Temporärer und permanenter Nahrungsraumverlust für Fledermäuse
Abstracts
Landschaftspflege fördert Lebensqualität und Biodiversität im urbanen Raum, kann aber auch Funktionsräume geschützter Arten beeinflussen. Um eine Beeinträchtigung zu vermeiden, gilt es, wichtige Lebensräume zu schonen. Der Beitrag zeigt, wie ein Konfliktfeld durch gezielte Maßnahmenplanung und sensible Umsetzung minimiert wurde: Die Sanierung eines Stadtteiches sollte dessen ökologische Funktion, Zustand und Ästhetik verbessern.
Die Maßnahmen standen im Konflikt mit seiner Funktion als Hauptnahrungsraum für eine lokale Population der Wasserfledermaus (Myotis daubentonii). Es wurde vermutet, dass der Nahrungsraumverlust zu einer erheblichen Beeinträchtigung führen könnte, da besonders reproduzierende Weibchen und Jungtiere auf eine genügende Nahrungsverfügbarkeit angewiesen sind. Eine Maßnahmenumsetzung im Winter war nicht durchführbar. Durch die Berücksichtigung der spezifischen Lebensraumnutzung demographischer Einheiten konnten gezielte Maßnahmen zur Konfliktminimierung und Kompensation entwickelt werden. Ein begleitendes Monitoring überprüfte den Erfolg der Umsetzung. Somit konnten Detailinformationen zu geschützten Arten die Umsetzung eines problematischen Eingriffs ermöglichen, ohne lokale Populationen erheblich zu beeinträchtigen.
Temporal and permanent loss of foraging habitats for bats How to minimize conflicts using the example of Myotis daubentonii at an urban pond
Landscape management promotes the quality of life and biodiversity in urban areas but also can influence functional habitats of protected species. To prevent detrimental effects it is essential to protect these habitats. The paper illustrates how a potential conflict could be minimized by specific measures and a sensitive implementation. The restoration of an urban pond aimed to improve its ecological functions, general condition, and visual appearance. These measures conflicted with its functionality as a foraging habitat for a local population of Daubentons bat (Myotis daubentonii). The loss of this foraging habitat could have had a significant detrimental effect on the population as in particular juveniles and actively reproductive females depend on a sufficient availability of food. The implementation of the measures in winter was not possible. By considering the explicit habitat use of these demographic units specific measures for conflict minimization and compensation could be developed. An accompanying monitoring validated the success of the implementation. The approach showed that detailed information on protected species allows to implement a problematic measure without significant detrimental effects on the local bat population.
- Veröffentlicht am

1 Einleitung
Eine wichtige Aufgabe der Landschaftspflege ist die Sicherung der Lebensraumqualität geschützter Arten, deren Vorkommen allerdings die naturschutzrechtlichen Anforderungen an landschaftspflegerische Eingriffe erhöht (Zucchi 1999).
Alle einheimischen Fledermausarten sind streng geschützt. Daher dürfen keine landschaftspflegerischen Maßnahmen durchgeführt werden, die zu einer erheblichen Beeinträchtigung lokaler Populationen führen könnten. Gleichzeitig können städtische Strukturen als Lebensraum für diese Artengruppe dienen. Einige Arten bevorzugen Dachböden oder Gebäudespalten als Tagesquartiere. Baumreihen, Hecken u.a. linienförmige Stadtstrukturen dienen als Flugwege in die Nahrungsräume, wie z.B. Alleen mit Straßenbeleuchtung, Stadtwälder und urbane Gewässer (Simon et al. 2004).
Urbane Gewässer können bei natürlichem Besatz ein Hotspot der Biodiversität sein. Neben Wasserlebewesen finden dort auch terrestrische Tiergruppen lebensnotwendige Ressourcen vor (Ineichen et al. 2012). Gleichzeitig befinden sich solche Gewässer häufig in Erholungsräumen, werden durch anthropogene Schadstoffe belastet und ihre Uferbefestigung destabilisiert sich. Dies führt dazu, dass sie aus landschaftspflegerischen und ökologischen Gründen gestaltet und gepflegt werden müssen, wodurch allerdings ein Konfliktfeld zum Artenschutz entstehen kann.
Ein Praxisbeispiel hierfür ist das Konfliktfeld zwischen der Sanierung eines Stadtteiches und dem dadurch entstehenden Nahrungsraumverlust für Wasserfledermäuse (Myotis daubentonii, Abb. 1) in Gießen. Bei einem temporären Nahrungsraumverlust muss die Nahrungsverfügbarkeit für die gesamte Population gewährleistet bleiben, um eine erhebliche Beeinträchtigung zu vermeiden.
Ziel der vorliegenden Studie ist es, die Entwicklung und Umsetzung konfliktminimierender Maßnahmen darzustellen. Dabei standen folgende Fragen im Mittelpunkt:
1. Welche Nahrungsräume werden in welchem Umfang von Alt- und Jungtieren der lokalen Wasserfledermauspopulation genutzt?
2. Kann der temporäre Verlust nahegelegener Nahrungsräume durch die intensivierte Nutzung alternativer Nahrungsräume kompensiert werden?
3. Kommt es zu einer Änderung der Habitatnutzung der lokalen Population als Reaktion auf den temporären Nahrungsraumverlust?
4. Kann ein permanenter Nahrungsraumverlust unmittelbar durch die Anlage neuer Nahrungsräume kompensiert werden?
2 Untersuchungsgebiet
Am Gießener Stadtrand befinden sich in einem Park mehrere Gewässer, darunter der Schwanenteich. Dieser ca. 39000 m2 große Teich wurde im Jahre 1935 in Verbindung mit einer südöstlich angrenzenden Kastanienallee und der Regulierung eines nordwestlich angrenzenden Bachlaufs angelegt. Durch die Ufervegetation kommt es zu regelmäßigem Laubeintrag, der zu einer hohen Eutrophierung führt. Die Dimensionen der Auffangbecken für Abwässer sind zu klein, um den Schadstoff-Eintrag aus dem angrenzenden Straßenverkehr (ca. 20m entfernt) und dem benachbarten Wohngebiet (ca. 50m) abzufangen (Abb. 2). Frischwasser wird manuell durch einen aufgestauten Bachlauf zugeführt.
Vorlaufende Analysen haben gezeigt, dass der Schwanenteich u.a. mit Schwermetallen aus dem Straßenverkehr und Abwasser belastet ist (Flache et al. 2015a). Eine Belastungseinschätzung ist indirekt über die chemischen Eigenschaften des Teichsediments möglich. Die Untersuchung der Eluat- und Feststoffparameter erfolgte anhand des LAGA-Merkblatts M20. Reduzierende Verhältnisse im Sediment wurden anhand des gebildeten Schwefelwasserstoffs organoleptisch festgestellt. Die Ergebnisse zeigten, dass der Schlamm die Eluatkriterien (d.h. herausgelöster Schadstoffgehalt) für eine Einstufung als Inertabfall (d.h. unbedenklich für Mensch und Umwelt) erfüllen würde, der organische Anteil jedoch oberhalb der Zuordnungswerte für Deponien der Klasse 2 liegt. Somit war das Material grundsätzlich verwertbar, aber nicht als mineralischer Abfall einzustufen.
Wiederholt wurden im Hochsommer verendete Wasservögel aufgefunden, deren Todesursache auf das im Bodenschlamm vorkommende Bakterium Clostridium botulinum zurückzuführen war. Der Zustand des Gewässers war zudem mit einer Geruchsbelästigung während Hitzeperioden verbunden. Der hohe Belastungsgrad des Stadtteichs sollte durch Abtragung der Bodensedimente, welches eine kosteneffizientere Maßnahme als z.B. Absaug- oder Umwälzverfahren darstellt, und Verbesserung der Frischwasser-Versorgung verringert werden. Eine Schlammentnahme ist allerdings nur in frostfreien Monaten und nach Ablassen des Wassers möglich. Der geeignete Zeitraum der Trockenlegung überschnitt sich somit mit der saisonalen Aktivitätszeit von Wasserfledermäusen und würde zu einem temporären Nahrungsraumverlust führen. Die letzte Sedimentreduzierung der Teichanlage wurde in den 1970er-Jahren durchgeführt, womit ein dringender Sanierungsbedarf des Schwanenteichs gegeben war.
Daneben kommen im Gießener Stadtpark mit dem Neuen Teich und dem Bachlauf der Wieseck zwei weitere Gewässer vor. Der Schadstoff-Eintrag in den ca. 65000 m2 großen Neuen Teich ist vermutlich durch die größere Entfernung zum Straßenverkehr und Wohngebieten (jeweils ca. 200 m entfernt) sowie umliegende Strukturen wie Kleingärten und die Wieseck geringer als der Eintrag in den Schwanenteich. Die Wasserqualität im Neuen Teich wird durch Schilfbewuchs und künstliche Belüftung verbessert. Die Wieseck verläuft aus nordwestlicher Richtung zwischen Schwanenteich und Neuem Teich und anschließend durch Gießen, um dann in die Lahn einzumünden. Im Gießener Lahntal kommen weitere Baggerseen vor (Abb. 2).
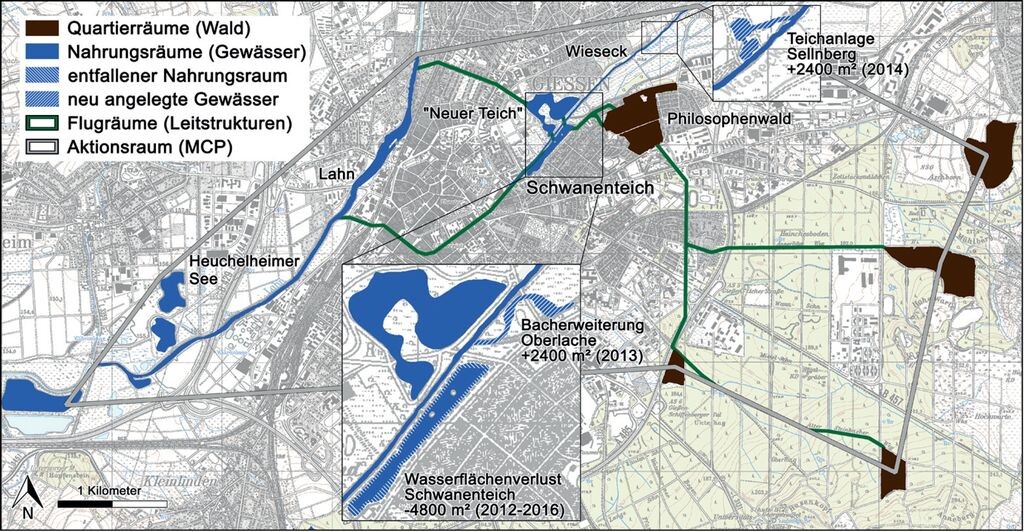
Neben dem temporären Nahrungsraumverlust (09/11 bis 02/12) sollte eine landschaftspflegerische Umgestaltung des Schwanenteichs gleichzeitig auch die Lebensbedingungen für den nachgewiesenen Bitterling (Rhodeus amarus) ökologisch durch die Bepflanzung mit einer Hochstaudenflur und Unterwasserbeeten verbessern (geplant für 2016). Gleichzeitig sollte eine Beobachtungsterrasse für Erholungssuchende im Jahr 2012 errichtet und das Teichufer zur Herstellung des ursprünglichen Verlaufs angepasst werden. Diese Maßnahmen würden langfristig zu einem permanenten Nahrungsraumverlust von ca. 12 % des Schwanenteichs (1600 m2 Hochstaudenflur + 1700 m2 Besucherterrasse + 1500 m2 Anpassung des Uferverlaufes = 4800 m2) führen (Abb. 2). Um diesen Verlust zu kompensieren, wurden neue Nahrungsräume in unmittelbarer Umgebung zum Schwanenteich angelegt. Dabei wurde auf eine Ausgestaltung entsprechend der artspezifischen Bedürfnisse der Wasserfledermaus geachtet.
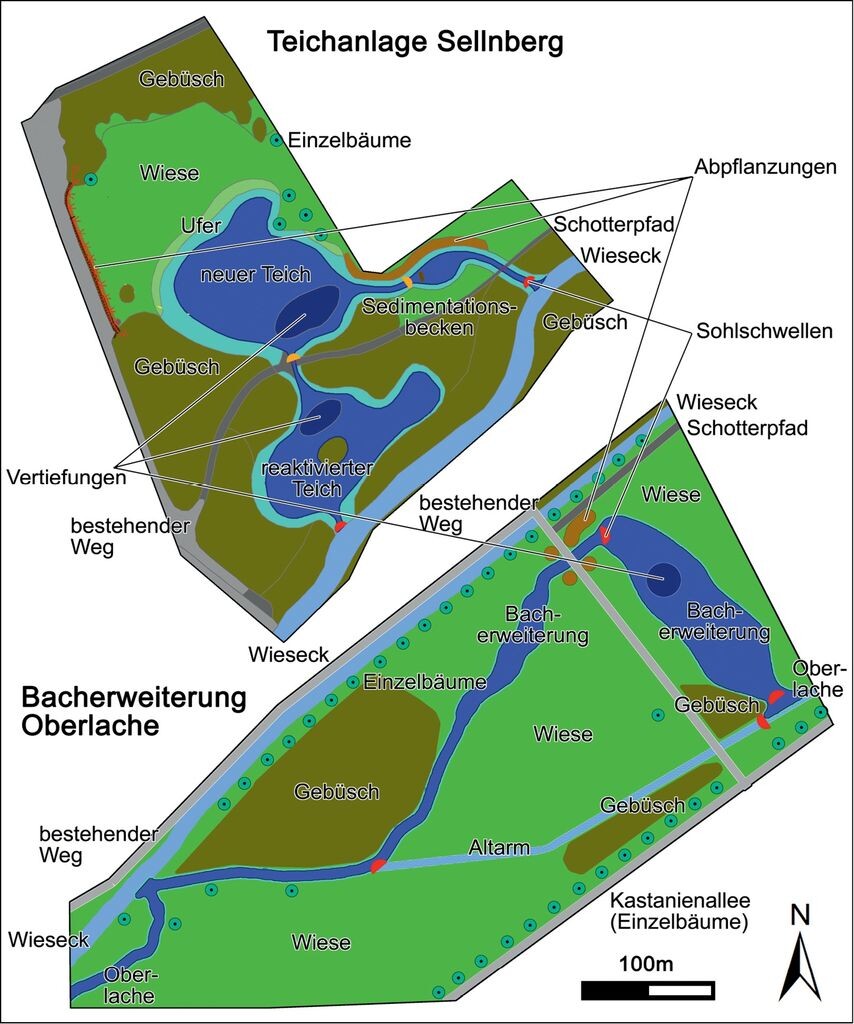
Im September 2013 stand der lokalen Wasserfledermaus-Population bereits eine zusätzliche Wasserfläche von ca. 2400 m2 durch die naturnahe Erweiterung eines Bachlaufs (Oberlache) als Nahrungsraum zur Verfügung (Abb. 2). Diese neue Wasserfläche wurde durch den dauerhaften Rückstau der Wieseck und eine Flachwasserzone mit nahezu stehendem Gewässer geschaffen. Der überwiegende Böschungsbereich wurde der natürlichen Sukzession überlassen, lediglich lokale Pflanzungen wurden vorgenommen. Vereinzelte Gruppen größerer Wasserbausteine bieten sowohl im Gewässer als auch an der Wasseroberfläche geschützte Lebens- und Nahrungshabitate (Abb. 3).
Im Jahr 2014 wurde ein weiteres Gewässer mit Frischwasser-Verbindung aus der Wieseck mit einer Ausdehnung von ca. 2400 m2 angelegt (Teichanlage Sellnberg), welches sich ca. 1000 m nördlich des Schwanenteichs in einer Entfernung von etwa 500m zum Philosophenwald befindet (Abb. 2). Dieses neue Gewässer verfügt über zwei Flachwasserteiche sowie einen Sedimentationsteich, welche über Gräben untereinander und mit der Wieseck verbunden sind. Integriert in das Vorhaben war neben der Neuanlage auch die Reaktivierung eines bereits vorhandenen, aber stark verlandeten Teiches, der weitgehend seine ökologische Funktion verloren hatte. Die beiden Teiche wurden mit verschiedenen Wassertiefen und Uferzonen angelegt. Flache Uferbereiche, kleinere Steinaufschüttungen sowie inselartige oder terrassierte Uferbereiche bieten verschiedenen Pflanzen- und Tierarten eine Vielzahl an Habitat- und Lebensraumstrukturen (Abb. 3). Somit stehen der lokalen Wasserfledermauspopulation im Jahr 2015 etwa 4800 m2 neuer Nahrungsraum zur Verfügung (Abb. 2).
In einer Entfernung von ca. 300m zum Schwanenteich befindet sich der Philosophenwald. Dieser ist als Bannwald ausgewiesen und wird als Naherholungsgebiet genutzt. Er besitzt eine Fläche von 28 ha, die Bäume sind im Durchschnitt 170 Jahre alt. Der Philosophenwald hat Inselcharakter und ist von den nächsten Waldbereichen etwa 1km entfernt (Abb. 2). Wochenstubenkolonien der Wasserfledermaus, des Kleinabendseglers (Nyctalus leisleri), der Mückenfledermaus (Pipistrellus pygmaeus) sowie Vorkommen der Rauhautfledermaus (Pipistrellus nathusii) und Fransenfledermaus (Myotis nattereri) nutzen vorkommende Baumhöhlen als Tagesquartier. Außerdem wurde der Große Abendsegler (Nyctalus noctula) als Wochenstuben- und Winterschlafgesellschaft nachgewiesen (eigene Daten; Hessen Forst FENA 2011, Roswag et al. 2014).
3 Untersuchungsart
Die hessischen Reproduktionsnachweise der Wasserfledermaus befinden sich überwiegend in Flusstälern wie dem Lahntal (Hessen Forst FENA 2003, 2006). In der Roten Liste wird sie als in Hessen gefährdet, auf nationaler Ebene als ungefährdet aufgeführt (Meinig et al. 2009). Die Wasserfledermaus bewohnt vorwiegend Baumhöhlen. Sie ist häufig auf gleichmäßigen Bahnen über ruhigen Wasseroberflächen zu beobachten, wobei sie vorwiegend Zuckmücken (Chironomiden) erbeutet (Dietz et al. 2007, Encarnação et al. 2005, 2012; Roer & Schober 2001). Die Dauer der Jagdflüge variiert je nach Reproduktionsstatus (Dietz & Kalko 2007, Encarnação et al. 2006).
Über die Lebensraumnutzung der lokalen Wasserfledermauspopulation in Gießen liegen aus verschiedenen Untersuchungen Informationen vor (Dietz et al. 2006, Dietz & Kalko 2007, Encarnação et al. 2005, Heimann 2001, Weise 1998, eigene Studien) und eine arttypische Habitatnutzung mit Baumquartieren in Wäldern, linienförmigen Landschaftselementen als Flugwegen und Gewässern als Nahrungsräume ist dokumentiert. Als zentrale Lebensraumelemente wurden der Philosophenwald (Quartierraum) mit den zwei nahegelegenen Stadtteichen in der Wieseckaue und die Lahn als Nahrungsräume identifiziert. In größerer Distanz liegende Quartier- und Nahrungsräume werden in geringerem Ausmaß aufgesucht. Es liegen trotz umfangreicher Telemetrie- und Fang-Wiederfangstudien keine Hinweise über einen größeren als den dargestellten Aktionsraum vor (Abb. 2).
Im Mai etablieren sich die Wochenstuben von Myotis daubentonii mit zumeist 20 bis 50 Weibchen in den Sommerlebensräumen. Während der Wochenstubenzeit leben die Männchen in unmittelbarer Nachbarschaft oder größerer Entfernung von den Weibchen. Die arttypischen Reproduktionsphasen (Dietz et al. 2007, Roer & Schober 2001) können für die Populationen im Landkreis Gießen bestätigt werden (Encarnação et al. 2005). Über eine Analyse der Körpergewichte und Fortpflanzungsbereitschaft von Alttieren (Weibchen n=780; Männchen n=1802) bzw. das Auftreten selbstständiger Jungtiere (n=579) der Jahre 1997 bis 2013 kann die Schwangerschaft (Gravidität) bis Anfang Juni (Kalendertag (KT) 160) bzw. die Stillzeit (Laktation) der adulten Weibchen (zwischen Ende Mai und Mitte Juli, KT 145-200), der Geburtszeitraum der Jungtiere (KT 145-160) sowie die einsetzende Fortpflanzungsbereitschaft (Spermatogenese) der Männchen (KT 170-260) zeitlich eingegrenzt werden. Im August löst sich die Wochenstubengemeinschaft auf und es bilden sich Paarungsgruppen in den Sommerlebensräumen (ab KT 220) (Encarnação 2012a, b; Encarnação et al. 2004a).
Eine Abgrenzung energieaufwendiger Reproduktionsphasen ist zur Einschätzung potenzieller Beeinträchtigungen lokaler Fledermauspopulationen durch Nahrungsraumverlust erforderlich, da dadurch die Nahrungsverfügbarkeit direkt beeinflusst wird. Dabei weisen die einzelnen demographischen Einheiten unterschiedliche Zeiträume mit erhöhtem Nahrungsbedarf auf. Je nach Klima und Nahrungsverfügbarkeit kann sich der Geburtszeitraum um ein bis zwei Wochen verschieben. Weibchen haben während Gravidität und Laktation, adulte Männchen während Spermatogenese und Paarungszeit einen hohen Nahrungsbedarf (Becker et al. 2013, Dietz & Kalko 2007, Encarnação & Dietz 2006). Jungtiere müssen wachstumsbedingt jederzeit ausreichend Nahrung vorfinden (Encarnação et al. 2004b) und in naheliegenden Nahrungsräumen ihre Jagdfertigkeiten erlernen. Junge Wasserfledermäuse unternehmen ab dem 21. Lebenstag erste Flugversuche, sind ab dem 25. Lebenstag flugfähig und haben ab dem 31. Lebenstag ein permanentes Gebiss (Roer & Schober 2001), wonach die Mutter die Milchversorgung des Jungtiers einstellt. Daher kann davon ausgegangen werden, dass ab dem 31. Lebenstag die Jungtiere vollständig autark sind und alle erforderlichen Fertigkeiten und räumlichen Kenntnisse zur Nahrungsaufnahme erlernt haben.
4 Material und Methoden
4.1 Untersuchungsphasen
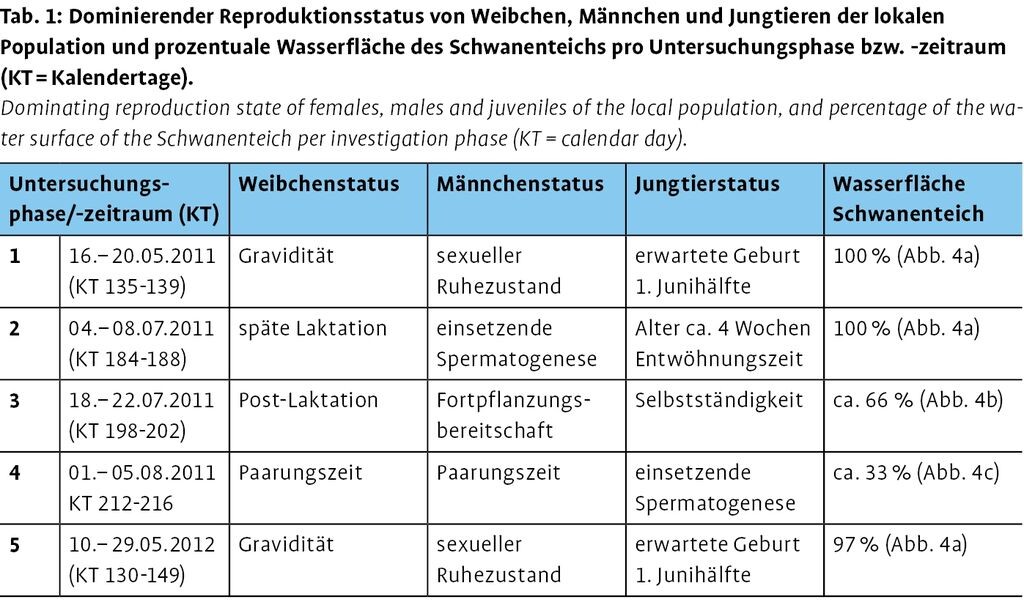
Bedingt durch die maßnahmenorientierten Fragestellungen und den spezifischen Reproduktionszyklus wurden fünf Untersuchungsphasen (1: 16.-20.05.2011 [Kalendertag (KT) 135-139], 2: 04.-08.07.2011 [KT 184-188], 3: 18.-22.07.2011 [KT 198-202], 4: 01.-05.08.2011 [KT 212-216], 5: 10.-29.05.2012 [KT 130-149]) anhand des dominierenden Reproduktionsstatus von Männchen und Weibchen bzw. dem Entwicklungsstand der Jungtiere festgelegt (Tab. 1). In diesen wurde jeweils eine Methodenkombination aus Bestandserfassung an Flugrouten, Aktivitätserfassung an Nahrungsräumen und Telemetrie bei trockener Witterung und Lufttemperaturen über 10°C angewandt.
4.2 Bestandserfassung an Flugroute
Zur standardisierten Erfassung der Bestandsgröße wurde an einer einsehbaren Stelle der Flugroute ein Beobachtungsstandort festgelegt, der vor dem artspezifischen Aktivitätsbeginn von zwei bis vier Beobachtern aufgesucht wurde. Telemetrische Studien belegen, dass nahezu alle Tiere den Beobachtungsstandort passieren. Mittels Detektorerfassung (Pettersson, D200) und Sichtbeobachtung wurden alle vom Philosophenwald Richtung Schwanenteich fliegenden Wasserfledermäuse für 45min ab dem ersten gesichteten Tier an fünf aufeinanderfolgenden Abenden gezählt. Tiere, welche in die entgegengesetzte Richtung flogen, wurden von der Summe abgezogen.
4.3 Aktivitätserfassung in Nahrungsräumen
Die Erfassung der Aktivität von Wasserfledermäusen am Schwanenteich und Neuen Teich erfolgte durch standardisierte Zählung mittels Halogenschweinwerfer (Conrad, 12V, 10W). Elf Beobachtungsstandorte (Schwanenteich n=6, Neuer Teich n=5) wurden hinsichtlich einer möglichst großen Abdeckung der Wasseroberfläche mit möglichst geringen Überschneidungsbereichen ausgewählt, ca. 60 min nach Aktivitätsbeginn an fünf aufeinanderfolgenden Abenden aufgesucht und zeitgleich die Wasseroberfläche dreimal in einem Abstand von jeweils 1min in einer Höhe von bis zu 1m in eine Richtung langsam abgeleuchtet. Alle den Lichtstrahl von links nach rechts durchquerenden Wasserfledermäuse wurden gezählt, in entgegengesetzter Richtung fliegende Individuen von der Summe abgezogen. Für die weitere Auswertung wurde der Mittelwert aus den drei Zählungen an einem Beobachtungsstandort verwendet, die Mittelwerte aller Standorte eines Gewässers (Schwanenteich oder Neuer Teich) summiert und der prozentuale Anteil erfasster Wasserfledermäuse pro Gewässer vom erfassten Gesamtbestand (Flugroute) ermittelt.
4.4 Radio-Telemetrie
Um die Raumnutzung zu analysieren, wurden Wasserfledermäuse mittels Japan-Netzen (Ecotone, 6-15m x 3-9m, 70 Denier) gefangen und während jeder Untersuchungsphase jeweils drei adulte Weibchen und Männchen mit einem minimalen Körpergewicht von 7,5g durch einen Transmitter markiert (Holohil, LB-2N, 0,35g). Die Befestigung des Senders erfolgte im Rückenfell mittels Hautkleber (Sauer, Typ 50.01) und hält ca. 48 Tage. In den Untersuchungsphasen 2, 3 und 4 wurden zusätzlich jeweils drei Jungtiere (≥ 7,5g Körpergewicht) mit Sendern versehen. Während vier Nächten erfolgte die Überwachung am Schwanenteich, Neuen Teich, an der Lahn und der Wieseck im Stadtpark durch drei Beobachtergruppen zwischen Sonnenuntergang und Sonnenaufgang mittels Telemetrie-Empfängern (Wagener, Yaesu VR500 mit HB9CV Yagi-Handantenne). Nicht anwesende Tiere wurden durch ein viertes Beobachterteam gesucht. Zur Auswertung der Gewässernutzung erfolgte eine prozentuale Präsenz-Absenz-Analyse pro Untersuchungsphase und Geschlechts- bzw. Altersgruppe, d.h. drei sendermarkierte Weibchen der Untersuchungsphase 1 mit einem Beobachtungszeitraum von vier Nächten ergeben in der Summe 12 Weibchennächte (100 %). Beispielsweise errechnet sich in der Untersuchungsphase 1 durch die Anwesenheit von nur einem Weibchen in drei Nächten am Heuchelheimer See ein Anwesenheits-Anteil von 25 % aller Weibchennächte (3 von 12 Nächten) für dieses Gewässer. Gleichzeitig ergibt sich für die Lahn ein Anwesenheits-Anteil von 100 % aller Weibchennächte durch den Besuch aller drei Weibchen in allen vier Nächten (12 von 12 Nächten).
5 Ergebnisse
5.1 Populationsstatus und Wasserfläche während der Untersuchungsphasen
Während der Untersuchungsphase 1 ließen die Konstitution der graviden Weibchen und die klimatischen Verhältnisse in den Vormonaten einen Geburtszeitraum in der ersten Junihälfte vermuten. In der Phase 2 befanden sich die Weibchen im Entwöhnungsprozess der Jungtiere, deren Konstitution den erwarteten Geburtszeitraum Anfang Juni bestätigte. Im Laufe der Phase 3 begann sich der Wochenstubenverband aufzulösen und die post-laktierenden Weibchen bildeten mit fortpflanzungsbereiten Männchen und den selbstständigen Jungtieren gemischte Gruppen. Die Untersuchungsphase 4 war gekennzeichnet durch eine Körpergewichtszunahme aller demographischen Einheiten und der einsetzenden Fortpflanzungsbereitschaft juveniler Männchen. Die Konstitution von Weibchen und Männchen und die klimatischen Verhältnisse während der Untersuchungsphasen 5 und 1 entsprachen sich (Tab. 1).
Nach der Untersuchungsphase 2 (Abb. 4a) wurde die Entleerung des Schwanenteichs eingeleitet, so dass die Wasseroberfläche in Phase 3 ca. 66 % (Abb. 4b) und Phase 4 ca. 33 % (Abb. 4c) betrug. Bis Ende September 2011 wurde das Wasser im Schwanenteich komplett abgelassen (0 %, Abb. 4d), um bis März 2012 wieder vollständig befüllt zu werden. Die Wasseroberfläche des Schwanenteichs betrug in Untersuchungsphase 5 durch das Baufeld der geplanten Terrasse ca. 97 % (Tab. 1). Die Bepflanzungen sind für das Jahr 2016 geplant.
5.2 Bestandserfassung an Flugroute
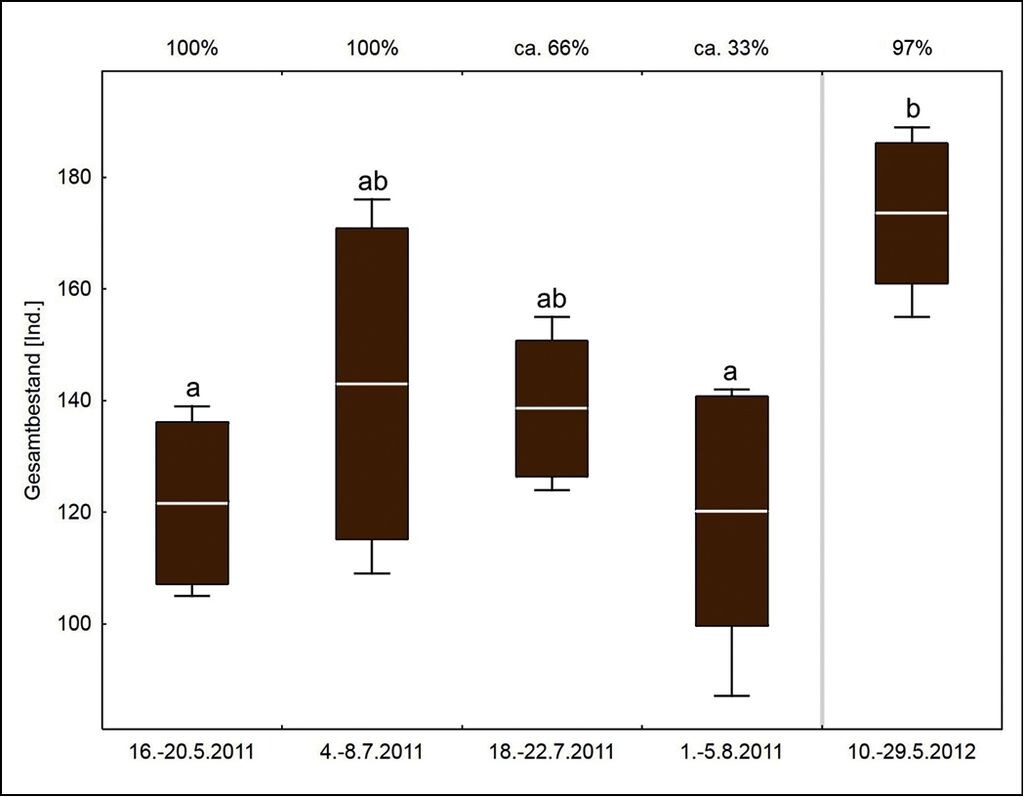
Der Gesamtbestand schwankte zwischen 87 und 189 Individuen (Abb. 5), die Daten entsprechen einer Normalverteilung (Kolmogrov-Smirnov und Lilliefors Tests: p>0,05). Zwischen den vier Untersuchungsphasen 2011 wurden keine signifikanten Unterschiede in der Bestandsgröße festgestellt. In den Untersuchungsphasen 1 (n=105 bis 139) und 4 (n=87 bis 142) im Jahr 2011 wurden signifikant weniger Wasserfledermäuse an der Flugroute erfasst als im Mai 2012 (Untersuchungsphase 5, n=155 bis 189, Tukey HSD-Test: p<0,01 für beide Vergleiche).
5.3 Aktivitätserfassung in naheliegenden Nahrungsräumen
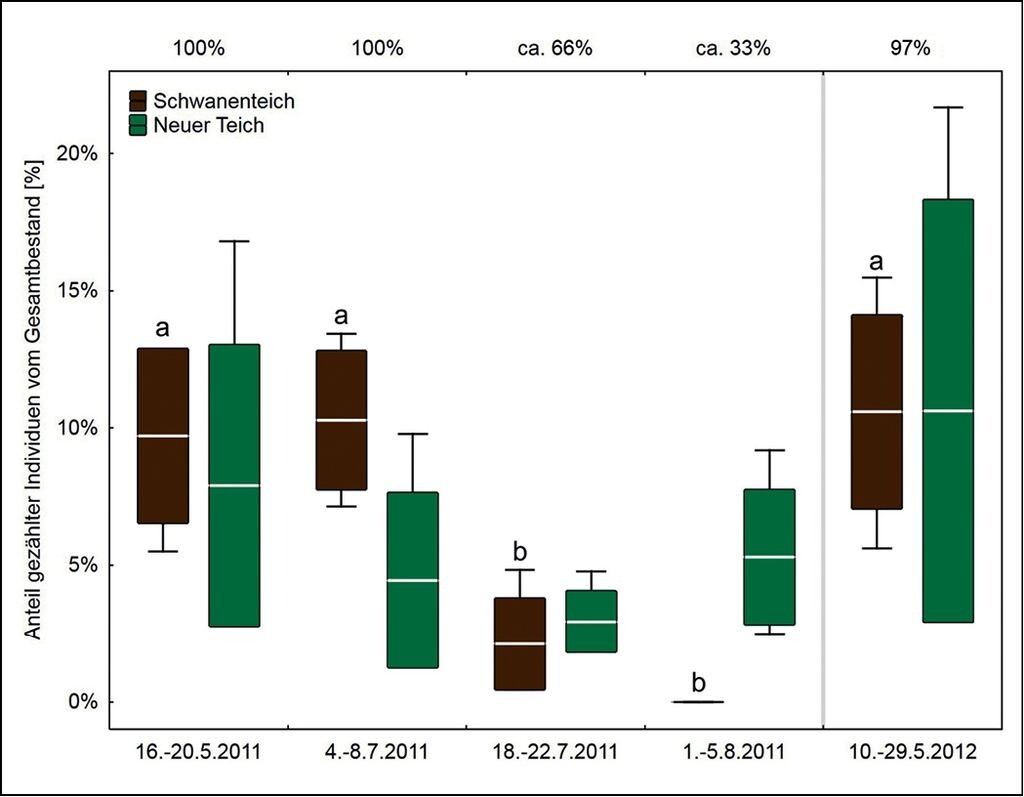
Die Aktivität jagender Wasserfledermäuse am Schwanenteich und Neuen Teich schwankte zwischen 2 und 37 % vom Gesamtbestand, die Daten entsprechen ebenfalls einer Normalverteilung (Kolmogrov-Smirnov und Lilliefors Tests: p>0,05). In der Untersuchungsphase 3 (0 bis 5 %) und 4 (0 %) während der Absenkung des Wasserstands wurden erwartungsgemäß signifikant weniger Wasserfledermäuse am Schwanenteich erfasst als in den Untersuchungsphasen 1 (6 bis 13 %), 2 (7 bis 13 %) und 5 (6 bis 15 %, Tukey HSD-Test: p<0,01 für alle Vergleiche) bei gefülltem Teich. Die Aktivität am Neuen Teich (1 bis 22 %), dessen Wasserspiegel nicht vom Eingriff betroffen war und ausschließlich natürlichen Schwankungen unterlag, unterschied sich nicht signifikant zwischen den fünf Untersuchungsphasen (Tukey HSD-Test: p>0,05 für alle Vergleiche) (Abb. 6).
5.4 Weitere Nahrungsraumnutzung im Aktionsraum
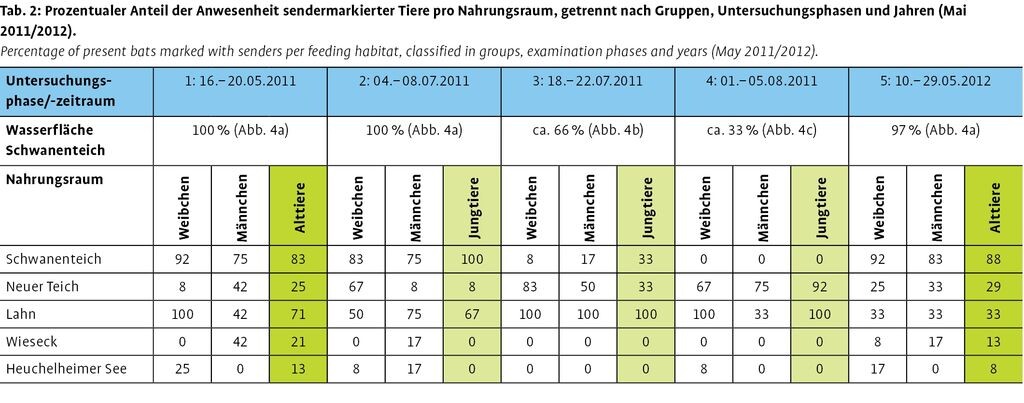
Die prozentuale Anwesenheit sendermarkierter Wasserfledermäuse in den verschiedenen Nahrungsräumen zeigt eine variierende Nutzung von fünf Nahrungsräumen (Abb. 2, Tab. 2).
In den Untersuchungsphasen 1 und 2 (100 % Wasseroberfläche) nutzten Weibchen in 83 % bzw. 92 % und Männchen in 75 % der Nächte den Schwanenteich als Nahrungsraum. Daneben wurden beide Gruppen ebenfalls am Neuen Teich, an der Lahn und am Heuchelheimer See beobachtet. Jagende Männchen konnten zusätzlich an der Wieseck erfasst werden. Jungtiere nutzten in Phase 2 zu 100 % der Nächte den Schwanenteich, aber ebenfalls den Neuen Teich (8 %) und die Lahn (67 %) als Nahrungsräume (Tab. 2). Daraufhin wurde eine langsame, reversible Absenkung des Wasserstands im Schwanenteich veranlasst.
In der Untersuchungsphase 3 (ca. 66 % Wasseroberfläche) jagten Weibchen in 8 % der Nächte, Männchen in 17 % der Nächte und Jungtiere in 33 % der Nächte am Schwanenteich. Daneben wurden alle drei Gruppen in 100 % der Nächte an der Lahn und ebenfalls am Neuen Teich beobachtet (Tab. 2). Auf dieser Grundlage wurde die langsame Absenkung des Wasserstands im Schwanenteich fortgesetzt.
In der Untersuchungsphase 4 (ca. 33 % Wasseroberfläche) nutzte keine der drei Gruppen den Schwanenteich als Nahrungsraum. Weibchen und Jungtiere wurden in 100 % der Nächte an der Lahn erfasst und jagten ebenfalls intensiv mit 67 % bzw. 92 % am Neuen Teich. Männchen zeigten eine hohe Mobilität und wurden ausschließlich am Neuen Teich (75 %) und an der Lahn (33 %) registriert (Tab. 2). Die langsame Absenkung des Wasserstands im Schwanenteich wurde weiter fortgesetzt.
In der Untersuchungsphase 5 (ca. 97 % Wasseroberfläche) konnten Weibchen in 92 % der Nächte und Männchen in 83 % der Nächte am Schwanenteich jagend beobachtet werden. Daneben wurden beide Gruppen am Neuen Teich und an der Lahn erfasst. Weibchen nutzten in dieser Phase zusätzlich den Heuchelheimer See, Männchen die Wieseck als Nahrungsraum (Tab. 2).
Zum Vergleich von Untersuchungsphase 1 vor Trockenlegung mit Phase 5 nach Trockenlegung wurden Männchen- und Weibchennächte (Alttiere) zusammengefasst. Es zeigt sich, dass sich die räumliche Nahrungsraumnutzung beider Jahre entspricht, wobei in 2012 der Schwanenteich und der Neue Teich häufiger, die Lahn, die Wieseck und der Heuchelheimer See tendenziell seltener besucht wurden (Tab. 2).
Im September 2013 konnte die Nutzung der neu geschaffenen Oberlache als Nahrungsraum durch Myotis daubentonii festgestellt werden. Im August 2014 wurden jagende Wasserfledermäuse auch an der Teichanlage Sellnberg beobachtet. Beide neu angelegten potenziellen Nahrungsräume wurden demzufolge bereits unmittelbar im Jahr ihrer Fertigstellung von der Wasserfledermauspopulation angenommen.
Als Erfolgskontrolle wurde am 12.08. 2015 der prozentuale Anteil jagender Wasserfledermäuse vom Gesamtbestand an der Oberlache und der Teichanlage Sellnberg im Vergleich zum nördlichen Ende des Schwanenteichs bestimmt. Der Schwanenteich stellte mit 8,5 % nach wie vor den dominanten Nahrungsraum der lokalen Population dar, beide neu angelegten Gewässer wurden aber ebenfalls regelmäßig als Nahrungsraum genutzt (Oberlache: 1,5 %, Sellnberg: 4 %).
5.5 Erheblichkeitsabschätzung
5.5.1 Nahrungsraumnutzung von Alt- und Jungtieren der lokalen Wasserfledermauspopulation
Es konnte gezeigt werden, dass in Untersuchungsphase 1 als auch 2 Alttiere beider Geschlechter mehrere Nahrungsräume nutzen. Durch den Bestandsanstieg und die Konstitution von Weibchen bzw. Jungtieren konnte der Geburtszeitraum auf Anfang Juni 2011 geschätzt werden. In der Untersuchungsphase 2 hatten die Jungtiere ein geschätztes Alter von 30 bis 40 Lebenstagen und waren seit etwa fünf bis 15 Tagen flugfähig. Die beobachtete Nahrungsraumnutzung der Jungtiere in dieser Phase bestätigte eine fortgeschrittene Selbstständigkeit. Die Lahn stellte ihren bevorzugten Nahrungsraum (67 %) dar. Diese Beobachtung wurde durch den fehlenden Aktivitätsanstieg am Neuen Teich gestützt.
5.5.2 Reaktion von Alt- und Jungtieren auf den temporären Nahrungsraumverlust
Nachdem von einer Kompensation des temporären Nahrungsraumverlusts aller demographischen Einheiten ausgegangen werden konnte, wurde eine langsame, jederzeit reversible Leerung des Schwanenteichs ab Mitte Juli 2011 veranlasst und die Reaktion der lokalen Population überwacht. Der Gesamtbestand verringerte sich nicht signifikant in den Untersuchungsphasen 3 und 4 während der Absenkung des Wasserstands des Schwanenteichs (Abb. 5), allerdings verringerte sich erwartungsgemäß die Aktivität jagender Wasserfledermäuse. Am Neuen Teich konnte jedoch kein gleichzeitiger Anstieg der Aktivität verzeichnet werden (Abb. 6). Die Nahrungsraumnutzung der sendermarkierten Tiere zeigte, dass die Lahn und der Neue Teich von allen demographischen Einheiten weiterhin genutzt wurden (Tab. 2). Daher ist zu vermuten, dass insbesondere die Lahn während der Untersuchungsphasen 3 und 4 den Hauptnahrungsraum für die gesamte lokale Population darstellt.
6 Diskussion
Zur Umsetzung der Sanierungsmaßnahmen und landschaftsplanerischen Umgestaltung des Schwanenteichs war es erforderlich, in der frostfreien Periode den Teich über drei Monate trocken zu legen. Nur auf diese Weise konnte ca. 14000m³ Bodenschlamm abgetragen werden. Allerdings kollidierte dieser Zeitraum mit der saisonalen Aktivitätszeit der lokalen Wasserfledermauspopulation im Gießener Philosophenwald, von der eine intensive Nutzung des Schwanenteichs als Nahrungsraum bekannt war. Eine vorlaufende Erheblichkeitsabschätzung konnte mangels Detailinformationen eine Beeinträchtigung der lokalen Population nicht ausschließen, wenn einzelnen demographischen Einheiten in energetisch anspruchsvollen Phasen durch den temporären Nahrungsraumverlust nicht genügend Nahrung zur Verfügung stünde. Um diesem Risiko Sorge zu tragen, wurde eine wissenschaftliche Begleitung der Maßnahme durchgeführt, welche die Machbarkeit überprüfen, den Zeitraum der Durchführung festlegen und die Reaktion der lokalen Population überwachen sollte. Auf Grundlage der dabei ermittelten Ergebnisse können die eingangs dargestellten Fragen beantwortet werden.
Welche Nahrungsräume werden in welchem Umfang von Alt- und Jungtieren der lokalen Wasserfledermaus-Population genutzt?
Eine detaillierte Differenzierung, für welche demographische Einheit welcher Nahrungsraum zu welchem Zeitpunkt eine dominierende Funktion übernimmt, lag nicht vor. Daher war es erforderlich, die Nutzung auf Grundlage der saisonal schwankenden energetischen Ansprüche der demographischen Einheiten separat zu analysieren. In dieser Studie konnte eine differenzierte Nahrungsraumnutzung aller demographischen Einheiten ermittelt und gezeigt werden, dass Alttiere beider Geschlechter mehrere Nahrungsräume intensiv in Untersuchungsphase 1 als auch 2 nutzen. Diese Nutzung hängt vermutlich vom saisonal bedingten Insektenvorkommen ab. In der Untersuchungsphase 2 zeigten die Jungtiere bereits eine fortgeschrittene Selbstständigkeit, die mit der Nutzung von entfernt gelegeneren Nahrungshabitaten einherging. In den Untersuchungsphasen 3 und 4 nutzten die Jungtiere einen den Alttieren entsprechenden Aktionsraum.
Kann der temporäre Verlust nahegelegener Nahrungsräume durch die intensivierte Nutzung alternativer Nahrungsräume kompensiert werden?
Nachdem sichergestellt war, dass allen demographischen Einheiten alternative Nahrungsräume bekannt waren, konnte davon ausgegangen werden, dass sie einen temporären Nahrungsraumverlust ausgleichen können. Allerdings war dies vermutlich vor Untersuchungsphase 2 nicht gegeben. Durch die artspezifische Ernährungsweise von Wasserfledermäusen sind diese zur erfolgreichen Jagd unmittelbar an Gewässer mit ruhiger Wasseroberfläche angewiesen. Zu Beginn ihrer Selbstständigkeit müssen Jungtiere ihre Jagdfertigkeiten in naheliegenden Nahrungsräumen von ihren Müttern erlernen und können zu dieser Zeit einen Nahrungsraumverlust an solchen Gewässern vermutlich kaum kompensieren. Daher müssen in Nahrungsräumen in unmittelbarer Nähe zu Wochenstubenkolonien Eingriffe in diesem Zeitraum vermieden werden, um die lokale Population nicht zu beeinträchtigen.
Kommt es zu einer Änderung der Habitatnutzung der lokalen Population als Reaktion auf den temporären Nahrungsraumverlust?
Es wurde eine Änderung der Habitatnutzung während der Maßnahme verzeichnet, die allerdings nicht ausschließlich als Reaktion auf die Absenkung des Wasserstands, sondern ebenfalls auf natürliche saisonale Veränderungen der Nahrungsverfügbarkeit an Gewässern zurückgeführt werden kann (Encarnação et al. 2010). Weder die Ergebnisse der Flugroutenzählungen noch die im Rahmen einer Langzeitstudie durchgeführte Fang-Markierung-Wiederfang-Analyse weisen auf eine maßnahmenbedingte Dismigration hin. Die Habitatnutzung der lokalen Population in Untersuchungsphase 5 nach dem temporären Nahrungsraumverlust entsprach der in Untersuchungsphase 1 davor.
Kann permanenter Nahrungsraumverlust unmittelbar durch die Anlage neuer Nahrungsräume kompensiert werden?
Unsere Ergebnisse zeigen, dass bereits im Jahr ihrer Fertigstellung und vor der geplanten Bepflanzung des Schwanenteichs in 2016 alle neu angelegten Gewässer die ökologische Funktion als Nahrungsraum für die lokale Wasserfledermauspopulation übernommen haben. Dies ist insbesondere darauf zurückzuführen, dass die neuen Nahrungsräume innerhalb des bestehenden Aktionsraums und vorlaufend zu den landschaftspflegerischen Eingriffen angelegt wurden. Auf diese Weise konnte die lokale Population diese Alternativen vor dem permanenten Nahrungsraumverlust erkunden und später intensivierter nutzen.
Die wiederhergestellte Aktivität jagender Tiere am Schwanenteich (2012 bis 2015), der Bestandsanstieg (2012), die umgehende Nutzung neu angelegter Gewässer (2013/14), der unveränderte Aktionsraum und eine Verminderung der Schwermetallkonzentration in Fledermaushaaren (2013, Flache et al. 2015a, b) sprechen für verbesserte Lebensbedingungen der lokalen Population trotz temporärem und permanenten Nahrungsraumverlust.
Welche Konsequenzen ergeben sich für die Stadt- und Landschaftsplanung?
Erhebliche Beeinträchtigungen lokaler Populationen geschützter Arten müssen vermieden und jegliche Beeinträchtigungen durch entsprechende Vermeidungs- und Kompensationsmaßnahmen minimiert werden. Fledermäuse sind streng geschützt und weisen eine hohe landschaftsplanerische Relevanz auf. Ihre Lebensweise führt durch die Nutzung unterschiedlicher Teillebensräume (Quartierräume, Nahrungsräume, Flugräume, Winterlebensräume) dazu, dass in der Stadt- und Landschaftsplanung ein Konfliktfeld zwischen Maßnahmenplanung und Fledermausvorkommen entstehen kann. Quartierräume können z.B. durch Eingriffe im Wald oder im Siedlungsbereich betroffen sein, Flugräume durch landschaftspflegerische Eingriffe oder Straßenbau beeinträchtigt werden.
Auch die Nahrungsräume von Fledermäusen machen dabei keine Ausnahme. Für Fledermäuse sind naturnahe und insektenreiche Landschaftsteile geeignete Nahrungsräume seien es Gewässer oder z.B. Wälder mit hohem Laubbaumanteil oder Streuobstwiesen. Der Untersuchungsumfang sollte über das Standardprogramm artenschutzrechtlicher Prüfungen hinausgehen, wenn es sich um Hauptnahrungsräume lokaler Populationen handelt, aber Detailinformationen über deren Nutzung fehlen und somit eine erhebliche Beeinträchtigung durch geplante Eingriffe nicht ausgeschlossen werden kann. Die variablen Aktivitätsphasen, die artspezifischen Jagdstrategien und der geschlechts- bzw. altersspezifische Nahrungsbedarf führen dazu, dass dann gezielte maßnahmenorientierte Erheblichkeitsabschätzungen und Kompensationen auf Grundlage gebietsspezifischer Informationen zu den lokalen Populationen in Verbindung mit einem begleitenden Monitoring erforderlich sind.
Im Mittelpunkt der Maßnahmenplanungen in Nahrungsräumen muss die Sicherstellung der Nahrungsverfügbarkeit für die einzelnen demographischen Einheiten während energetisch anspruchsvoller Reproduktions- bzw. Wachstumsphasen und eine Kompensation verlorener Nahrungsräume stehen. Auf dieser Grundlage ist es möglich, das Konfliktfeld zwischen Maßnahmenplanung und Fledermausvorkommen aufzulösen und die Lebensbedingungen lokaler Populationen letztendlich zu verbessern.
Dank
Unser Dank gilt den Mitgliedern und Studierenden der Arbeitsgruppe Säugetierökologie an der Universität Gießen für ihre tatkräftige Unterstützung bei der Datenaufnahme. Außerdem bedanken wir uns bei den Mitgliedern der Sportangler Gießen und Umgebung e.V. für ihre Mitarbeit in diesem Projekt.
Literatur
Becker, N.I., Tschapka, M., Kalko, E.K.V., Encarnação, J.A. (2013): Balancing the energy budget in free-ranging male Myotis daubentonii bats. Physiol. Biochem. Zool. 86 (3), 361-369. doi: 134.176.155.219.
Dietz, C., Helversen, O., Nill, D. (2007): Handbuch der Fledermäuse Europas und Nordwestafrikas. Franckh Kosmos, Stuttgart, 1. Aufl.
Dietz, M., Encarnação, J.A., Kalko, E.K.V. (2006): Small scale distribution patterns of female and male Daubentons bats (Myotis daubentonii). Acta Chiropt. 8 (2), 403-415.
, Kalko, E.K.V. (2007): Reproduction affects flight activity in female and male Daubentons bats, Myotis daubentoni. Can. J. Zool. 85 (5), 653-664.
Encarnação, J.A. (2012a): Mating at summer sites: indications from parentage analysis and roosting behaviour of Daubentons bats (Myotis daubentonii). Conserv. Genet. 13, 1161-1165. doi: 10.1007/s10592-012-0343-0.
(2012b): Spatiotemporal pattern of local sexual segregation in a tree-dwelling temperate bat Myotis daubentonii. J. Ethol. 30 (2), 271-278. doi: 10.1007/s10164-011-0323-8.
, Becker, N. I., Ekschmitt, K. (2010): When do Daubentons bats (Myotis daubentonii) fly far for dinner? Can. J. Zool. 88, 1192-1201.
, Dietz, M. (2006): Estimation of food intake and ingested energy in Daubentons bats (Myotis daubentonii) during pregnancy and spermatogenesis. Eur. J. Wildl. Res. 52 (4), 221-227.
, Dietz, M., Kierdorf, U. (2004a): Reproductive condition and activity pattern of male Daubentons bats (Myotis daubentonii) in the summer habitat. Mamm. Biol. 69 (3), 163-172.
, Dietz, M., Kierdorf, U., Wolters, V. (2004b): Body mass changes in male Daubentons bats Myotis daubentonii (Chiroptera, Vespertilionidae) during the seasonal activity period. Mammalia 68 (4), 291-297.
, Kierdorf, U., Holweg, D., Jasnoch, U., Wolters, V. (2005): Sex-related differences in roost-site selection by Daubentons bats Myotis daubentonii during the nursery period. Mammal. Rev. 35 (3-4), 285-294.
, Kierdorf, U., Wolters, V. (2006): Seasonal variation in nocturnal activity of male Daubentons bats, Myotis daubentonii (Chiroptera: Vespertilionidae). Folia Zool. 55 (3), 237-246.
Flache, L., Czarnecki, S., Düring, R.-A., Kierdorf, U., Encarnação, J.A. (2015a): Trace metal concentrations in hairs of three bat species from an urbanized area in Germany. Journal of Environmental Sciences 31, 184-193.
, Ekschmitt, K., Kierdorf, U., Czarnecki, S., Düring, R.-A., Encarnação, J.A. (2015b): Reduction of metal exposure of Daubentons bats (Myotis daubentonii) following remediation of pond sediment as evidenced by metal concentrations in hair. Science of The Total Environment 547, 182-189.
Heimann, H. (2001): Untersuchungen zur Territorialität bei Wasserfledermäusen (Myotis daubentonii Kuhl, 1819) im Jagdgebiet. Unveröff. Dipl.-Arb., Justus-Liebig-Univ. Gießen.
Hessen Forst FENA (2003): Artgutachten zur gesamthessischen Situation der Wasserfledermaus Myotis daubentonii: Verbreitung, Kenntnisstand, Gefährdung. Gießen.
(2006): Artensteckbrief Wasserfledermaus (Myotis daubentonii). Gießen.
(2011): Artgutachten 2011 Großer Abendsegler: Bundesstichprobenmonitoring 2011 von Fledermausarten (Chiroptera) in Hessen. Gießen.
Ineichen, S., Ruckstuhl, M., Klausnitzer, B. (2012): Stadtfauna: 600 Tierarten unserer Städte. Haupt, Bern.
Meinig, H., Boye, P., Hutterer, R. (2009): Rote Liste und Gesamtartenliste der Säugetiere (Mammalia) Deutschlands. In: Haupt, H., Ludwig, G., Grunke, H., Binot-Hafke, M., Otto, C., Pauly, A., Hrsg., Rote Liste gefährdeter Tiere, Pflanzen und Pilze Deutschlands, Band 1: Wirbeltiere. Bundesamt für Naturschutz, Hrsg., Bonn, 115-158.
Roer, H., Schober, W. (2001): Myotis daubentonii (Leisler, 1819) Wasserfledermaus. In: Krapp, F., Hrsg., Handbuch der Säugetiere Europas, Bd. 4, Aula, Wiebelsheim, 257-280.
Roswag, A., Becker, N.I., Encarnação, J.A. (2014): Factors influencing stable nitrogen isotope ratios in wing membranes of insectivorous bat species: A field study. Mammalian Biology 79 (2), 110-116.
Simon, M., Hüttenbügel, S., Smit-Viergutz, J. (2004): Ökologie und Schutz von Fledermäusen in Dörfern und Städten. Schr.-R. Landschaftspfl. Naturschutz 76.
Weise, C. (1998): Untersuchungen zur Lebensraumnutzung männlicher Wasserfledermäuse Myotis daubentonii, Kuhl 1819. Unveröff. Dipl.-Arb., Justus-Liebig-Univ. Gießen.
Zucchi, H. (1999): Naturschutz/Landschaftspflege/Artenschutz/Biotopschutz: Definitionen, Abgrenzungen. In: Konold, W., Böcker, R., Hampicke, U., Hrsg., Handbuch Naturschutz und Landschaftspflege: Kompendium zu Schutz und Entwicklung von Lebensräumen und Landschaften. Wiley-VCH, Weinheim.
-
Abb. 1: Wasserfledermaus (Myotis daubentonii) bei einer arttypischen Beutefangbewegung. Sie ist in der Lage, mit durchschnittlich sieben Fanghandlungen pro Minute und einem Fangerfolg von über 88 % bis zu 3000 Zuckmücken in einer Nacht zu erbeuten (Becker et al. 2013)© Marko KönigDaubenton’s bat (Myotis daubentonii) during a typical movement to catch prey. With an average of seven catches per minute and a catching success of 88 % the animal is able to prey up to 3.000 chironomids per night (Becker et al. 2013). -
Abb. 2: Lebensraumnutzung der lokalen Wasserfledermaus-Population in Gießen. Den dominanten Quartierraum stellt der Philosophenwald dar, dominante Nahrungsräume sind der Schwanenteich und der „Neue Teich“ in ca. 300m westlicher Entfernung zum Philosophenwald sowie die ca. 1200m vom Schwanenteich entfernte Lahn. Ausschnitte: maßnahmenbedingter Verlust von Nahrungsräumen und neu angelegte Gewässer als Kompensationsmaßnahmen. Habitat use of the local population of Daubenton’s bat in the town of Gießen. Their dominant roosting area is situated in the forest “Philosophenwald”. In a distance of 300 m they have their dominant feeding habitats, the “Schwanenteich” and the “Neuer Teich”, and additionally the river “Lahn” in a distance of 1,200 m from the Schwanenteich. Sections: loss of feeding habitats due to the measure; newly constructed water body as compensation measure. -
Abb. 3: Struktur neu angelegter Gewässer. Durch die Erweiterung der Oberlache bzw. die Neuanlage und Reaktivierung einer Teichanlage entstanden 4800m² neue Wasserfläche als Nahrungsraum für Wasserfledermäuse.Structure of newly constructed water bodies. By extending the “Oberlache” and reactivating existing ponds 4,800 m2 of new water surface could be created as foraging habitat for Daubenton’s bats. -
Abb. 4a-d: Wasserstand des Schwanenteichs in den verschiedenen Untersuchungsphasen. a) In den Untersuchungsphasen 1, 2 und 5 betrug die Wasseroberfläche zwischen 97-100 %, b) in Phase 3 ca. 66 % und c) in Phase 4 ca. 33 %. Ab Mitte Juli 2011 wurde das Wasser bis zur Trockenlegung abgelassen (d, 0 % von 9/2011 bis 2/2012). Im März 2012 wurde der Schwanenteich wieder befüllt.Water level of the Schwanenteich during different phases of the study. a) During the phases 1, 2, and 5 the water surface covered between 97 and 100 %, b) during phase 3 about 66 % were covered, and c) in phase 4 only about 33 %. Starting middle of June 2011 the pond was drained (d, 0 % from 09/11 to 2/12). In March 2012 the Schwanenteich was flooded again. -
Abb.5: Gesamtbestand der lokalen Wasserfledermauspopulation (Individuen pro 45min). Der Untersuchungszeitraum ist auf der unteren, die Wasseroberfläche des Schwanenteichs auf der oberen x-Achse für alle fünf Untersuchungsphasen angegeben. Signifikante Unterschiede zwischen den Phasen (Tukey HSD-Test: p < 0,01) sind durch unterschiedliche Buchstaben gekennzeichnet. Dargestellt sind jeweils der Mittelwert (Strich), ± Standardabweichung (Box) und Streuungsweite (Antennen).Complete local population of Daubenton’s bat (individuals per 45 min). The investigation period is shown on the lower x-axis, the water surface of the Schwanensee on the upper x-axis. Significant differences between the phases (Tukey HS test: p<0.01) are marked by different letters. The illustration shows the mean value (line), ± standard deviation (box) and distribution (antennas). -
Abb. 6: Prozentualer Anteil jagender Wasserfledermäuse vom Gesamtbestand am Schwanenteich und Neuen Teich. Der Untersuchungszeitraum ist auf der unteren, die Wasseroberfläche des Schwanenteichs auf der oberen x-Achse für alle fünf Untersuchungsphasen angegeben. Signifikante Unterschiede zwischen den Phasen (Tukey HSD-Test: p < 0,01) sind durch unterschiedliche Buchstaben gekennzeichnet. Dargestellt sind jeweils der Mittelwert (Strich) ± Standardabweichung (Box) und Streuungsweite (Antennen).Percentage of foraging bats compared to the total stock at Schwanenteich and Neuer Teich. The lower x-axis shows the examination period, the upper x-axis depicts the water surface for all five phases investigated. Significant differences between the phases (Tukey HSD test: p < 0.01) are marked by different letters. The illustration shows the mean value (line), ± standard deviation (box) and distribution (antennas). -
Temporärer und permanenter Nahrungsraumverlust für Fledermäuse -
Temporärer und permanenter Nahrungsraumverlust für Fledermäuse -
Temporärer und permanenter Nahrungsraumverlust für Fledermäuse



Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.