erhaltungsmaßnahmen und Schutz. Tagungsband 7, 69-90.
Becker, T., Becker, U. (2010): Successful transplantation of a harts tongue fern population (Asplenium scolopendrium L.) with ten years of monitoring. Tuexenia 30, 47-58.
Brabec, J. (2010): Monitoring horecku mnohotvareho ceskeho (Gentianella praecox subsp. bohemica) vCR (sezona 2010). Ms., 251p. + 28p. Elektronick apriloha. [Depon in: ust redn pracoviste AOPK CR, Praha].
Brunzel, S. (2010): Ex-situ-Kultivierung und In-situ-Management als Beitrag zum Artenschutz am Beispiel von Frauenschuh (Cypripedium calceolus), Sumpf-Gladiole (Gladiolus palustris), Böhmischer Enzian (Gentianella bohemica) und Karpaten-Enzian (Gentianella lutescens). Naturschutz und Landschaftsplanung 42 (5), 148-156.
, Sommer, M. (2011): Ex-Situ und In-Situ-Management des in Sachsen vom Aussterben bedrohten Enzians Gentianella lutescens im Rahmen der Richtlinie Natürliches Erbe. Endbericht des Förderprojekts. unveröff. Mskr.
Cockburn, A. (1995): Evolutionsökologie. G. Fischer, Stuttgart.
Deno, N.C. (1996): The first Supplement to the second edition of seed germination theory and practice. Self-published and self-distributed.
Diekmann, M., Müller, J., Heinken, T., Dupré, C. (2015): Wiederansiedlung von Gefäßpflanzen in Deutschland eine Übersicht und statistische Auswertung. Survey and statistical analysis of re-introductions in Germany. Tuexenia 35, 249-265.
Dolek, M. (2008): Artenhilfsprogramm Böhmischer Enzian (Gentianella bohemica), Ergebnisse 2006 bis 2008. Unveröff. Abschlussber.
, Thamke, I., Königer, J. (2009): Artenhilfsprogramm Böhmischer Enzian (Gentianella bohemica). Unveröff. Gutachten im Auftrag des Bayerischen Landesamtes für Umwelt (LfU), Augsburg.
Fischer, M., Matthies, D. (1997): Mating structure and inbreeding and outbreeding depression in the rare plant Gentianella germanica (Gentianaceae). Am. J. Bot. 84, 1685-1692.
, Matthies, D. (1998): RAPD variation in relation to population size and plant tness in the rare Gentianella germanica (Gentianaceae). Am. J. Bot. 85, 811-819.
Grzes, I.M. (2007): Does rare Gentianella germanica (Wild.) Börner originating from calamine spoils differ in selected morphological traits from reference populations. Plant Species Biology 22, 49-52.
Königer, J., Rebernig, C.A., Brabec, J., Kiehl, K., Greimler, J. (2012): Spatial and temporal determinants of genetic structure in Gentianella bohemica. Ecology and Evolution 2 (3), 636-648.
Lauterbach, D., Borgmann, P., Daumann, J., Kuppinger, A.-L., Listl, D., Martens, A., Nick, P., Oevermann, S., Poschlod, P., Radkowitsch, A., Reisch, C., Stevens, A.-D., Straubinger, C., Zachgo, S., Zippel, E., Burkart, M. (2015): Allgemeine Qualitätsstandards für Erhaltungskulturen gefährdeter Wildpflanzen. Gärtnerisch-Botanischer Brief 200, 16-39.
, Burkart, M., Gemeinholzer, B. (2012): Rapid genetic differentiation between ex situ and their in situ source populations: an example of the endangered Silene otites (Caryophyllaceae). Botanical Journal of Linnean Society 168, 64-75.
Milberg, P. (1994): Germination ecology of the endangered grassland biennial Gentianella campestris. Biological Conservation 70, 287-290.
Müller, F., Zoephel, B. (2008): Abschlussbericht Erarbeitung eines Konzeptes zur Sicherung und Entwicklung der Population des Karpatenenzians am Geisingberg bei Altenberg Osterzgebirge, April Juni 2008. Technische Universität Dresden, Institut für Botanik, Lehrstuhl für Botanik. Auftraggeber: Regierungspräsidium Dresden, Umweltfachbereich, Dienststelle Radebeul.
Reisch, C. (2008): Naturschutzgenetische Untersuchung von Gentianella campestris. Abschlussbericht der Universität Regensburg, Lehrstuhl für Botanik, im Auftrag der Regierung der Oberpfalz.
Röder, D., Kiehl, K. (2008): Vergleich des Zustandes junger und historisch alter Populationen von Pulsatilla patens (L.) Mill. in der Münchner Schotterebene. Tuexenia 28, 121-132.45
Uhlig, D., Müller, F. (2001): Zur Bestandssituation ausgewählter vom Aussterben bedrohter und stark gefährdeter Pflanzenarten im Osterzgebirge. Förderverein für die Natur des Osterzgebirges e.V. 1, 1-35.
Zillig, P., Lebermann, K., Morba, D., Arneth, G., Lauerer, M. (2010): Ex-situ-Erhalt des Böhmischen Fransenenzians: Erfahrungen bei der Erhaltungskultur im Botanischen Garten Bayreuth. Naturschutz und Landschaftsplanung 42 (3), 78-84.
-
Abb. 1: Der Karpaten-Enzian (Gentianella lutescens).
Carpathian gentian (Gentianella lutescens).Randolf Manderbach -
Abb. 2: Lage des Projektgebiets im Osterzgebirge mit den Probeflächen in Rot.
Location of the project area in the Osterzgebirge with application sites (in red). -
Abb. 3: Erhaltungskultur im Botanischen Garten Schellerhau mit Besichtigung der Projektbeteiligten und des LfULG Sachsen.
Conservation culture in the Botanical Garden Schellerhau; inspection of the participants of the project and of the State Agency for the Environment.Holger Menzer -
Abb. 4: Ausbringungsfläche mit 1-m²-Rasterung für das Jungpflanzen-Monitoring.
Application site with grids of 1 m² for the monitoring of the juvenile plants.Bernd König -
Abb. 5: Aussaat mit Erdsieben.
Seeding using sieves.Bernd König -
Abb. 6: Dynamik der Pflanzengrößen in den Erhaltungskulturen: Größen (Länge längstes Laubblatt in mm) einjähriger G. lutescens 2010 bis 2015.
Dynamics of the size of the plants in the conservation cultures: Sizes (length showing the longest foliage leaf in mm) of one-year old plants of G. lutestcens between 2010 and 2015. -
Abb. 7: Anzahl der Blüten in den neu- bzw. wiederbegründeten Teilpopulationen 2015.
Number of flowers in the newly or re-established partial populations in 2015. -
Abb. 8: Populationsdynamik (Anzahl blühender Individuen) über den gesamten Zeitraum 1989 bis 2015 und erstes (bzw. einziges) Jahr bestimmter Naturschutzmaßnahmen. Die Nachmahd im Herbst wird seit 2008 jedes Jahr durchgeführt.
Population dynamics (number of flowering individuals) over the complete period 1989 to 2015, and first (resp. only) year of certain nature conservation measures. The after-mowing in autumn has been conducted every year from 2008 onwards. -
Abb. 9: Hoch signifikante (p≤0.01) positive Bestandsentwicklung der Population am ursprünglichen, natürlichen Standort über alle Beobachtungsjahre.
Highly significant (p≤0.01) positive development of the population on its initial natural site over the whole monitoring period. -
Erhaltungskulturen zur Wiederansiedlung des Karpaten-Enzians im Osterzgebirge -
Erhaltungskulturen zur Wiederansiedlung des Karpaten-Enzians im Osterzgebirge -
Erhaltungskulturen zur Wiederansiedlung des Karpaten-Enzians im Osterzgebirge





Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.