Bestandsdynamik von Moor-Schmetterlingen in Mecklenburg-Vorpommern über 125 Jahre
Abstracts
Zur Bewertung der Populationsschwankungen von Schmetterlingen nährstoffarmer Moore in den letzten 125 Jahren in Mecklenburg-Vorpommern wurden Funddaten aus Literatur, Datenbanken sowie privaten und öffentlichen Sammlungen zusammengetragen. Nachfolgend fand eine Systematisierung und Visualisierung dieser Daten mittels eines Geographischen Informationssystems statt. Zudem wurden die Fundortnachweise, auf einzelne Jahre bezogen, ausgezählt. Ziel war es, Bestandsschwankungen zu detektieren, ihre möglichen Ursachen zu klären und auf die biozönotischen Wirkungen zum heutigen Zeitpunkt zu schlussfolgern.
Es stellte sich heraus, dass bestimmte Bestandstrends bei den untersuchten Schmetterlingsarten zum einen mit Änderungen in der Produktivität und Entwässerungseffizienz in der Landwirtschaft, zum anderen mit klimatischen Änderungen zusammenfielen. In der Folge wanderten Arten, die eine größere ökologische Amplitude hatten und auch außerhalb von Hochmooren leben konnten, offensichtlich massiv in die nährstoffarmen Habitate ein, was zu einem Wandel in der Artenzusammensetzung geführt hat bzw. führt. Als Schutzstrategien gegen diese Veränderungen sind v.a. ökologische Sanierungen der nährstoffarmen Moore (insbesondere ihrer Wasserverhältnisse) zu nennen, die auf einer gründlichen Analyse der Artenzusammensetzung und der ökologischen Defizite basieren müssen. Auch die Möglichkeit einer Umsiedlung von Moorarten sollte in den Sanierungskonzepten geprüft werden.
Spatio-temporal dynamics of tyrphobiontic/tyrphophilic butterflies and moths in nutrient-poor bogs of Mecklenburg-Western Pomerania Anthropogenic and climatic causes of population fluctuations during the past 125 years
The study investigated existing data on the evaluation of population fluctuations of butterflies and moths of low-nutrient moors in Mecklenburg - Western Pomerania during the past 125 years. It compiled all data available from literature, databases, as well as from private and public collections. The data were systematized and visualized using a Geographical Information System. Further, the findings of the various species were counted referring to single years. The analysis aimed to detect the above-mentioned fluctuations, to identify their causes, and to draw conclusions on current biocoenotic effects.
The results show that population fluctuations of the species investigated coincided on the one hand with productivity and de-watering measures of agriculture, and on the other hand with climatic trends. In addition, species with a greater ecological amplitude and those able to also live outside of bogs obviously have migrated massively into the nutrient-poor habitats. There they have caused a significant change of the species composition. The study suggests protection strategies, such as the environmental restoration of low-nutrient bogs (esp. In terms of their water conditions). These concepts have to base on a thorough analysis of species composition and ecological deficits. In this context the feasibility of a transfer of bog species should also be examined.
- Veröffentlicht am

Mating of the tyrphobiontic Cranberry Fritillary in a raised bog in Austria.
1 Einleitung
Nährstoffarme Moore bilden mit ihrem vielfach humiden und „kontinentaler“ temperierten Eigenklima ein Paläorefugium für Lepidopteren des boreo-alpinen bzw. euro-sibirischen Verbreitungstyps (Burmeister 1990, Sommer et al. 2015, Thiele & Luttmann 2015). Die Vegetationsstruktur dieser Moore weist zudem Elemente der borealen Zonen auf, so dass sich die Arten bis heute in ihnen erhalten konnten (de Lattin 1967). Auf Grund der Kleinflächigkeit und Isoliertheit der Habitate sowie ihres zumeist hohen anthropogenen Beeinträchtigungsgrads sind viele der dort siedelnden Taxa in ihrem Bestand gefährdet. Deshalb gehören sie oft zu den im Anhang 1 der Bundesartenschutzverordnung (BArtSchV) gelisteten, besonders oder streng geschützten Arten (u.a. Hochmoor-Scheckenfalter Boloria aquilonaris, Moosbeeren-Spanner Carsia sororiata, Gagelstrauch-Moor-Holzeule Lithophane lamda). Für lokal ausgestorbene, tyrphobionte Taxa gibt es im nordostdeutschen Tiefland so gut wie keine natürlichen Wiederbesiedlungsmöglichkeiten aus anderen nährstoffarmen Mooren, da diese häufig weit entfernt in der zersiedelten Kulturlandschaft liegen. Die stark angepassten Individuen sind daher auf naturnahe Lebensraumverhältnisse ihres Standortes angewiesen, werden nur selten verdriftet und haben dann kaum eine Chance, zufällig ein Moorhabitat zu erreichen.
Stellten bisher Entwässerungen der Moore in Verbindung mit großflächigen Veränderungen im Landschaftswasserhaushalt eine der stärksten Bedrohungen für das Ökosystem dar, so kommt seit einigen Jahren mit dem Klimawandel eine weitere hinzu. Steigen die Jahresmitteltemperaturen, wie vom IPCC (2013) prognostiziert, weiter, so wird sich nicht nur die Vegetation ändern, sondern es steht auch zu erwarten, dass sich die Struktur der Lepidopteren-Vergesellschaftungen umgestaltet.
In diesem Zusammenhang stellt sich die Frage, welche zoozönotischen Effekte der Klimaerwärmung bezogen auf die europäischen Lepidopteren bereits bekannt sind. Grundsätzlich muss dazu angemerkt werden, dass die Forschung zu diesem Themenkomplex erst begonnen hat (Hill et al. 2002, Radchuk et al. 2013, Settele et al. 2008) und keine abschließenden Ergebnisse zu erwarten sind. Nachfolgend sollen einige Beispiele genauer ausgeführt werden.
Parmesan et al. (1999) stellten erstmals eine nordwärts gerichtete und durch die klimatischen Änderungen getragene Verlagerung der Verbreitungsgrenzen von Schmetterlingen fest. Von den 35 beobachteten Arten, die nicht zu den Wanderfaltern gehörten, wiesen in den letzten hundert Jahren 63% eine Nordverlagerung von 35 bis 240km auf. Hingegen hatten nur 3% ihr Verbreitungsareal nach Süden ausgedehnt. Elder (2015) prognostiziert bis 2050 für Großbritannien eine Kolonisationswelle an Schmetterlingsarten aus südlicheren Bereichen (u.a. Spiegelchen Heteropterus morpheus, Mittelmeer-Zitronenfalter Gonepteryx cleopatra, Großer Wanderbläuling Lampides boeticus, Blauschwarzer Eisvogel Limenitis reducta), wohingegen einige typische britische Arten nur noch Verbreitungsgebiete in Cumbrien und im schottischen Hochland besiedeln werden (z.B. Schornsteinfeger Aphantopus hyperantus, Weißes C Polygonium c-album, Aurorafalter Anthocharis cardamines).
Im Hinblick auf diese Beobachtungen und Projektionen gilt es zu klären, in welcher Beziehung dazu Habitatdegradation und klimatischer Wandel stehen. Warren et al. (2001) haben sich an den jeweils nördlichen Verbreitungsgrenzen von 46 Schmetterlingsarten in Großbritannien damit auseinandergesetzt. Es wurde erwartet, dass die untersuchten Schmetterlinge generell positiv auf die Erwärmung reagierten, was die Wirkungen des Habitatverlustes mildern müsste aber Dreiviertel dieser Arten reagierten negativ. Somit überwogen die pessimalen Folgen des Habitatverlustes die postulierten positiven Wirkungen des Klimawandels. Das traf insbesondere bei den Spezialisten unter den Schmetterlingsarten zu, wohingegen die Generalisten ihre Habitate zumeist ausdehnen konnten. Fox et al. (2014) bestätigte im Wesentlichen diese Aussagen, fügte aber hinzu, dass Arten, die auf Böden mit niedrigem N-Gehalt und/oder in Offenlandhabitaten vorkommen, besonders betroffen sind. Das gilt natürlich in besonderer Weise für die stark spezialisierten Arten der Arm- und Zwischenmoore. Somit wird deutlich, dass auch in Zeiten des Klimawandels die anthropogen verursachten Degradationen in Mooren (u.a. Entwässerung, Torfabbau) einen bedeutenden Einfluss auf die Stabilität der Biozönosen haben.
In der Summation aller Faktoren zeigt sich, dass den hochspezialisierten Schmetterlingen der nährstoffarmen Moore besondere naturschutzfachliche Aufmerksamkeit entgegen gebracht werden muss. Da große Bestandsschwankungen bei diesen Taxa schon seit Jahrzehnten beobachtet werden, erscheint es sinnvoll, ihre historische Bestandsdynamik an einem regionalen Beispiel zu erfassen und zu bewerten. Daraus lassen sich unter Zugrundelegung bestehender klimatischer Trends und postulierter Hemerobiegrade Schlussfolgerungen für die Zukunft ziehen. Ziel ist es, aus diesen Daten Maßnahmen abzuleiten, die den Trend des lokalen Aussterbens von Arten abmildern und diese nachhaltig sowie langfristig in ihrem Bestand sichern.
2 Methode
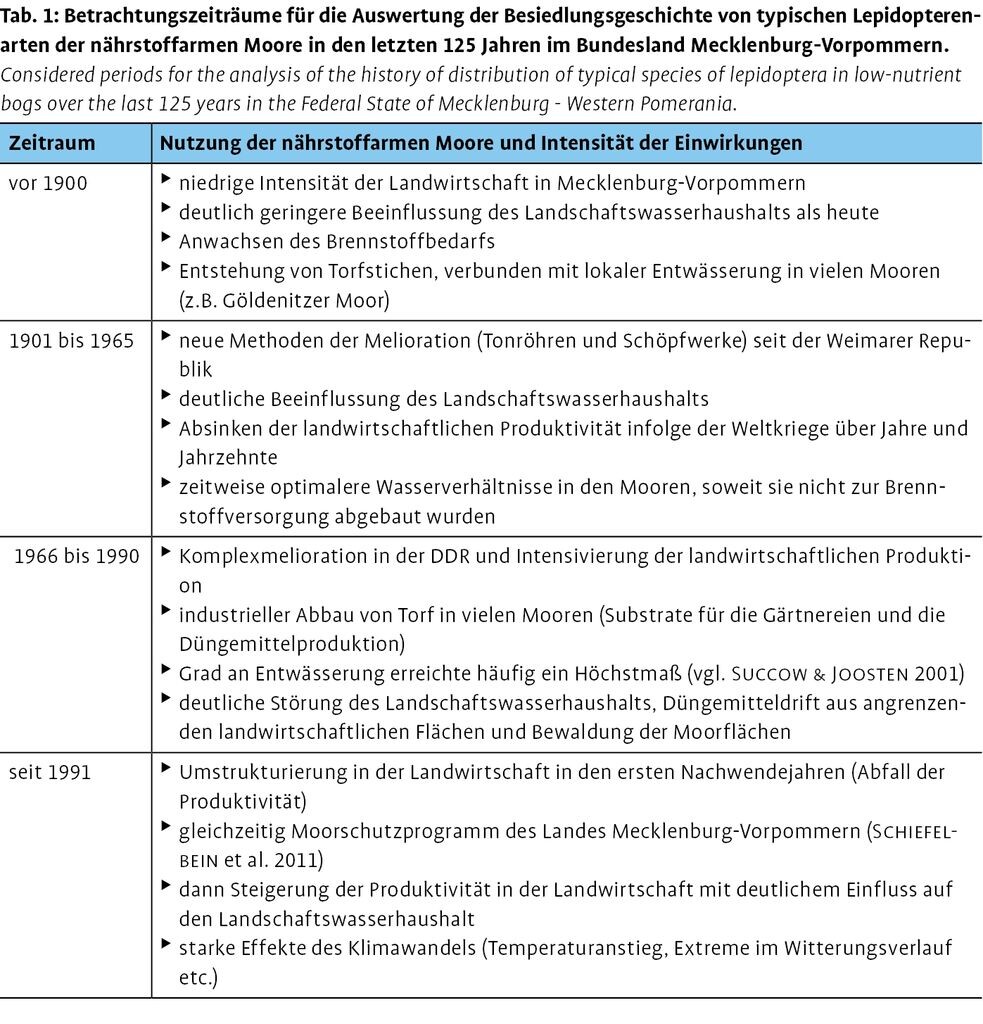
Um die historische Bestandsdynamik der tyrphobionten/-philen Schmetterlingsarten nährstoffarmer Moore am Beispiel Mecklenburg-Vorpommerns bewerten zu können, wurde stufenweise vorgegangen. Im ersten Schritt sind die als tyrphobiont bzw. -phil anzusprechenden Arten anhand einer parametergestützten Analyse abgegliedert und begründet worden. Dazu wurden Parameter genutzt, wie die Bindung der Raupen an moortypische Fraßpflanzen und Lebensräume sowie ihr Verbreitungstyp. Die Ergebnisse werden in Thiele (2014) und Thiele & Luttmann (2015) ausführlich dargestellt.
In einem zweiten Schritt wurden Karten zur Verbreitung der Taxa in Mecklenburg-Vorpommern erarbeitet (vgl. Thiele et al. 2015). Diesen liegen umfangreiche Literaturrecherchen und Befragungen von aktuell tätigen Entomologen zugrunde. Zudem wurden der Sammlungsbestand des Müritzeums in Waren (Landessammlungen) ausgewertet und Daten aus dem Landesamt für Umwelt, Naturschutz und Geologie Mecklenburg-Vorpommern (LUNG M-V) eingeholt. Darüber hinaus sind eigene, systematisch über einen Zeitraum von mehr als 40 Jahren erhobene Daten eingepflegt worden. Trotz größter Sorgfalt bei der Zusammenstellung der Daten kann aber nicht ausgeschlossen werden, dass es noch Datenquellen gibt, die den Autoren zum Zeitpunkt der Erstellung dieser Publikation nicht zugänglich waren.
Die Datensätze wurden zusammengestellt, digitalisiert und mittels eines Geographischen Informationssystems visualisiert. Bei älteren Meldungen und/oder Rasterdatendarstellungen mussten gewisse Lageungenauigkeiten in Kauf genommen werden. In die artbezogenen Darstellungen ist auch das jeweilige europäische Verbreitungsgebiet integriert worden, wobei Verbreitungsinformationen von Aarvik et al. (2009), Henriksen & Kreutzer (1982), Kudnra et al. (2015), Leraut (2006), Macek et al. (2008, 2012) und Skou (1991) eingingen. Die Karten können einschließlich einer kurzen Bewertung für alle untersuchten tyrphobionten/-philen Arten in Thiele et al. (2015) eingesehen werden. Aus Platzgründen werden in dieser Publikation nur die Verbreitungskarten der tyrphobionten Arten abgebildet.
Im dritten Schritt sollen die Erkenntnisse zusammengefasst und Schlussfolgerungen zu möglichen Veränderungen in der biozönotischen Struktur von Lepidopteren-Vergesellschaftungen gezogen werden. Besonderes Augenmerk wurde dabei auf die von Precker (2001) betrachteten 40 Regenmoore in Mecklenburg-Vorpommern gelegt. Zudem sind die Etappen in der Nutzungsgeschichte der Moore sowie klimatische Daten (auf eine Periode von zehn Jahren kumulierte durchschnittliche Niederschläge, Jahresdurchschnittstemperaturen) der Wetterstationen Schwerin (DWD 2015) ausgewertet worden. Um den potenziellen Einfluss von Nutzungsänderungen auch kartographisch nachvollziehen zu können, wurden als Betrachtungszeiträume in den Verbreitungskarten keine äquidistanten Zeitspannen gewählt. Stattdessen finden vor allem Perioden unterschiedlicher Nutzungsintensitäten in den Mooren Beachtung (vgl. Tab. 1).
Zusätzlich zur rein qualitativen Auswertung wurde das Auftreten der jeweiligen Arten pro Zeitraum (15 Jahre) und über alle Moore des Bundeslands ausgezählt. Nur bei guter bis sehr guter Datendichte sind auch detailliertere Auszählungen für kürzere Zeiträume vorgenommen worden. Der besseren Übersichtlichkeit halber wurden die Fundzahlen klassifiziert (15, 615, 1640, >40 Nachweise/Zeitraum). Dabei sind mehrere Funde an einem Ort und im gleichen Jahr standardisiert immer nur als ein Nachweis gewertet worden. So konnte vermieden werden, dass gut bearbeitete Moore gegenüber seltener befangenen Habitaten überwertet werden oder identische Fundnachweise aufgrund ihrer Nennung in unterschiedlichen Quellen mehrfach in die Zählung eingehen. Ziel dieser Auswertung war es, Perioden mit gehäuftem/verringertem Auftreten einer Art herauszukristallisieren.
3 Ergebnisse und Diskussion
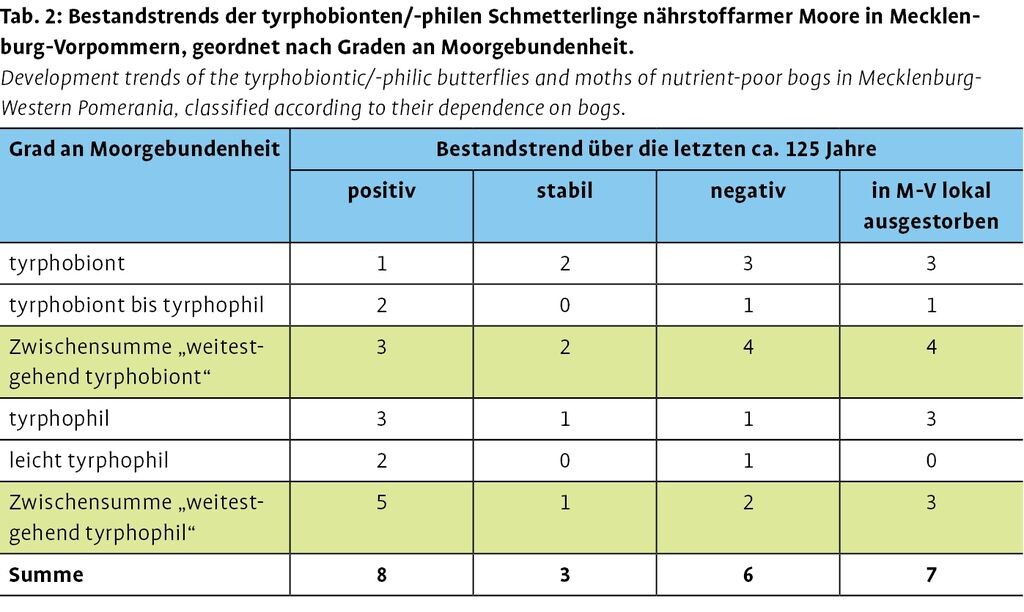
Von den 24 in Mecklenburg-Vorpommern relevanten tyrphobionten bzw. tyrphophilen Großschmetterlingsarten der nährstoffarmen Moore müssen sieben Arten als derzeit in Mecklenburg-Vorpommern nicht mehr nachweisbar gelten (Hochmoor-Gelbling Colias palaeno, Heidelbeeren-Silbereule Syngrapha interrogationis, Sumpfporst-Blütenspanner Eupithecia gelidata, Heidemoor-Bodeneule Protolampra sobrina, Randring-Perlmutterfalter Proclossiana eunomia, Heidebürstenbinder Orgyia antiquoides, Moorwiesen-Erdeule Diarsia dahlii; vgl. Tab. 2 und Thiele et al. 2015). Es ist aber möglich, dass über Verdriftung oder aktiven Zuzug (Oszillieren an der Verbreitungsgrenze) Moore von einzelnen Taxa wiederbesiedelt werden. Sechs Taxa haben einen zumeist stark negativen Bestandstrend (Hochmoor-Scheckenfalter Boloria aquilonaris [Abb. 1], Hochmoor-Bläuling Vacciniina optilete, Moosbeeren-Spanner Carsia sororiata, Gagelstrauch-Moor-Holzeule Lithophane lamda, Großes Wiesenvögelein Coenonympha tullia, Ampfer-Kleinspanner Scopula corrivalaria). Nur bei drei Arten können die Bestände als stabil betrachtet werden, wenn sie auch immer schon selten waren (Hochmoor-Bodeneule Coenophila subrosea, Moor-Bunteule Anarta cordigera, Heidemoor-Rindeneule Acronicta menyanthidis).
Acht Arten weisen einen positiven Bestandstrend auf, was eine unerwartet hohe Zahl darstellt (Gefleckter Rauschbeerenspanner Arichanna melanaria, Haworths-Mooreule Celaena haworthii, Moor-Stengeleule Amphipoea lucens, Rollflügel-Holzeule Lithomoia solidaginis, Moor-Motteneule Hypenodes humidalis, Waldmoorspanner Itame brunneata, Rundflügel-Flechtenbärchen Thumata senex, Purpurstreifen-Zwergspanner Idaea muricata). Es wird aber deutlich, dass der größte Teil dieser Gruppe von den „weitestgehend tyrphophilen Arten“ mit größerer ökologischer Amplitude gestellt wird. Hingegen rekrutieren sich die Gruppen mit negativem Bestandstrend bzw. mit fehlenden aktuellen Nachweisen vornehmlich aus tyrphobionten Arten mit enger ökologischer Amplitude. Für die tyrphobionten Arten ist ihre raum-zeitliche Verbreitung in den Abb. 2 bis 10 verdeutlicht. Zusätzlich wird die europäische Verbreitung kartographisch dargestellt.
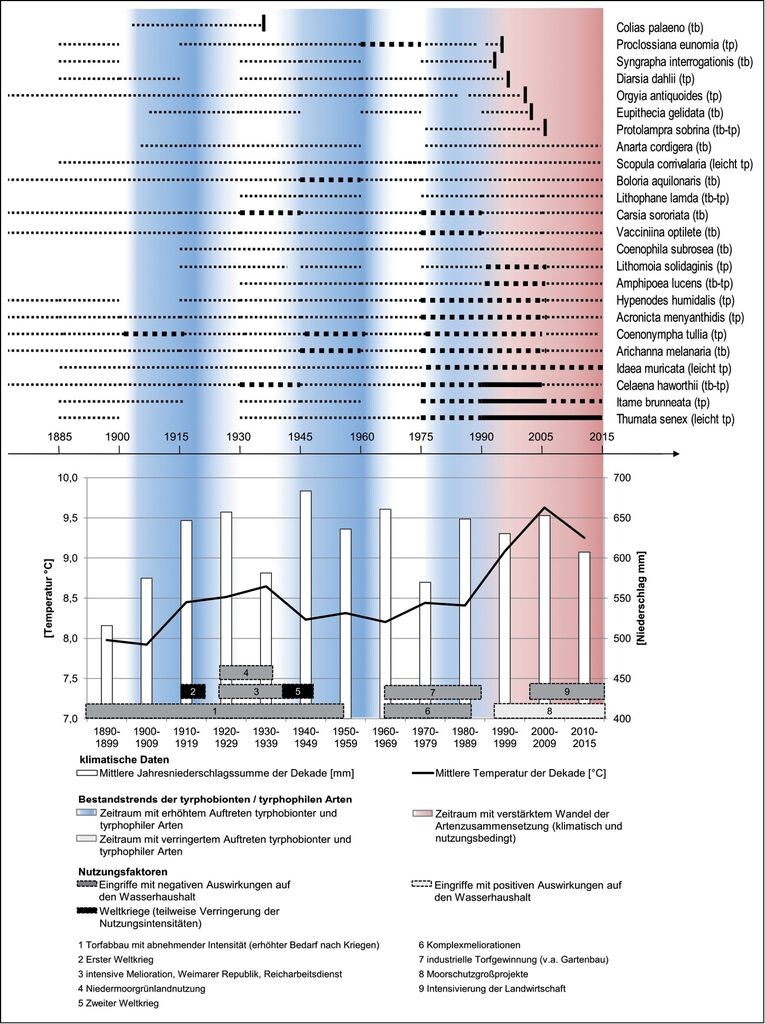
Die Ergebnisse der Auszählung von Fundnachweisen, bezogen auf Intervalle von jeweils 15 Jahren, sind in Abb. 11 dargestellt. Dort repräsentieren die Dicke und Durchgängigkeit der Striche die kategorisierte Anzahl an Fundmeldungen. Für die Zeitspannen ohne Striche liegen keine Meldungen vor. Die Arten sind von oben nach unten nach dem Zeitpunkt ihres letzten Nachweises bzw. nach steigender Anzahl der Fundmeldungen in den letzten drei betrachteten Zeiträumen geordnet. Im Hintergrund wurden die Jahresmitteltemperaturen und die auf zehn Jahre kumulierten durchschnittlichen Niederschlagsmengen mitgeführt. Außerdem sind u.a. auf den Wasserhaushalt der Moore wirkende (Nutzungs-) Faktoren zeitlich eingeordnet und temporäre Abschnitte mit erhöhtem bzw. verringertem Auftreten tyrphobionter/-philer Schmetterlinge gekennzeichnet. Es zeigen sich mehrere Tendenzen:
Stenotope Arten, die selten nachgewiesen worden sind, hat es schon immer in den nährstoffarmen Mooren Mecklenburg-Vorpommerns gegeben.
Perioden mit hoher Produktivität in der Landwirtschaft und/oder Einführung neuer Entwässerungsmethoden (Störungen im Landschaftswasserhaushalt, Entwässerung von Mooren zur Gewinnung von landwirtschaftlichen Flächen, Nährstoffdrift im Zuge von Düngung) fallen oft mit einem Populationsrückgang von stenotopen Moorarten zusammen (z.B. Weimarer Republik Einführung von Tonröhren und Pumpwerken; 1965-1980 Komplexmelioration).
In niederschlagsreicheren Perioden kam es bis in die 1990er Jahre immer zur Erholung vieler Bestände an tyrphobionten/-philen Arten in zahlreichen Mooren.
Das lokale Aussterben des Hochmoorgelblings (Colias palaeno) dürfte im Wesentlichen eine Folge geringer Niederschläge in den 1920er und 1930er Jahren sein. Zudem sind in diesem Zeitraum effizientere Entwässerungsmethoden entwickelt worden.
Mit Beginn des Klimawandels kommt es zum vermehrten Aussterben von stenotopen Moorarten. Gleichzeitig erhöht sich die Effizienz der Landwirtschaft im Umfeld der Moore, was die komplexe ökologische Sanierung dieser Ökosysteme (insbesondere durch Herstellung naturnaher Wasserverhältnisse) vielfach erschwert oder verhindert.
Die Wirkungen des Klimawandels spiegeln sich darüber hinaus durch einen statistischen Zusammenhang zwischen sinkendem Anteil der tyrphobionten Arten an den Gesamtnachweiszahlen und ansteigender mittlerer Jahrestemperatur (Korrelationskoeffizient nach Pearson r = 0,77 mit Irrtumswahrscheinlichkeit p = 0,008) wider.
Arten, die eine größere ökologische Amplitude haben und auch außerhalb von Hochmooren leben können, wandern massiv in die nährstoffarmen Habitate ein, was zu einem Wandel in der Artenzusammensetzung geführt hat und weiter führen wird.
Zusammenfassend kann man konstatieren, dass sich in der ursprünglich aus meist stenotopen Arten bestehenden Biozönose nährstoffarmer Moore sukzessive tyrphophile Arten durchgesetzt haben. Diese Verschiebung hat mit dem Einsetzen des Klimawandels deutlich an Fahrt gewonnen. Arten mit breiterer ökologischer Amplitude drängen verstärkt in die Moore und nehmen dort die Nischen strenger angepasster, tyrphobionter und tyrphophiler Taxa ein. Dabei erreichen sie oft hohe Abundanzen. Es steht zu erwarten, dass sich dieser Trend verstetigt und beschleunigt.
4 Ableitung von Schutzmaßnahmen
Die o.g. Befunde sind grundsätzlich auf zwei Ursachen zurückzuführen:
Störungen im Wasserhaushalt der Moore und der umgebenden, zumeist landwirtschaftlich genutzten Flächen (inkl. sekundärer Folgen, wie Bewaldung, Änderungen in der Vegetationszusammensetzung und Nährstoffdrift),
Klimawandel (u.a. erhöhte Temperaturen, Extremwetterlagen, veränderte Niederschlagsverhältnisse).
Um mögliche Schutzmaßnahmen ableiten zu können, muss geklärt werden, wodurch vornehmlich Probleme bei der Anpassung von Schmetterlingsarten an den Klimawandel entstehen und welche Rolle dabei anthropogen verursachten Veränderungen zukommt.
Mit Sicherheit gibt es dabei nicht „das eine Problem“, sondern einen ganzen, untereinander interagierenden Problemkreis (u.a. auch Nährstoffeinträge in Moore). Da die Forschung bezüglich der klimatischen Wirkungen aber erst am Anfang steht, können nur einige Hinweise gegeben werden. So ist vom Hochmoorgelbling (Colias palaeno) bekannt, dass Jungraupen in trockenen Habitaten einer erhöhten Mortalität unterliegen (Bräu et al. 2013). Auch Einzelereignisse mit extremen Witterungen, wie sie typisch für den Klimawandel sind, können verheerend wirken. Fehlende Schneebedeckung (Dasse 1934, zitiert in Maey 1986 und Bräu et al. 2013), und damit der Mangel an Frost- und Austrocknungsschutz, wirken auf die Organismen gefährdend. Tiefe Temperaturen können durch den niedrigen Super-Cooling-Point der Raupen von 26°C zwar gut überstanden werden, liegen sie aber bei oder über 0°C, so führen Schimmelbefall und Fäulnis u.U. zum Absterben der Larven (Dolek et al. 2008 und Bräu et al. 2013). Auch Thiele et al. (2013) kommen bezogen auf die prognostizierte Reaktion xylobionter Käfer auf die Klimaerwärmung zu einem ähnlichen Ergebnis.
Van Dyck et al. (2015) beschreiben die thermische Plastizität von ektothermen Organismen als eine Anpassung, die es gestattet, bei verlängerten wärmeren Bedingungen eine weitere Generation von Tieren hervorzubringen. Durch den klimatischen Wandel wird diese Möglichkeit für viele Arten von Insekten eröffnet. Wie sich beim Mauerfuchs (Lasiommata megera) herausstellte, reicht aber häufig die Zeit nicht aus, um vor dem Winter die Entwicklung abzuschließen. Dadurch kann es zu einer deutlichen Schwächung der Populationen bis hin zum lokalen Aussterben kommen.
Bei den Schmetterlingen nährstoffarmer Moore müssen zusätzlich noch die Isoliertheit, die extreme Bindung vieler Arten an das Habitat und das spezifische „kühlere“ Eigenklima der Moore Beachtung finden. Zudem tritt der oben beschriebene Wandel in den Biozönosen bereits seit Anfang der 1990er Jahre auf und man muss die beobachteten Wirkungen einfach anerkennen. Sie sind oftmals nicht mehr umkehrbar. Auch wenn die Moorvegetation und damit die Fraßpflanzen für viele Schmetterlingsarten der nährstoffarmen Moore noch lange vorhanden sind, können die Lepidopteren mit ihren größeren und komplexeren Raumansprüchen in einem degradierten Moor schnell lokal aussterben. Um das zu verhindern, sollten alle Bemühungen darauf gerichtet sein, die Moorhabitate so optimal wie möglich ökologisch zu sanieren. Kompromisse führen oft zu keinen hinreichenden Ergebnissen, v.a. wenn sie den Wasserhaushalt betreffen.
Wichtig ist häufig auch die Reduzierung des Waldanteils, kann doch so effektiv die Verdunstung gesenkt werden, was automatisch zu einer Verbesserung der Wasserverhältnisse führt. Zudem sind viele Moorinsekten schattenfliehend und können allein durch die Beschattung an den Rand des lokalen Aussterbens gedrückt werden. Mit der ökologischen Sanierung von Mooren kann Zeit für eine Anpassung dieser Schmetterlinge an den Klimawandel gewonnen werden. Stenotopen Arten wird das wenig nutzen. Sie werden in vielen Fällen angesichts der beschriebenen biozönotischen Veränderungen nicht zu halten sein. Aber schon bei der Gruppe der tyrphobionten bis tyrphophilen Taxa kann ein Anpassungsprozess postuliert werden.
Als letztes Mittel sei auf eine Wiederansiedlung solcher Arten nach erfolgter Sanierung verwiesen. Die Erfolge sind dabei sehr wechselnd. Ist eine hinreichend starke Spenderpopulation vorhanden und sind die „Rezipienten-Habitate“ hinreichend „gereift“, so sollte ein Versuch nicht ausgeschlossen werden. Wesentlich ist aber die Primärdatengewinnung über wissenschaftlich orientierte Erfassungen. Nur dadurch können der Zustand der Biozönose bestimmt und damit die Maßnahmen für ein nachhaltiges Management getroffen werden.
Dank
Die Autoren möchten sich beim Landesamt für Umwelt, Naturschutz und Geologie Mecklenburg-Vorpommern für die Bereitstellung von Fundortdaten zu den tyrphophilen und tyrphobionten Schmetterlingsarten bedanken. Ein herzlicher Dank geht auch an Dipl.-Biol. Renate Seemann im Müritzeum in Waren/Müritz, die ihren Fundus für die Auswertung bereitwillig öffnete. Zudem sei allen Entomologen des Entomologischen Vereins Mecklenburg gedankt, insbesondere Heinz Tabbert (Negast) und Uwe Deutschmann (Buchholz), die mit ihren Fundortangaben zum Gelingen des Vorhabens beitrugen.
Literatur
Aarvik, L., Hansen, L.O., Kononenko, V. (2009): Norges Sommerfugler. Håndbok over Norges dagsommerfugler og nattsvermere. Norsk entomologisk forening, Naturhistorik museum, Oslo, 432S.
BArtSchV, Verordnung zum Schutz wild lebender Tier- und Pflanzenarten (Bundesartenschutzverordnung) vom 16. Februar 2005, BGBl. I S. 258, zuletzt geändert am 29. Juli 2009, BGBl. I S. 2542.
Bräu, M., Bolz, R., Kolbeck, H., Nummer, A., Voith, J., Wolf, W. (2013): Tagfalter in Bayern. Ulmer, Stuttgart, 784S.
Burmeister, E.-G. (1990): Die Tierwelt der Moore (speziell der Regenmoore). In: Göttlich, K., Hrsg., Moor- und Torfkunde, Schweizerbart, Stuttgart, 29-47.
de Lattin, G. (1967): Grundriss der Zoogeographie. Gustav Fischer, Stuttgart, 602S.
Dolek, M., Bräu, M., Freese-Hager, A., Zapp, A., Thamke, I. (2008): Ursachenanalyse zum Rückgang des Hochmoorgelblings (Colias palaeno) in Bayern. Gutachten im Auftrage der Bayerischen Akademie für Naturschutz und Landschaftspflege (38S.). In: Bräu, M., Bolz, R., Kolbeck, H., Nummer, A. Voith, J., Wolf, W., 2013, Tagfalter in Bayern, Ulmer, Stuttgart, 784S.
DWD (2015): Klimadaten Deutschland, Tageswerte Niederschlag und Temperatur der Station Schwerin. Download: http://www.dwd.de (àKlima und Umwelt à Klimadaten à Klimadaten online frei; 30.04.2015).
Elder, C. (2015): Wild Britain 2050. BBC-Wildlife 33 (8), 26-36.
Fox, R., Oliver, T.H., Harrower, C., Parsons, M.S., Thomas, C.D., Roy, D.B. (2014): Long-term changes to the frequency of occurrence of British moths are consistent with opposing and synergistic effects of climate and land-use changes. J. Appl. Ecol. 51 (4), 949957.
Henriksen, H. J., Kreutzer I. (1982): The butterflies of Scandinavia in nature. Skandinavisk Bogforlag, Odense, 215S.
Hill, J. K., Thomas, C.D, Fox, R., Telfer, M.G., Willis, S.G., Asher, J., Huntlex, B. (2002: Responses of butterflies to twentieth century climate warming: implications for future ranges. Proceeding of the Royal Society B: Biological Sciences, 2163-2171.
IPCC (2013): Zusammenfassung für politische Entscheidungsträger. In: Stocker, T.F., Quin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Hrsg., Klimawandel 2013: Wissenschaftliche Grundlagen, Beitrag der Arbeitsgruppe I zum Fünften Sachstandsbericht des Zwischenstaatlichen Ausschusses für Klimaänderung, Cambridge University Press, Cambridge (UK), New York (USA).
Kudnra, O., Pennersdorfer, J., Lux, K. (2015): Distribution atlas of European butterflies and skippers. PEKS, Schwanfeld, 632S.
Leraut, P. (2006): Moths of Europe. Volume 1. Saturnids, Lasiocampids, Hawkmoths, Tiger Moths. N.A.P Editions, Verrières le Busisson, 272 pp.
Macek, J., Dvorák, J., Traxler, L., Cervenka, V. (2008): Motýli a housenky strední Evropy. Nocní motýli II. Academia, Praha, 490S.
, Procházka, J., Traxler, L. (2012): Motýli a housenky strední Evropy. Nocní motýli III. Academia, Praha, 417S.
Maey, H. (1986): Der Hochmoorgelbling Colias palaeno Linnaeus 1761 und seine Unterarten. Mitteilungen der Arbeitsgemeinschaft rheinisch-westfälischer Lepidopterologen: Beiheft 1. In: Bräu, M., Bolz, R., Kolbeck, H., Nummer, A., Voith, J., Wolf, W., 2013, Tagfalter in Bayern, Ulmer, Stuttgart, 784S.
Parmesan, C., Ryrholm, N., Stefanescu, C, Hill, J.K., Thomas, C.D., Descimon, H., Huntley, B., Kaila, L., Kullberg, J., Tammaru, T., Tennent, W.J., Thomas, J.A., Warren, M. (1999): Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature 399, 579-583.
Precker, A. (2001): Hydrogeologische Aspekte der Entstehung und der Möglichkeit der Restitution norddeutscher Moore. TELMA 31, 53-63.
Radchuk, V., Turlure, C., Schtickzelle, N. (2013): Predicting responses to climatic change requires all life-history stages. J. Anim. Ecol. (82), doi 10.1111/1365-2656.12032, 3-5.
Schiefelbein, U., Lenschow, U., Otto, D. (2011): Moorrevitalisierungen in Mecklenburg-Vorpommern eine Bilanz der letzten 20 Jahre. TELMA Beih. 4, 73-84.
Settele, J., Kudrna, O., Harpke, A., Kühn, I., van Swaay, C., Verovnik, R., Warren, M., Wiemers, M., Hanspach, J. Hickler, T., Kühn, E., van Halder, I., Veling, K., Vliegenthart, A., Wynhoff, I., Schweiger, O. (2008): Climatic Risk Atlas of European Butterflies. Pensoft, Sofia-Moscow, 712 pp.
Skou, P. (1991): Nordens Ugler. Danmarks Dyreliv. Apollo Books, Stenstrup, 565 pp.
Sommer, R.S., Thiele, V., Seppä, H. (2015): Use and misuse of the term „glacial relict“ in the Central European biogeography and conservation ecology of insects. Insect Conservation and Diversity, doi: 10.1111/icad. 12109, 3 pp.
Succow, M., Joosten, H. (2001): Landschaftsökologische Moorkunde. Schweitzerbart, Stuttgart, 622S.
Thiele, V. (2014): Welche Schmetterlingsarten sind an Arm- und Zwischenmoore im nordostdeutschen Tiefland gebunden? Virgo 17, 35-41.
, Luttmann, A. (2015): Tyrphobionte Schmetterlingsarten nährstoffarmer Moore. Eine parametergestützte Analyse zum Artenspektrum als Grundlage für Schutzstrategien mit Hinblick auf den Klimawandel. Natur und Landschaftsplanung 47 (4), 101-108.
Thiele, V., Luttmann, A., Liebe Edle von Kreutzner, K., Degen, B., Berlin, A., Lipinski, A., Niederstrasser, J., Koch, R., von dem Bussche, J. (2013): Durchführung einer Untersuchung zu den Folgen des Klimawandels in Sachsen-Anhalt. Wirkungen des Klimawandels auf europäisch geschützte Arten und Lebensräume. Ber. Landesamt f. Umweltschutz Sachsen-Anhalt 6, 231S. und div. Anh.
, Tabbert, H., Schuhmacher, S., Blumrich, B., Gohr, C. (2015): Die raum-zeitliche Verbreitung der Schmetterlinge von nährstoffarmen Mooren in Mecklenburg-Vorpommern. TELMA 45, 105-132.
Van Dyck, H., Bonte, D., Puls, R., Gotthard, K., Maes, D. (2015): The lost generation hypothesis: could climate change drive ectotherms into a developmental trap? Oikos 124 (1), 54-61.
Warren, M.S., Hill, J.K., Thomas, J.A., Asher, J., Fox, R., Huntley, B., Roy, D.B., Telfer, M.G., Jeffcoate, S., Harding, P., Jeffcoate, G., Willis, S.G., Greatorex-Davies, J.N., Moss, D., Thomas, C.D. (2001): Rapid responses of British butterflies to opposing forces of climate and habitat change. Nature 414, 65-69.
-
Abb. 1: Paarung des tyrphobionten Hochmoor-Scheckenfalters (Boloria aquilonaris) in einem österreichischen Hochmoor.
Mating of the tyrphobiontic Cranberry Fritillary in a raised bog in Austria. -
Abb. 210: Raum-zeitliche Verbreitung tyrphobionter Schmetterlingsarten der nährstoffarmen Moore in Mecklenburg-Vorpommern (vier Zeitebenen, Visualisierung tyrphophiler Lepidopteren vgl. Thiele et al. 2015, Kartengrundlage: © GeoBasis DE/M-V 2015).
Spatio-temporal distribution of tyrphobiobtic butterfly and moth species in low-nutrient bogs in Mecklenburg-Vorpommern (four temporal levels, visualisation of thyrophilic lepidoptera see THIELE et al. 2015, map base © GeoBasis DE/M-V 2015). -
Abb. 11: Dynamik der biozönotischen Veränderungen bei den tyrphobionten (tb)/-philen (tp) Schmetterlingen unter Berücksichtigung klimatischer Daten und anthropogen bedingter Einflüsse in den Mooren Mecklenburg-Vorpommerns. Die Dicke bzw. Durchgängigkeit der Striche repräsentiert die kategorisierte Anzahl an Fundmeldungen.
Dynamics of biocoenotic changes of tyrphobiontic (tb) / tyrphophilic (tp) butterflies and moths, considering data on climate and anthropogenic influences in the bogs of Mecklenburg - Western Pomerania. The thickness resp. consistency of the lines represents the categorized number of findings. -
Bestandsdynamik von Moor-Schmetterlingen in Mecklenburg-Vorpommern über 125 Jahre -
Bestandsdynamik von Moor-Schmetterlingen in Mecklenburg-Vorpommern über 125 Jahre -
Bestandsdynamik von Moor-Schmetterlingen in Mecklenburg-Vorpommern über 125 Jahre



Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.