Auswirkungen des rezenten Klimawandels auf die Biodiversität in Mittelgebirgen
Abstracts
Für montan verbreitete Arten und Habitate wird die Gefährdung durch den Klimawandel als besonders hoch eingestuft, so dass dringender Handlungsbedarf für den Naturschutz besteht. Die vorliegende Literaturstudie fasst die klimawandelbedingten Auswirkungen auf Arten und Lebensräume in Mittelgebirgen mit besonderem Fokus auf dem Hochsauerland (Nordrhein-Westfalen) zusammen.
Eine der bedeutendsten Gefährdungsursachen durch den Klimawandel ist die Erwärmung im Winter. Kalt-stenotherme Arten werden hierdurch bei der Überwinterung zusehends beeinträchtigt. Innerhalb von Fließgewässern werden Arten der Äschen- und Forellenregion durch die Potamalisierung in höhere Lagen verdrängt. In Folge verringerter Sommerniederschläge sind vor allem obere Bachläufe und Feuchthabitate durch Austrocknung bedroht. Auch innerhalb trockenerer Habitate besteht ein erhöhtes Risiko, dass trockenheitsempfindliche Arten zurückgehen.
Naturschutz in den Mittelgebirgen muss die Anpassungsfähigkeit der Biodiversität an den Klimawandel stärken. Dabei sind Maßnahmen zur Erhöhung der Habitatqualität und heterogenität sowie der Konnektivität der Lebensräume auf vertikaler und horizontaler Ebene (Biotopverbund) von besonderer Bedeutung.
Effects of recent climate changes on the biodiversity of low mountain ranges Literature review on species and habitats
Montane species and habitats are highly threatened by recent climate change. Consequently, there is urgent need for action by nature conservation policy to face this challenge. This study reviews the effects of climate change on biodiversity within low mountain ranges with special reference to the Hochsauerland region (central Germany).
One of the greatest threats for biodiversity associated with climate change is the increase of winter temperatures. Especially the overwintering stages of cold-adapted species are influenced negatively by warmer winter periods. Within rivers and streams species distributed within the upper reaches shift their ranges upward due to the potamalization effect. As a cause of reduced summer precipitation wetlands and upper reaches of streams are threatened by drying up. But also in drier habitats species sensitive to dryness are endangered by reduced summer precipitation. Nature conservation within low mountain ranges must aim at promoting the adaptability of montane species and habitats to climate change. Therefore, the implementation of measures for increasing habitat quality, heterogeneity and connectivity is highly recommended.
- Veröffentlicht am

1 Einleitung
Der globale Rückgang der Biodiversität hat ein bislang unbekanntes Ausmaß erreicht. Hauptverantwortlich für das weltweite Artensterben ist der Mensch. Insbesondere die Intensivierung der Landnutzung hat erheblich zum Verlust der Artenvielfalt beigetragen (Sala et al. 2000). Seit Ende des letzten Jahrtausends steht zudem die Bedeutung des Klimawandels für den Rückgang der Artenvielfalt zunehmend im wissenschaftlichen Fokus (Leuschner & Schipka 2004). Aufgrund der dramatischen Veränderung des Klimas wird erwartet, dass der Klimawandel zukünftig stärker auf die Biodiversität einwirken wird, als dies direkte Habitatveränderungen tun (Leuschner & Schipka 2004). Aufgrund der altitudinal eingeschränkten Habitatverfügbarkeit werden vor allem für montan verbreitete Arten erhebliche Verluste prognostiziert (Engler et al. 2011).
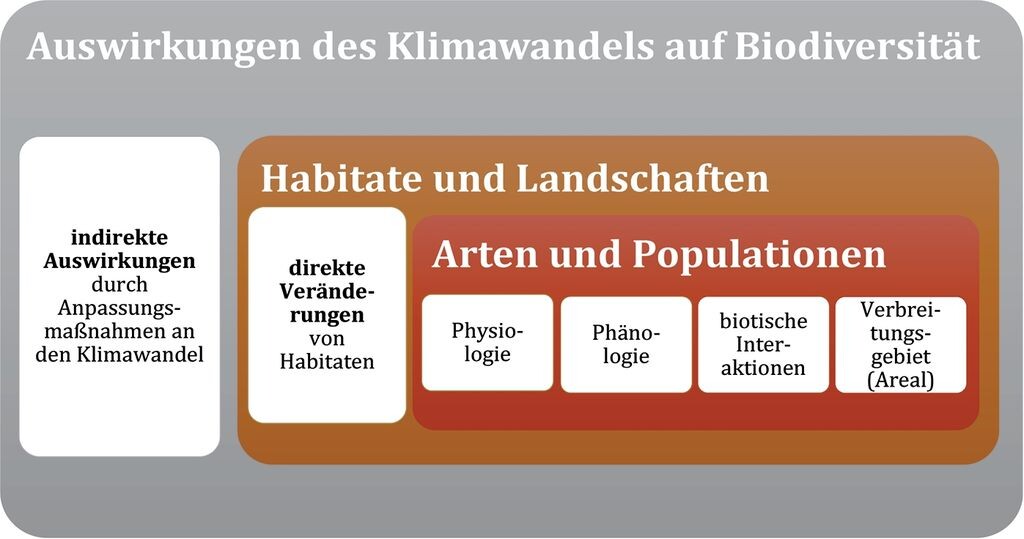
Die Änderung des Klimas wirkt auf komplexe Art und Weise auf Arten ein. Dabei spielen direkte und indirekte Auswirkungen eine Rolle (Essl & Rabitsch 2013, Mosbrugger et al. 2014) (Abb. 1). Direkte Auswirkungen auf der Art- und Populationsebene betreffen insbesondere die Physiologie, Phänologie, biotischen Interaktionen und das Verbreitungsgebiet (Areal). Auf der Habitat- und Landschaftsebene wirken sich direkte, durch den Klimawandel hervorgerufene Lebensraumveränderungen auf die Artenvielfalt aus. Indirekte Auswirkungen sind auf Anpassungsmaßnahmen an den Klimawandel insbesondere klimawandelbedingte Landnutzungsänderungen und der Ausbau regenerativer Energien zurückzuführen.
Die vorliegende Literaturstudie fasst die klimawandelbedingten Auswirkungen auf die Biodiversität in Mittelgebirgen zusammen. Ein Schwerpunkt der Betrachtungen liegt hierbei auf dem Hochsauerland (Nordrhein-Westfalen). Neben einer allgemeinen Darstellung der Auswirkungen des Klimawandels auf die Biodiversität werden insbesondere der Einfluss der Klimaänderung auf für den Untersuchungsraum repräsentative Habitattypenkomplexe (Fließgewässer, Grasland, Bergheiden, Wälder) zusammengefasst und daraus resultierende Anpassungsmaßnahmen aus Sicht des Naturschutzes abgeleitet.
2 Aktuelle Entwicklung des Klimas in Deutschland und dem Hochsauerland
In Folge der steigenden Treibhausgasemissionen seit Mitte des 20. Jahrhunderts überwiegt der menschliche Einfluss auf das Klima die natürlichen Prozesse (Jonas et al. 2005). So stieg die globale Jahresmitteltemperatur im Zeitraum von 1880 bis 2012 um 0,85°C (IPCC 2013). Für Deutschland ist im letzten Jahrhundert ein Anstieg von bis zu 1°C zu verzeichnen (Jonas et al. 2005, Rapp 2000, UBA 2006). Bis zum Ende des 21. Jahrhunderts wird ein Anstieg der Jahresmitteltemperatur von 2,5 bis 3,5°C erwartet (UBA 2006). Die Prognosen zur Niederschlagsentwicklung sind hingegen noch mit großen Unsicherheiten behaftet. Tendenziell werden vor allem im Westen Deutschlands Rückgänge der Sommerniederschläge und eine Zunahme der Winterniederschläge prognostiziert (UBA 2006).
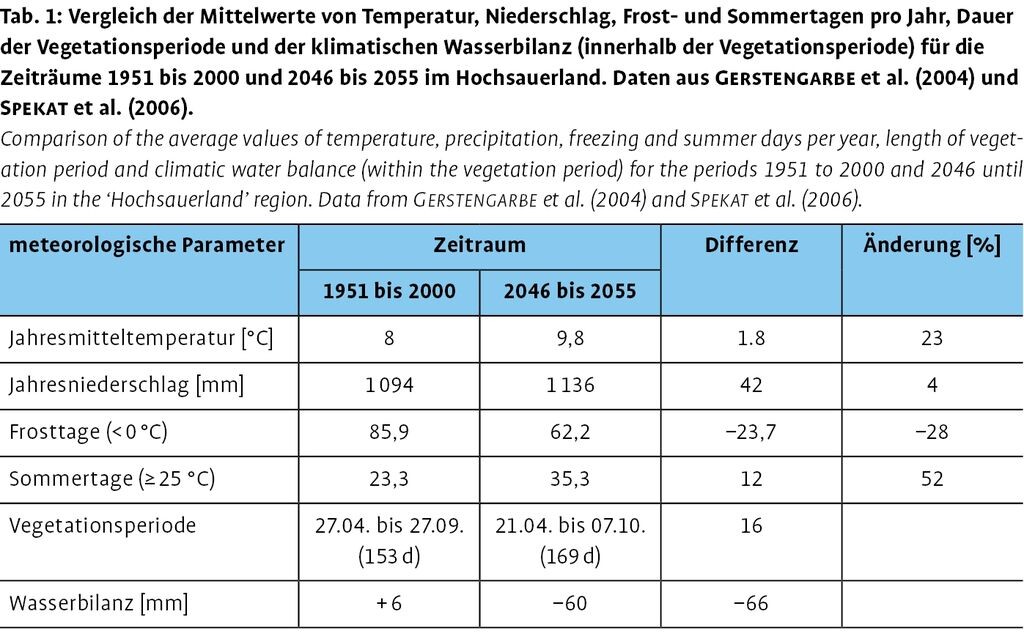
Die regionale Klimaentwicklung im Hochsauerland (Tab. 1) ist mit dem Klimatrend im Westen Deutschlands vergleichbar. Die Jahresmitteltemperatur wird von 8,0 auf 9,8°C ansteigen. In weiten Teilen des Sauerlands ist für den Zeitraum 2046 bis 2055 mit einer leichten Zunahme der Jahresniederschläge um bis zu 50mm zu rechnen. Bezogen auf das gesamte Sauerland führen die höheren Temperaturen zu einer negativen Wasserbilanz während der Vegetationsperiode von etwa 60mm.
3 Auswirkungen des Klimawandels auf die Biodiversität
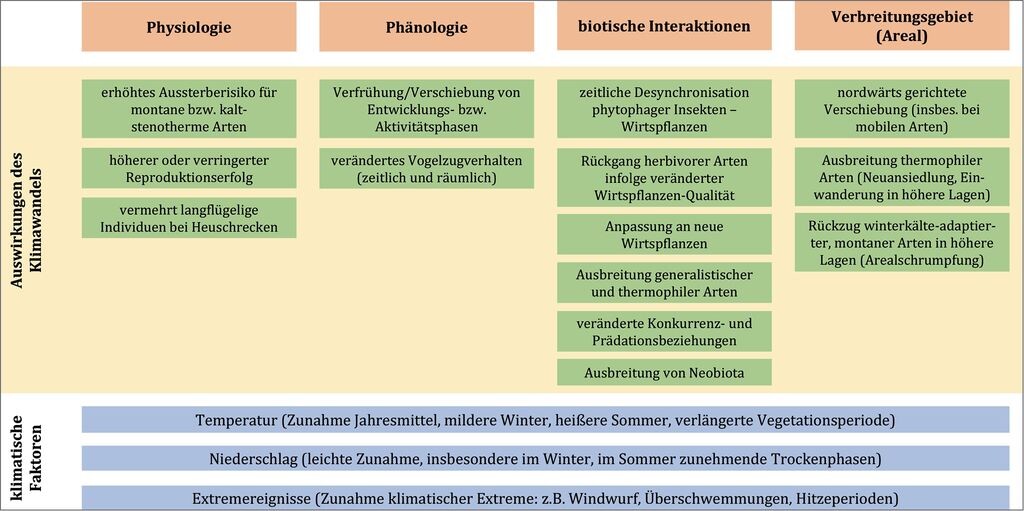
3.1 Art- und Populationsebene (Abb. 2)
3.1.1 Physiologie
Temperatur- und Niederschlagsveränderungen beeinflussen die Stoffwechselprozesse und Reproduktion von Arten. Problematisch für die Überwinterung vieler montan verbreiteter bzw. kalt-stenothermer Arten sind vor allem milde Winter mit einer verringerten Anzahl an Frosttagen (Behrens et al. 2009a, Rabitsch et al. 2010). Das wurde experimentell für den Rundaugen-Mohrenfalter (Erebia medusa) nachgewiesen (Stuhldreher et al. 2014), der im Hochsauerland vor allem in verbrachtem Magergrasland vorkommt (Fartmann 2004, Stuhldreher & Fartmann 2014). Durchschnittliche Temperaturen von 7°C während der Überwinterung führen zu einer höheren Sterblichkeit überwinternder Raupen im Vergleich zu Temperaturen um den Gefrierpunkt.
Weiter ist mit einem Rückgang von Pflanzenarten zu rechnen, die obligatorisch an eine Kälteperiode zur Überwindung der Samenruhe gebunden sind (Fernández-Pascual et al. 2013). Diese Problematik wird sich vor allem innerhalb wintermilder Gebiete bzw. tieferer Lagen der Mittelgebirge bemerkbar machen. Auch bei Amphibien und Fledermäusen haben hohe Wintertemperaturen einen erhöhten Energieverbrauch zur Folge, was eine erhöhte Sterblichkeit der Tiere nach sich ziehen kann (Reading 2007, Weise & Vohland 2010).
Neben der Erwärmung nimmt die Zunahme an klimatischen Extremereignissen einen starken Einfluss auf Arten. Hiervon sind besonders solche Taxa betroffen, die eine enge physiologische Toleranz besitzen (Rabitsch & Essl 2013a). Eine große Gefahr durch zunehmenden Trockenstress besteht beispielsweise für immobile Stadien trockenheitsempfindlicher Insektenarten (Behrens et al. 2009a). Durch lang andauernde Trockenphasen steigt das Austrocknungsrisiko der Fortpflanzungsgewässer von Libellen (Ott 2010), wodurch Arten, die im Eistadium überwintern, gegenüber als Larve überwinternden Arten gefördert werden (Schlotmann 2011, Zeuner 2014). Andererseits profitieren wärmebedürftige Arten von einer Verlängerung der Vegetationsperiode. Beispielsweise bilden diverse Libellenarten zunehmend eine zweite Generation in Mitteleuropa aus (Ott 2010).
Neben einer verlängerten Vegetationsperiode wirken sich erhöhte Temperaturen positiv auf den Reproduktionserfolg vieler Arten aus. Poniatowski & Fartmann (2011a) wiesen nach, dass durch zunehmende Frühjahrstrockenheit und -erwärmung die Larvensterblichkeit bei Roesels Beißschrecke (Metrioptera roeselii) deutlich sinkt. Durch den dadurch bedingten Dichtestress treten gehäuft langflügelige und flugfähige Individuen auf (Poniatowski & Fartmann 2011a, Poniatowski et al. 2012). Diese makropteren (langflügeligen) Tiere haben eine höhere Mobilität als kurzflügelige Individuen und sind maßgeblich für die aktuelle Ausbreitung der Art verantwortlich (Poniatowski & Fartmann 2011b, Poniatowski et al. 2012). Auch für die Reproduktion von Reptilienarten wie Zauneidechse (Lacerta agilis) und Waldeidechse (Zootoca vivipara) wurden positive Auswirkungen einer Erwärmung festgestellt (Chamaillé-Jammes et al. 2006, Olsson et al. 2010a, b).
3.1.2 Phänologie
Die Zunahme der Jahresmitteltemperatur bewirkt eine jahreszeitliche Verschiebung der Entwicklungs- bzw. Aktivitätsphasen von Pflanzen und Tieren (Menzel et al. 2006). In Deutschland hat sich in der zweiten Hälfte des 20. Jahrhunderts der Beginn der Vegetationsperiode um etwa eine Woche vorverlagert und etwa ein bis zwei Wochen verlängert (Menzel et al. 2005).
Auf Tiere wirkt sich die verlängerte Vegetationsperiode ebenfalls durch eine Vorverlagerung der Aktivitätsmuster aus (Beebee 1995, Lomann 2014, Ott 2010, Roy & Sparks 2000, Thomas 2002). Ähnliches konnte auch für Tierarten im Hochsauerland nachgewiesen werden, wie etwa beim Schlüsselblumen-Würfelfalter (Hamearis lucina; Fartmann 2006): Der Flugzeithöhepunkt lag Ende der 1990er-Jahre um zwei Wochen früher als in den 30 Jahren zuvor.
In Folge der Erwärmung wird außerdem ein verändertes Zugverhalten bei Vögeln beobachtet, indem die Tiere zunehmend länger in den Brutgebieten verweilen, früher in diese zurückkehren und/oder Überwinterungsgebiete in geringerer Entfernung zu den Brutgebieten aufsuchen (z.B. Anthes et al. 2004, Prange 2010, Sommerhage 2009, Tryjanowski & Sparks 2008, Vergara et al. 2008, Visser et al. 2009). Auch ein früherer Brutbeginn konnte bei verschiedenen Singvogelarten festgestellt werden (Dunn & Møller 2014, Porkert et al. 2014, Visser et al. 2012).
3.1.3 Biotische Interaktionen
Durch unterschiedliche Reaktionen von Interaktionspartnern auf den Klimawandel werden biotische Beziehungen beeinflusst. In Folge der Verfrühung der Vegetationsperiode erhöht sich beispielsweise das Risiko, dass eine zeitliche Desynchronisation zwischen den Entwicklungsstadien pflanzenfressender Insekten und deren Wirtspflanzen hervorgerufen wird, wenn Insekten der Entwicklung der Wirtspflanzen zeitlich nicht folgen (Singer & Parmesan 2010, van Asch & Visser 2007). Neben einer zeitlichen Entkopplung steigt durch den Klimawandel das Risiko einer räumlichen Entkopplung, wenn Interaktionspartner durch eine Veränderung des Verbreitungsgebiets bzw. Aussterbeereignisse räumlich getrennt werden. Ein hohes Gefährdungsrisiko durch eine Entkopplung ergibt sich vor allem für Arten, die spezifische Ansprüche an ihre Interaktionspartner stellen, wie etwa monophage Schmetterlingsarten (Fartmann 2006; Schweiger et al. 2008, 2012).
Auch die Nahrungsqualität wird durch den Klimawandel beeinflusst, was für Rückgänge herbivorer Arten verantwortlich sein kann. Schwarz (2014) wies nach, dass sich Trockenstress negativ auf die Vitalität von Primula veris, der Wirtspflanze des Schlüsselblumen-Würfelfalters (Hamearis lucina) im Hochsauerland, auswirkt, wodurch die Überlebensrate der Raupen verringert ist. Durch zunehmende Sommertrockenheit ist daher mit einem Rückgang der Art zu rechnen, vor allem auf warmen Südhängen (Fartmann 2006).
Andererseits gibt es Beispiele für sich ausbreitende Arten wie den Kleinen Sonnenröschen-Bläuling (Aricia agestis), die zeigen, dass eine Anpassung an neue Wirtspflanzen stattfinden kann (Thomas et al. 2001). Eine Ausbreitung dieser Art wird seit 1996 auch in Westfalen beobachtet (Fartmann et al. 2002).
Auf die chemische Zusammensetzung von Pflanzen nimmt darüber hinaus auch der erhöhte atmosphärische CO2-Gehalt Einfluss (Lindroth et al. 1997, Knepp et al. 2007). Goverde et al. (2004) wiesen experimentell nach, dass erhöhte CO2-Konzentrationen zu einem Anstieg der Larvensterblichkeit des Hauhechel-Bläulings (Polyommatus icarus) führten. Als mögliche Ursache wird ein vergrößertes C/N-Verhältnis der Wirtspflanze angesehen.
In Folge des Klimawandels werden sich die Konkurrenz- und Prädationsbeziehungen zwischen Arten ändern und neue Lebensgemeinschaften entwickeln. In montanen Regionen wird die Ausbreitung generalistischer und thermophiler Pflanzenarten in höhere Lagen gefördert, so dass sich die Konkurrenzsituation verschärft (Behrens et al. 2009b, Moradi et al. 2012). Durch die Erwärmung des Klimas wird die Ausbreitung invasiver, nicht-heimischer Arten weiter gefördert, da viele Neobiota die Fähigkeit besitzen, sich an veränderte Umweltbedingungen rasch anzupassen und neue Gebiete schnell zu besiedeln (Rabitsch & Essl 2010, Walter et al. 2009). Bei 63% der derzeit in Deutschland vorkommenden Neophyten wird angenommen, dass sie vom Klimawandel profitieren (Kleinbauer et al. 2010, Nehring et al. 2013).
3.1.4 Verbreitungsgebiet (Areal)
Es liegen zahlreiche Studien vor, die die zukünftige Verbreitung von Arten unter diversen Klimawandelszenarien modellieren (z.B. Araújo et al. 2006, Huntley et al. 2007, Levinsky et al. 2007, Settele et al. 2008, Pompe et al. 2011). Arealveränderungen sind bislang vor allem bei mobilen Artengruppen belegt.
Durch eine zunehmende Erwärmung kommt es zu einer nordwärts gerichteten Arealverschiebung. Insbesondere mobile Arten reagieren schnell auf klimatische Änderungen (Warren et al. 2001). In Großbritannien stellten Hickling et al. (2005) innerhalb von 40 Jahren eine nordwärts gerichtete Arealerweiterung bei vielen der dort vorkommenden Libellenarten fest. Vor allem südlich verbreitete Arten profitieren von der Erwärmung in Mitteleuropa und breiten sich hier aus (Ott 2010). Seit 1980 wird auch in Nordrhein-Westfalen die Ausbreitung thermophiler Libellenarten beobachtet (Conze et al. 2010).
Durch die Erwärmung ist mit einem vermehrten Rückzug winterkälteadaptierter montaner Arten in höhere Gebirgslagen zu rechnen (Lenoir et al. 2008). Verluste ergeben sich dabei vor allem an der unteren Verbreitungsgrenze (Franco et al. 2006). Daher werden bei vielen dieser Arten erhebliche Arealschrumpfungen prognostiziert. Zu den betroffenen Pflanzenarten zählen z.B. Arnika (Arnica montana) (Abb. 3) und Trollblume (Trollius europaeus) (Pompe et al. 2011). Zu den winterkälteadaptierten Insektenarten, bei denen ein Rückgang in tieferen Lagen beobachtet wird, gehört im Hochsauerland z.B. der Rundaugen-Mohrenfalter (Erebia medusa) (Glöckner & Fartmann 2003; Fartmann 2004a, b; Stuhldreher & Fartmann 2014).
Wärmeliebende Arten werden sich hingegen zunehmend in höhere Gebirgslagen ausbreiten (Franco et al. 2006, Gottfried et al. 2012, Konvicka et al. 2003, Moradi et al. 2012). Im Sauerland treten thermophile und schwach thermophile Insektenarten zusehends in höheren Lagen auf. Bei dieser Ausbreitung kann es sich sowohl um die Besiedlung neuer Areale als auch um eine Verdichtung der Vorkommen handeln. Belege für eine aktuelle Ausbreitung aufgrund der Klimaerwärmung im Hochsauerland gibt es für Heuschrecken wie Gewöhnliche Sichelschrecke (Phaneroptera falcata), Große Goldschrecke (Chrysochraon dispar), Kurzflügelige Schwertschrecke (Conocecephalus dorsalis), Sumpfschrecke (Stethophyma grossum) (Abb. 4) und Weißrandigen Grashüpfer (Chorthippus albomarginatus) (Distel et al. 2010, Poniatowski et al. 2006). Wie schon für M. roeselii erläutert (Abschnitt 3.1.1), dürften auch bei C. dispar und C. dorsalis makroptere Tiere eine entscheidende Rolle bei der Arealerweiterung gespielt haben (Distel et al. 2010, Poschmann et al. 2009); P. falcata und S. grossum sind generell gut flugfähig.
Zu den Tagfalterarten, die sich aufgrund des Klimawandels zunehmend in höheren Lagen des Hochsauerlands ausbreiten, zählen Brauner Feuerfalter (Lycaena tityrus), Kaisermantel (Argynnis paphia), Waldbrettspiel (Pararge aegeria), Kleiner Sonnenröschen-Bläuling (Aricia agestis) (Abb. 5) und Mauerfuchs (Lasiommata megera) (Brunzel et al. 2008, Fartmann et al. 2002, Fartmann 2004a, Krech 2011).
3.2 Habitat- und Landschaftsebene
3.2.1 Fließgewässer
Ein Temperaturanstieg wurde für diverse Fließgewässer in Mitteleuropa bereits nachgewiesen werden (z.B. Brunke 2008, Hari et al. 2006). Neben der Erwärmung wirken sich Niederschlagsveränderungen durch ein modifiziertes Abflussregime auf Fließgewässerlebensgemeinschaften aus. Das Ausmaß des Klimawandeleinflusses auf Fließgewässer ist von einer großen Zahl an Faktoren wie dem Einzugsgebiet und der Gewässerstruktur abhängig. Vor allem kleinere, belastete und/oder ausgebaute Gewässer sind besonders vom Klimawandel betroffen (Haase et al. 2014, Rabitsch & Essl 2013b).
Durch eine Gewässererwärmung wird für kalt-stenotherme Arten eine nordöstliche Arealverlagerung prognostiziert (Domisch et al. 2013). Auf lokaler Ebene verschieben sich die Verbreitungsgebiete von Arten stromaufwärts (Haase et al. 2014, Lehikoinen et al. 2013, Matulla et al. 2007). Vor allem die Makroinvertebratenfauna montaner Bäche ist durch den Klimawandel bedroht. Viele der hier vorkommenden Arten sind kalt-stenotherm und in ihrer Ausbreitungsfähigkeit limitiert (Haase et al. 2014, Hering et al. 2010, Tierno de Figeuroa et al. 2010). Durch erhöhte Gewässertemperaturen werden kälteadaptierte Arten zunehmend in die Oberläufe der Gewässer verdrängt, während sich wärmeliebende Arten ausbreiten (Domisch et al. 2011, Sauer et al. 2011). Am Beispiel eines mitteleuropäischen Gebirgsbachs prognostizierten Domisch et al. (2011) im Zuge des Klimawandels ein Aufwärtswandern der Arten um bis zu 80 bis 120 Höhenmeter. Aufgrund der eingeschränkten Habitatverfügbarkeit ist eine vertikale Arealverschiebung innerhalb der montanen Stufe für viele Arten allerdings unmöglich.
Ähnliches gilt auch für die im Fließgewässer-Längsverlauf typischen Fischzönosen. Durch die Gewässererwärmung wird eine Verschiebung der Zönosen stromaufwärts und eine Potamalisierung oberer Gewässerabschnitte (Ausdehnung des Potamals als Unterlauf auf Kosten des Rhithrals, der Bachregion) begünstigt (Buisson & Grenouillet 2009, Wolf & Angersbach 2010). Aufgrund der eingeschränkten räumlichen Verfügbarkeit derartiger Ausweichhabitate sind vor allem Arten der Äschen- und Forellenregionen wie die namengebende Äsche (Thymallus thymallus) und Bachforelle (Salmo trutta) gefährdet (Behrens et al. 2009a). Arten wie der Döbel (Squalius cephalus) zählen dagegen zu den Profiteuren einer Erwärmung.
In Folge der Erwärmung ist auch mit einer weiteren Ausbreitung nicht-einheimischer wärmeliebender Arten (Leuven et al. 2011, Verbrugge et al. 2012, Rabitsch et al. 2013, Wiesner et al. 2010) sowie einer Förderung der Ausbreitung von Krankheitserregern zu rechnen. Insbesondere innerhalb tiefer liegender, milderer Regionen der Mittelgebirge erhöht sich die Anfälligkeit kälteadaptierter Arten gegenüber Krankheiten (Hari et al. 2006).
Mit steigenden Temperaturen nimmt die Löslichkeit von Nährstoffen zu, so dass eine Ausbreitung von Makrophyten begünstigt und die Primärproduktion gesteigert wird (Haase et al. 2014, Hering et al. 2010). Infolgedessen ist durch verstärkte Respiration mit zunehmenden Sauerstoffdefiziten zu rechnen (Brunke 2008). Ein Rückgang von Arten, die an nährstoffarme und sauerstoffreiche Bedingungen gebunden sind, wird daher gefördert (Behrens et al. 2009b).
Durch eine erhöhte Variabilität der Niederschlagsverteilung nimmt die Häufigkeit extremer Wasserstände zu. Vor allem bei Gewässern, die aus Oberflächen- und Bodenwasserabfluss gespeist werden, steigt das Austrocknungsrisiko (Brunke 2008). Andererseits nehmen Hochwasser durch Starkregenereignisse und erhöhte Winterniederschläge zu (Haase et al. 2014). Ein daraus resultierender Oberflächenabfluss hat höhere Gehalte an gelöstem organischen Kohlenstoff innerhalb der Gewässer zur Folge (Nickus et al. 2010).
Bei kleineren Fließgewässern besteht die Gefahr, dass sie sich zunehmend in temporäre Gewässer umwandeln. Hierdurch werden univoltine Arten (Arten mit einer Generation pro Jahr) gefördert (Hering et al. 2010). Besonders problematisch wirken sich Hitzeperioden in Kombination mit Niedrigwasserabflüssen aus. Durch die Verkleinerung des Wasserkörpers wird eine Erwärmung des Wassers begünstigt, wodurch sich das Risiko von Massensterben hitzeempfindlicher Arten erhöht (Koop et al. 2007). Aufgrund einer verringerten Schleppkraft bei Niedrigwasser lagern sich verstärkt Feinsedimente und Schwebstoffe ab, wodurch die Eutrophierung begünstigt wird (Hering et al. 2010). Durch eine verstärkte Kolmation der Gewässersohle (Reduktion des Porenvolumens und Verfestigung des Sohlenmaterials durch Verschlämmung) werden Kiesbewohner zunehmend gefährdet (Verdonschot et al. 2010).
3.2.2 Grasland
Innerhalb des Graslands wirkt sich der Klimawandel abhängig vom Graslandtyp und den abiotischen Standortbedingungen unterschiedlich aus. Bei Mager- und Trockenrasen wird ein positiver Einfluss des Klimawandels erwartet. Für montane sowie feuchteabhängige Graslandhabitate wird die Gefährdung durch den Klimawandel dagegen als besonders hoch eingestuft (Behrens et al. 2009b, Bittner et al. 2011). Durch die zunehmende Erwärmung blühen viele Pflanzenarten eher (Bock et al. 2013). Aufgrund einer früher einsetzenden Vegetationsentwicklung kommt es zu einer mikroklimatischen Abkühlung bodennaher Bereiche. Hierdurch sind Tierarten, die im Frühjahr auf ein warmes Mikroklima für ihre Entwicklung angewiesen sind (z.B. als Ei oder Raupe überwinternde Schmetterlingsarten), zunehmend gefährdet (WallisdeVries & van Swaay 2006).
Langzeituntersuchungen bestätigen, dass Vegetationsveränderungen der letzten Jahrzehnte im Grasland eher auf Nutzungsänderungen zurückzuführen sind als auf den Klimawandel (Holz et al. 2013, Wesche et al. 2012). Allerdings wirken sich klimatische Extremereignisse zunehmend auch auf Graslandhabitate aus (Grime et al. 2008, Holz et al. 2013). Durch lang andauernde Trockenphasen wird die Primärproduktivität reduziert (Bütof et al. 2012, Weisshuhn et al. 2011). Für Trockenrasen ergeben sich hierdurch positive Auswirkungen, indem wärmeliebende Arten gefördert werden (Behrens et al. 2009b). Insbesondere bei thermophilen Insektenarten der Trockenrasen wie Gemeiner Sichelschrecke (Phaneroptera falcata), Kleinem Sonnenröschen-Bläuling (Aricia agestis) (Abb. 5) und Mauerfuchs (Lasiommata megera) ist von einer weiteren Ausbreitung in die Hochlagen des Sauerlands durch zunehmende Trockenperioden im Sommer auszugehen (Brunzel et al. 2008, Fartmann 2004a, Fartmann et al. 2002, Krech 2011, Poniatowski et al. 2006).
Die positiven Auswirkungen des Klimawandels werden in Magerrasen allerdings abhängig von den Standortbedingungen sein. Insbesondere am nördlichen Verbreitungsrand der Kalkmagerrasen in Mitteleuropa wie es im Sauerland der Fall ist sind viele Pflanzenarten eher an wechseltrockene als an dauerhaft trockene Bedingungen angepasst (Fartmann 2004) und entsprechend empfindlich gegenüber zunehmendem Trockenstress (Hill et al. 2008). Die negativen Auswirkungen trockener Sommer wurden bereits an diversen Enzian- und Orchideenarten beobachtet (Hill et al. 2008). Rückgänge sind daher auch bei Tierarten zu erwarten, die für ihre Entwicklung an trockenheitsempfindliche Wirtspflanzen gebunden sind, wie z.B. Kleines Fünffleck-Widderchen (Zygaena viciae), Schlüsselblumen-Würfelfalter (Hamearis lucina) und Zwerg-Bläuling (Cupido minimus) (Fartmann 2006, Hill et al. 2008, Pissens et al. 2009, Schwarz 2014). Durch Bewirtschaftung kann ein Rückgang trockenheitsempfindlicher Pflanzenarten zudem beschleunigt werden (Maalouf et al. 2012, Zwicke et al. 2013).
In Berg-Mähwiesen (Abb. 6) und Borstgrasrasen ist vor allem mit einem Rückzug montaner Arten in höhere Lagen zu rechnen. Durch zunehmend trockenere Bedingungen wird die Mineralisation beschleunigt und damit die Eutrophierung gefördert (Behrens et al. 2009b). Erhöhte Temperaturen begünstigen die Zunahme an Wärme- und Trockenheitszeigern. Negativ betroffen sind vor allem Magerkeitszeiger kühler Mittelgebirgslagen (z.B. Bärwurz Meum athamanticum), die durch die Ausbreitung wärmeliebender Tieflandsarten zurückgedrängt werden (Behrens et al. 2009b). Zu Bestandsrückgängen kommt es zudem bei Arten, die auf Winterkälte angewiesen und montan verbreitet sind, wie etwa Dukaten-Feuerfalter (Lycaena virgaureae) (Abb. 7), Lilagold-Feuerfalter (Lycaena hippothoe) und Rundaugen-Mohrenfalter (Erebia medusa) (Fartmann 2004 a, b; Fartmann & Hermann 2006, Glöckner & Fartmann 2003, Meyer 2006, Stuhldreher & Fartmann 2014).
In Feuchtwiesen bewirken die zunehmenden Fluktuationen des Grundwasserspiegels durch ausgeprägte Trockenphasen im Sommer und Herbst erhebliche Standortveränderungen. Lang andauernde Trockenphasen begünstigen eine erhöhte Nährstofffreisetzung durch Humus- und Torfmineralisation. Im Zuge des Klimawandels ist daher zunehmend mit einem Rückgang stenotoper Feuchtgebietsarten zu rechnen (Behrens et al. 2009b). Von dieser Entwicklung negativ betroffen ist z.B. der Wiesenpieper (Anthus pratensis).
3.2.3 Bergheiden
Zum Einfluss des Klimawandels auf Bergheiden gibt es bislang nur wenige Erkenntnisse; die meisten Studien behandeln Sandheiden des Tieflands. Im Fokus des wissenschaftlichen Interesses steht dabei vor allem der Einfluss des Klimawandels auf die schlüsseldominante Besenheide (Calluna vulgaris). Hierzu liegen experimentelle Studien vor, die die Reaktion von Heiden auf klimawandelbedingte Phänomene wie Trockenstress oder Starkregenereignisse untersuchen (z.B. Jentsch et al. 2008, Kreyling et al. 2008). Erste Untersuchungen belegen, dass die Besenheide äußerst empfindlich gegenüber Trockenstress ist (Jentsch et al. 2008, Peñuelas et al. 2007, Southon et al. 2012). Zunehmende Trockenschäden an der Besenheide wurden auch im Freiland bereits beobachtet (Schellenberg & Bergmeier 2014).
In Hochlagen der Mittelgebirge ist hingegen davon auszugehen, dass die Besenheide durch ausreichend hohe Niederschläge weniger durch Trockenstress beeinträchtigt wird und möglicherweise sogar von einer Erwärmung profitieren wird (Peñuelas et al. 2007). Allerdings ist die Vitalität der Besenheide von weiteren Klimafaktoren abhängig, so dass der Klimawandeleinfluss auf die Art schwer vorherzusagen ist (Kreyling et al. 2010). Insbesondere Spätfröste scheinen sich negativ auf die Vitalität bergheidetypischer Zwergsträucher auszuwirken (Borchard et al. 2014).
Durch wärmere Winter und verringerte Schneedecken erhöht sich für Zwergsträucher in montanen Heiden die Gefahr eine Schädigung durch Spätfröste. Hier ist außerdem zu erwarten, dass erhöhte Temperaturen die Mineralisation und damit die Eutrophierung begünstigen. Die Etablierung konkurrenzkräftigerer Arten in Heidebeständen durch wird dadurch möglicherweise gefördert (Behrens et al. 2009b, Kreyling et al. 2008, Werkman & Callaghan 2002). Innerhalb montaner Heiden besteht vor allem für boreal-montane sowie kalt-stenotherme Arten (Abb. 8) aufgrund von Ausbreitungs- und Habitatlimitierung ein hohes Aussterberisiko.
3.2.4 Wälder
Aufgrund der unterschiedlichen Anpassungsfähigkeit einzelner Baumarten an den Klimawandel werden langfristig erhebliche Änderungen in der Baumartenzusammensetzung heimischer Wälder erwartet. Vor allem für montan verbreitete Baumarten wie die Fichte (Picea abies) werden Arealrückgänge in Mitteleuropa prognostiziert (Kölling & Ewald 2013, Thuiller et al. 2006). Innerhalb montaner Lagen besteht ein hohes Risiko, dass die Fichte durch wärmeliebendere Arten wie die Rotbuche (Fagus sylvatica) verdrängt wird (Jantsch et al. 2014, Kölling & Ewald 2013). Die Anpassungsfähigkeit der Rotbuche an den Klimawandel ist allerdings stark von den Standortbedingungen abhängig. Insbesondere auf trockenen bzw. flachgründigen Standorten leidet die Rotbuche zunehmend unter Trockenstress (Friedrichs et al. 2009, Michelot et al. 2012, Milad et al. 2012, Scherrer et al. 2011). Es ist daher zu erwarten, dass sich trockenheitstolerante Baumarten (z.B. Quercus petraea) auf Extremstandorten zunehmend zugunsten der Rotbuche ausbreiten werden.
Vor allem für feuchteabhängige Waldtypen wie bachbegleitende Erlen- und Eschenwälder besteht ein hohes Gefährdungsrisiko durch die zunehmende Trockenheit. Innerhalb dieser Waldtypen wird die Konkurrenzkraft der dominanten Baumarten durch zunehmende Fluktuationen des Grundwasserspiegels beeinflusst. Durch verstärkte Austrocknung in Trockenphasen und Mineralisation wird die Ausbreitung mesophiler bzw. nitrophytischer Arten gefördert (Behrens et al. 2009b). Innerhalb von Hang- und Schluchtwäldern, die im Hochsauerland vor allem im Hoppecketal verbreitet vorkommen, wird sich die Erwärmung negativ auf kälteadaptierte Arten wie etwa den Berg-Ahorn (Acer pseudoplatanus) auswirken (Jantsch et al. 2014, Scherrer et al. 2011) und die Ausbreitung trockenheitsresistenterer Arten begünstigen (Behrens et al. 2009b). Hier ist außerdem mit einem vermehrten Rückgang von Arten zu rechnen, die empfindlich gegenüber erhöhten Wintertemperaturen sind (Behrens et al. 2009b). Laubwälder trockener Standorte, wie Orchideen-Buchenwälder, wird hingegen eine positive Reaktion auf den Klimawandel zugesprochen. Durch die Erwärmung und länger anhaltende Trockenperioden werden lebensraumtypische Licht- und Wärmezeiger gefördert (Lenoir et al. 2010).
Natürliche Änderungen der Baumartenzusammensetzung sind ein langsamer Prozess und werden sich erst langfristig bemerkbar machen. Hingegen lassen sich klimatisch bedingte Änderungen der Krautschicht bereits beobachten. Vor allem früh austreibende Arten profitieren von einer Vorverlagerung des Blühbeginns (Dierschke 2013). In Folge der Erwärmung werden außerdem thermophile Arten in ihrer Ausbreitung gefördert (Lenoir et al. 2010). In montanen Lagen besteht daher ein hohes Risiko, dass sich Tieflandsarten in höhere Lagen ausbreiten und montane Arten verdrängt werden (Jantsch et al. 2014). Allerdings konnten Patsias & Bruelheide (2013) am Beispiel des montan verbreiteten Wolligen Reitgrases (Calamagrostis villosa) im Gegensatz zu anderen Pflanzenarten experimentell nachweisen, dass sich erhöhte Temperaturen positiv auf die Biomasseproduktion auswirken. Durch die Förderung einzelner Arten sind daher Dominanzverschiebungen in der Krautschicht zu erwarten. Von milderen Wintern profitieren außerdem immergrüne Arten wie Efeu (Hedera helix; Dierschke 2013).
Die Zunahme klimatischer Extremereignisse nimmt besonderen Einfluss auf die Entwicklung der Wälder (Lindner et al. 2010). Vor allem lang andauernde Hitzeperioden oder Stürme werden vermehrt Baumschäden hervorrufen (Klaus et al. 2011, Usbeck et al. 2010). In Folge dessen ist mit einer Ausbreitung von Krankheiten und Parasiten zu rechnen, die langfristig zu Baumartenveränderungen führen können (Deprez-Loustau 2006, Niesar et al. 2007, Sturrock et al. 2011). Durch eine zunehmende Erwärmung und die dadurch bedingte verlängerte Flugperiode und erhöhte Reproduktionsrate werden Gradationen herbivorer Insekten zusätzlich begünstigt (Lindner et al. 2010). Vor allem für Fichtenbestände wird das Risiko gegenüber einem gehäuften Befall durch Borkenkäfer aufgrund der Empfindlichkeit der Baumart gegenüber dem Klimawandel als besonders hoch eingestuft (Müller-Kroehling et al. 2009).
3.3 Indirekte Auswirkungen durch Anpassungsmaßnahmen an den Klimawandel
Indirekte Auswirkungen des Klimawandels ergeben sich vor allem durch klimawandelbedingte Landnutzungsänderungen und den Ausbau CO2-neutraler, regenerativer Energien. Insbesondere in Regionen mit hohen Niederschlägen ist durch die verlängerte Vegetationsperiode und eine erhöhte Produktivität in Folge der Erwärmung mit einem früheren Beginn und einer Intensivierung der Graslandnutzung zu rechnen (Essl 2013). Eine früher einsetzende Bewirtschaftung wird sich möglicherweise negativ auf Arten auswirken, die phänologisch weniger gut mit dem Klimawandel Schritt halten können (Kleijn et al. 2010).
Kurzfristige Auswirkungen auf die Baumartenzusammensetzungen in Wäldern ergeben sich durch Anpassungsmaßnahmen an den Klimawandel seitens der Forstwirtschaft. Vor allem innerhalb niederschlagsreicher Regionen ist mit einer intensivierten forstlichen Nutzung zu rechnen, da die Produktivität von Waldbeständen durch die Erwärmung zunimmt (Lindner et al. 2010). Durch die Forstwirtschaft wird vor allem ein Umbau von Fichtenforsten in anpassungsfähigere Waldtypen angestrebt (Milad et al. 2012). Im Zuge des Klimawandels werden historische Waldnutzungsformen (Nieder- und Mittelwald) vermutlich neue Bedeutung erlangen, da sie auf Bestandes- und Landschaftsebene zu einer erhöhten Vielfalt beitragen und eine höhere Flexibilität des Bewirtschafters ermöglichen (Milad et al. 2012). Aufgrund des steigenden Bedarfs an Festbrennstoffen ist die Wiedereinführung historischer Waldnutzungsformen auch aus ökonomischer Sicht inzwischen wieder sinnvoll (Fartmann et al. 2013, Helbing et al. 2015).
Auch der Ausbau erneuerbarer Energien stellt den Biodiversitätsschutz vor neue Herausforderungen. Im Jahr 2013 deckten erneuerbare Energien bereits 25% des Brutto-Stromverbrauchs in Deutschland ab (Fraunhofer IWES 2014). Aufgrund des hohen Flächenbedarfs zur Erreichung der nationalen Ausbauziele erhöht sich das Risiko, dass Lebensräume durch die zunehmende Nutzung regenerativer Energie weiter zerstört werden (s. GFN & ZSW 2011).
Wie sich indirekte Folgen des Klimawandels auf Gewässer auswirken, erscheint noch ungewiss. Durch den Ausbau der Wasserkraft werden alte und stillgelegte Wasserkraftwerke verstärkt in Nutzung genommen (BfN 2010). Die ökologischen Folgen sind derzeit aber noch nicht abschätzbar.
Windenergieanlagen können stark negative Auswirkungen insbesondere auf die Populationen von Vögeln und Fledermäusen haben (Bellebaum et al. 2013, Flade 2012, Hurst et al. 2015, Pearce-Higgins et al. 2009, 2012; Rydell et al. 2010, Zahn et al. 2014). Zu direkten Verlusten kommt es durch Kollisionen mit den Rotorblättern und bei Fledermäusen zusätzlich durch das Barotrauma, bei dem aufgrund von starken Luftdruckänderungen in Nähe der Rotorblätter die inneren Organe zerreißen. Populationsökologisch relevant ist zudem die Scheuchwirkung, die von den Windrädern ausgeht. Seit 2010 erfolgt durch den Einsatz neuer Technologien ein zunehmender Ausbau von Windenergie an Mittelgebirgs- und Waldstandorten (Callies et al. 2012, Fraunhofer IWES 2014). Mit dem Rotmilan (Milvus milvus) kommt im Sauerland eine Vogelart regelmäßig vor, bei der besonders häufig Schlagopfer durch Windenergieanlagen nachgewiesen werden (Bellebaum et al. 2013).
Neben dem Ausbau der Windkraft hat der intensive Anbau von Bioenergiepflanzen erhebliche Auswirkungen auf die Biodiversität. Für den Energiepflanzenanbau wurde in den letzten Jahren ein erheblicher Umbruch von Graslandhabitaten vorgenommen (Herden et al. 2011), was unter anderem für den dramatischen Rückgang an Vögeln der Agrarlandschaft mitverantwortlich gemacht wird (Flade 2013, Hötker et al. 2014). Durch die gestiegene Nachfrage nach Festbrennstoffen ist darüber hinaus langfristig mit einer intensivierten forstlichen Nutzung der Wälder zu rechnen. Dies stellt eine Gefahr für seltene Waldarten dar, z.B. den im Sauerland verbreiteten Schwarzstorch (Ciconia nigra) (Flade 2012, Kerth et al. 2014).
4 Schlussfolgerungen
In den Mittelgebirgen besteht für viele Arten und Habitate ein hohes Gefährdungsrisiko durch den Klimawandel. Aufgrund der fehlenden subalpinen und alpinen Höhenstufe ist die Flächenverfügbarkeit begrenzt und eine Vertikalverschiebung der Areale bestenfalls geringfügig möglich (Schlumprecht et al. 2010). Ein hohes Aussterberisiko besteht insbesondere in Gipfellagen (Dirnböck et al. 2011, Pauli et al. 2003) und für boreal-montane, arktisch-alpine oder kalt-stenotherme Arten (Brunner et al. 2013, Möseler & Wunder 1999, Müller et al. 2009).
Im Gegensatz dazu existiert in tieferen Lagen der Mittelgebirge eine hohe Gefahr, dass sich wärmeliebende Tieflandarten zunehmend in höhere Lagen ausbreiten und spezialisierte oder konkurrenzschwache Arten verdrängen (Gottfried et al. 2012, Moradi et al. 2012). Zusätzlich werden politische oder land- und forstwirtschaftliche Anpassungsmaßnahmen zunehmend die Biodiversität in Mittelgebirgen beeinflussen. Insbesondere durch den Ausbau erneuerbarer Energien wird sich Flächennutzungsdruck auch in den Mittelgebirgen weiter verschärfen. Aufgrund von fehlenden Langzeituntersuchungen sind die konkreten Auswirkungen des Ausbaus erneuerbarer Energien auf montan verbreitete Arten und Habitate derzeit allerdings noch schwer abschätzbar.
Zur Vermeidung von Biodiversitätsverlusten durch den Klimawandel besteht die wichtigste Anforderung des Naturschutzes darin, die Anpassungsfähigkeit von Arten und Lebensräumen an die veränderten Umweltbedingungen zu stärken. Um dieses Ziel auf lokaler Ebene zu erreichen, ist es dringend erforderlich, die Habitatqualität- und -heterogenität innerhalb von Habitaten zu erhöhen (s. Behrens et al. 2009c, Fartmann et al. 2012, Fridley et al. 2011). Darüber hinaus kommt dem Ausbau des Biotopverbunds auf vertikaler und horizontaler Ebene eine besondere Bedeutung zu, um Ausweichbewegungen und Arealverlagerungen zu ermöglichen und bestehende Metapopulationsstrukturen auf der Landschaftsebene zu stärken (s. Behrens et al. 2009c, Fartmann et al. 2012, Jedicke 2015). Die Renaturierung von Lebensräumen ist hierbei explizit eingeschlossen und ein unverzichtbarer Beitrag. Im Zuge des Ausbaus erneuerbarer Energien ist die Berücksichtigung artspezifischer Schutzmaßahmen (z.B. ausreichend große Abstände von Brutvogelhabitaten zu Windenergieanlagen, s. LAG VSW 2014) zudem zwingend erforderlich.
Dank
Diese Publikation entstand im Rahmen des Projekts Biotopverbund als Klimaanpassungs-Strategie des Naturschutzes in der Beispielregion Naturpark Diemelsee, welches durch die Deutsche Bundesstiftung Umwelt (DBU, Az. 30530-33/2; Betreuer: Dr. Volker Wachendörfer) sowie das Hessische Ministerium für Umwelt, Klimaschutz, Landwirtschaft und Verbraucherschutz und das Ministerium für Klimaschutz, Umwelt, Landwirtschaft, Natur- und Verbraucherschutz des Landes Nordrhein-Westfalen gefördert wird. Projektträger ist der Zweckverband Naturpark Diemelsee (Ansprechpartner: Dieter Pollack). Allen genannten Institutionen und Personen sei herzlich für die Unterstützung des Projektes gedankt.
Literatur
Aus Umfangsgründen steht das Literaturverzeichnis zum Download bereit unter http://www.nul-online.de, Webcode 2231.
-
Abb. 1: Ebenen der Auswirkungen des Klimawandels auf die Biodiversität.Levels of effects of the climate change on biodiversity. -
Abb. 2: Exemplarische Auswirkungen des Klimawandels und die hierfür verantwortlichen klimatischen Faktoren auf der Ebene von Arten und Populationen.Exemplary effects of climate change and responsible climate factors on the level of species and populations. -
Abb. 3: Arnika (Arnica montana), eine in Deutschland heutzutage vorwiegend montan verbreitete Art, für die im Zuge des Klimawandels weitere Arealverluste vorhergesagt werden. © Dominik PoniatowskiLeopard’s Bane (Arnica montana), currently mainly spread in mountainous regional. For these species further habitat losses have been predicted. -
Abb. 4: Die Sumpfschrecke (Stethophyma grossum) breitet sich aktuell aufgrund der Klimaerwärmung im Hochsauerland aus. © Thomas FartmannThe Large Marsh Grasshopper (Stethophyma grossum) is currently expanding in the Hochsauerland region due climate warming. -
Abb. 5: Der Kleine Sonnenröschen-Bläuling (Aricia agestis) zählt zu den Gewinnern des Klimawandels im Hochsauerland. © Thomas FartmannThe Brown Argus (Aricia agestis) ranks among the winners of the climate change in the Hochsauerland region. -
Abb. 6: In Berg-Mähwiesen ist im Zuge des Klimawandels mit einem Rückzug montaner Arten in höhere Lagen zu rechnen. © Thomas FartmannIn mountain hay meadows it is assumed that the climate change will lead to a decrease of montane species. -
Abb. 7: Arten wie der Dukaten-Feuerfalter (Lycaena virgaureae), die auf Winterkälte angewiesen sind, zählen zu den Verlierern des Klimawandels. © Thomas FartmannSpecies such as the Scarce Copper (Lycaena virgaureae) are among the losers of climate change since they depend on cold winters. -
Abb. 8: Durch den Klimawandel dürften boreal-montane Arten der Bergheiden zunehmend verdrängt werden. © Eckhard JedickeThe climate change might cause an increasing suppression of boreal montane species of the mountain haylands. -
Auswirkungen des rezenten Klimawandels auf die Biodiversität in Mittelgebirgen -
Auswirkungen des rezenten Klimawandels auf die Biodiversität in Mittelgebirgen







Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.