Welche Effekte haben große Beutegreifer auf Huftierpopulationen und Ökosysteme?
Abstracts
Nach Jahrhunderten der Verfolgung breiten sich die Populationen verschiedener großer Beutegreifer in Nordamerika und Europa derzeit wieder aus. Dieser Beitrag stellt die theoretischen Arbeiten und die Ergebnisse von empirischen Forschungsergebnissen vor, die darauf hindeuten, dass große Beutegreifer Schlüsselarten sind, die trotz ihrer geringen Biomasse das Potenzial haben, die Bestände von Huftieren und Mesoprädatoren zu limitieren, also deren Bestände unterhalb der Kapazität ihres Lebensraumes zu halten und darüber hinaus deren Verhalten wesentlich zu beeinflussen. Dadurch können weitreichende Effekte im Ökosystem, wie die Verjüngung von Wäldern, angestoßen werden.
Allerdings weisen die bislang vorgelegten Studien vielfach methodische Schwächen auf, die durch die großen räumlichen und zeitlichen Skalen der beobachteten Prozesse begründet sind. Daher müssen deren Aussagen mit gewisser Vorsicht interpretiert werden. Darüber hinaus wird diskutiert, ob die bisher vorliegenden Forschungsergebnisse, die meist aus naturnahen Landschaften mit geringer Ökosystemproduktivität, wie dem Yellowstone Nationalpark, stammen, auf Mitteleuropa übertragen werden können.
Effects of large predators on populations of hoofed animals and on ecosystems Bottom up versus top down control
After centuries of tracking the population of large predators have started to spread again in Northern America and in Europe. The contribution introduces the theoretical investigations and the results of empiric research. These indicate that large predators despite their rather low biomass are key species which have the potential to limit the stock of hoofed mammals and meso-predators, which means that they can keep the stock of these animals below the capacity of their habitat and additionally can significantly influence their behaviour. This influence can trigger far-reaching effects in eco-systems, e.g. the rejuvenation of forests.
The studies so far presented however frequently show methodological deficits due to the large spaces and long-term scales of the processes monitored. Therefore their statements have to be interpreted carefully. Additionally the study discusses if the existing results can simply be transferred to Central Europe since they frequently stem from near-natural landscapes with a low productivity of their ecosystems, such as the Yellowstone National Park.
- Veröffentlicht am
1 Einleitung
Große Beutegreifer faszinieren seit jeher die Menschen. Gleichzeitig sind sie aber auch die Artengruppe, die am stärksten verfolgt wurde. Dadurch kam es zu einem dramatischen Rückgang ihrer Populationen mit dem Ergebnis, dass das ursprüngliche Verbreitungsgebiet z.B. der Wölfe (Canis lupus) um 33% und das der Tiger (Panthera tigris) und Löwen (Panthera leo) um 83% geschrumpft ist (Ripple et al. 2014a). Geringe Populationsdichten und Reproduktionsraten, kombiniert mit hohen Nahrungs- und Raumansprüchen, sind Charakteristika der großen Beutegreifer (Carbone et al. 1999, Cardillo et al. 2005). Daraus ergeben sich von Natur aus hohe Aussterbewahrscheinlichkeiten. Belege dafür bestehen bereits aus dem mittleren und späten Miozän, also zu einer Zeit, als es noch keine Menschen gab. Damals wiesen die großen Beutegreifer die höchsten Aussterberaten aller Säugetiere auf (Viranta 2003). Zusätzlich führt die karnivore Ernährungsweise, mit Fokus auf große Huftiere und Haustiere, zu erheblichen Konflikten mit Menschen.
In den letzten Jahrzehnten kam es in den westlichen Industrienationen zu einem Wandel der Einstellung gegenüber großen Beutegreifern. Insbesondere die urbane Bevölkerung steht ihnen positiv gegenüber und sieht sie als wichtigen Bestandteil der Ökosysteme. In Folge der gesellschaftlichen Änderungen wurden auch die gesetzlichen Normen angepasst und große Beutegreifer einem strengen Schutz unterstellt. Durch Expansion bestehender Populationen und gezielte Wiederansiedlungen haben sich die Verbreitungsgebiete einiger Arten, besonders in Nordamerika und Europa, deshalb wieder vergrößert (Chapron et al. 2014).
Allerdings trifft die Rückkehr der großen Beutegreifer nur auf geteilte Zustimmung der Bevölkerung. Insbesondere Menschen mit einem utilitaristischen Naturbild, die in ländlichen Gebieten leben, stehen dieser Rückkehr kritisch gegenüber. Deshalb stellt sich die Frage: Brauchen wir große Beutegreifer überhaupt in unserer stark vom Menschen geprägten Landschaft und sollte der Naturschutz so viel Energie und Geld in ihre Rückkehr investieren? Als Begründung für ihre Wiederansiedlung wurden bislang weniger ihre ökologische Bedeutung, sondern vor allem ethische Argumente ins Feld geführt, wie sie Wotschikowsky (1978) mit der folgenden Aussage auf den Punkt bringt. Der Luchs ist so nützlich wie Michelangelos Gemälde oder Giuseppe Verdis Aida. Die Welt dreht sich auch ohne diese Dinge, aber sie wäre ohne sie eindeutig ärmer.
In jüngerer Zeit wird der Schutz und die Wiederansiedlung von großen Beutegreifern auch als Maßnahme zum Schutz der Biodiversität (Ray 2005) und der Lösung des Wald-Wild-Konfliktes gesehen, denn sowohl theoretische als auch eine steigende Anzahl empirischer Studien zeigen, dass große Beutegreifer einen erheblichen Einfluss auf Artengemeinschaften und die biologische Vielfalt ausüben können (Estes et al. 2011, Ripple et al. 2014a).
Ziel des vorliegenden Beitrags ist es, den theoretischen Hintergrund und den aktuellen Forschungsstand zum Einfluss von großen Beutegreifern auf Huftiere und Ökosysteme zusammenzufassen, kritisch zu diskutieren und aus den Befunden Schlussfolgerungen für die Situation in Mitteleuropa zu ziehen.
2 Theoretische Grundlagen
Nach der klassischen Lehrmeinung wird die Funktion von Ökosystemen durch Energie- und Nährstoffflüsse bestimmt. Entsprechend dem Modell der ökologischen Pyramide ist jede trophische Ebene von der darunter liegenden, die als Energiequelle dient, abhängig. Damit sind die Lebewesen der höheren Ebene durch die Ressourcen der darunter liegenden in ihrer eigenen Produktion limitiert, was eine nach oben gerichtete Kontrolle der Lebensgemeinschaften zur Folge hat (Lindeman 1942). Es gibt also immer mehr grüne Pflanzen als Pflanzenfresser und mehr Pflanzenfresser als Räuber, da mit jedem Konsumptionsvorgang Energieverluste auftreten. Typischerweise ist das Verhältnis von Stufe zu Stufe 1:10. Solche nach oben gerichteten Effekte (bottom-up-effekte) treten dann auf, wenn die Wechselbeziehungen der Arten durch Nährstoffverfügbarkeit und Konkurrenz geprägt werden (Ray et al. 2005). Daraus ergibt sich die Sicht von Prädatoren als Luxusgeschöpfen, die von den verfügbaren Ressourcen in den Ökosystemen gesteuert werden und selbst keinen Einfluss auf die Energie- und Nährstoffflüsse haben.
Bereits 1960 erschienen erste theoretische Arbeiten, die diese Sichtweise in Frage stellten und den Raubtieren eine besondere Rolle in den Ökosystemen zuwiesen. In einer richtungsweisenden Veröffentlichung formulierten Hairston et al. (1960) die Hypothese, dass die Welt deshalb grün sei, weil sie nicht von Pflanzenfressern überweidet wird (Green World Hypothesis). Als ein Argument für diese Annahme führten die Autoren Kohlelagerstätten auf, die klar beweisen sollen, dass sich totes Pflanzenmaterial in geologischen Zeiträumen akkumuliert und nicht von Pflanzenfressern genutzt wurde. Wenn nun die Pflanzenfresser nicht durch dichteabhängige Prozesse, also durch die Ressourcenverfügbarkeit, reguliert werden, muss ein anderer Mechanismus wirken. Daraus schlossen sie, dass die Regulierung der Pflanzenfresserpopulationen dichteunabhängig durch Raubtiere erfolgt und nur die grünen Pflanzen an der Basis der Nahrungspyramide und die Raubtiere an deren Spitze durch dichteabhängige Prozesse reguliert werden. Mit ihrer Arbeit legten sie das Fundament für eine Top-Down-Sicht ökosystemarer Prozesse: Raubtiere haben bedeutenden Einfluss auf die Verteilung und Dichte ihrer Beutetiere.
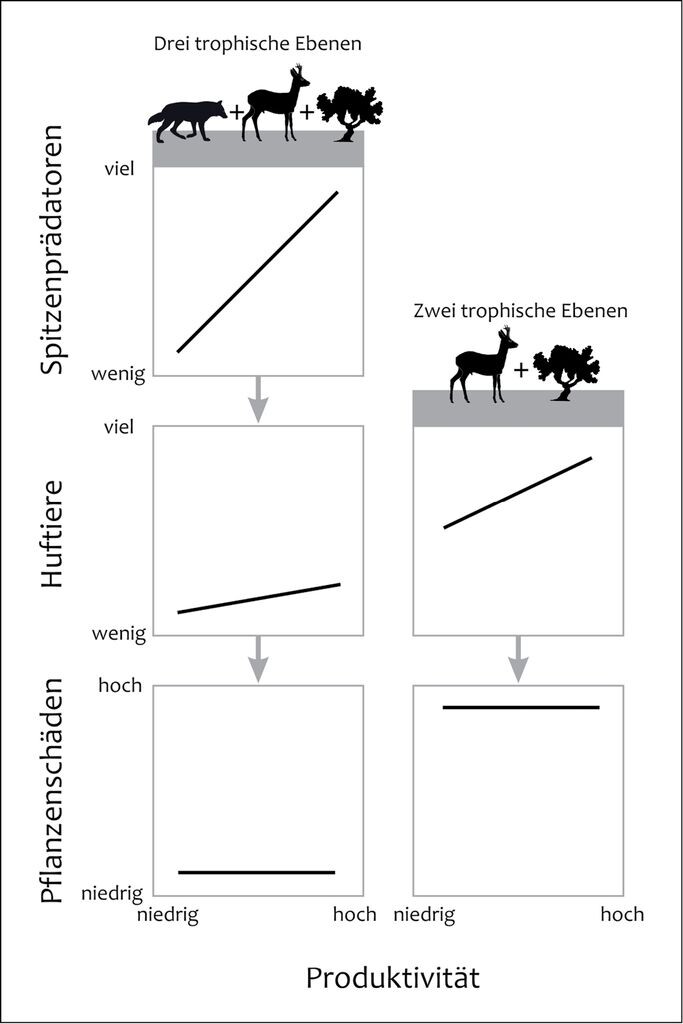
In den 1980er-Jahren verfeinerten und erweiterten Oksanen et al. (1981) die Green World Hypothesis zur Exploitation Ecosystems Hypothesis (EEH). Laut dieser Theorie hat die Primärproduktion einen Einfluss darauf, ob die Effekte in einem Ökosystem von unten nach oben oder umgekehrt verlaufen. Die EEH sagt voraus, dass in unproduktiven Ökosystemen (Primärproduktion <0,7kg/m2) mit nur zwei trophischen Ebenen die Herbivorenbiomasse linear mit der Primärproduktion ansteigt und eine von unten nach oben gerichtete Kontrolle vorliegt. Bei höherer Primärproduktion können auch Räuber leben und es entstehen Ökosysteme mit drei trophischen Ebenen. Mit steigender Primärproduktion kommt es in diesen Systemen zu einem starken Anstieg der Pflanzen- und Raubtierbiomasse, während die Pflanzenfresserbiomasse konstant bleibt. Bei einer Ausrottung der großen Beutegreifer sagt diese Theorie einen starken Anstieg der Pflanzenfresserbiomasse und hohe Schäden an Pflanzen voraus.
In den letzten Jahren häufen sich empirische Studien, die die Vorhersagen der EEH bestätigen. Allerdings kommen die meisten Arbeiten aus marinen Systemen, was vor allem damit zusammenhängt, dass Prozesse in terrestrischen Systemen schwieriger zu beobachten sind, weil sie dort in der Regel über lange Zeiträume ablaufen. Beispielsweise zeigen sich Einflüsse von Herbivoren auf Waldökosysteme aufgrund der Langlebigkeit von Bäumen erst nach Jahrzehnten.
3 Von großen Beutegreifern beeinflusste Wirkungsketten in Ökosystemen
Große Beutegreifer können über ihre Beutetiere und Mesoprädatoren (mittelgroße Raubtiere, wie z.B. Füchse Vulpes vulpes) eine Vielzahl von Effekten auf Ökosystemebene auslösen. Dabei unterscheidet man zwischen direkten Effekten, die auf Beutetiere und Mesoprädatoren gerichtet sind, und indirekten Effekten, wie der Einfluss auf die Vegetationsentwicklung oder auf die Gewässermorphologie. Die direkten Effekte werden wiederum in letale und nicht-letale Effekte unterteilt.
Das Schlüsselcharakteristikum von großen Beutegreifern ist ihr letaler Einfluss durch das Töten von Tieren. Dadurch können sie limitierend oder regulierend auf deren Populationen einwirken. Limitierung ist die Absenkung der Populationsdichte unter die Lebensraumkapazität, Regulation ist das Steuern der Population hin auf eine bestimmte Dichte, beispielsweise nach einem Rückgang oder Anstieg der Population. Dass große Beutegreifer einen stark limitierenden Einfluss auf die Beutetierpopulationen ausüben, konnten Ripple & Beschta (2012a) für die nördliche Hemisphäre zeigen. Zuvor hatten beide Autoren bereits ähnliche Einflüsse für den Westen der Vereinigten Staaten zusammengestellt (Beschta & Ripple 2009). Darüber hinaus können große Beutegreifer auch die Populationen von Mesoprädatoren beeinflussen, was bis hin zu deren lokalem Aussterben führen kann (Prugh et al. 2009, Smith et al. 2003).
Zusätzlich zum Töten üben Prädatoren bereits durch ihre bloße Anwesenheit sog. nicht-letale Effekte auf ihre Beutetiere aus, denn diese ändern ihr Verhalten so, dass die Wahrscheinlichkeit, erbeutet zu werden, möglichst gering ist. Die Lebensraumnutzung der Beutetiere orientiert sich dann nicht mehr alleine am Vorkommen von Nahrung und intraspezifischer Konkurrenz, sondern vor allem am Prädationsrisiko, das in verschiedenen Habitaten unterschiedlich stark ausgeprägt sein kann. Man spricht diesbezüglich von einer Ökologie der Angst (ecology of fear; Brown et al. 1999) und, auf die Landschaft übertragen, von einer landscape of fear, in der Gebiete mit hohem Prädationsrisiko und solche mit geringem abwechseln (Laundré et al. 2001).
Dabei kann die Reaktion der Beutetiere auf verschiedenen Skalenebenen erfolgen. So können die Tiere saisonale Wanderungen in Gebiete unternehmen, in denen die Prädatorendichte gering ist (Gower et al. 2008), oder die Lage ihrer Homeranges so wählen, dass das Prädationsrisiko minimiert wird, beispielsweise im Grenzgebiet zwischen den Territorien zweier Wolfsrudel (Mech 1977, Sinclair & Norton-Griffiths 1982, White et al. 2012). Innerhalb ihrer Homeranges halten sich Beutetiere bevorzugt in Habitaten auf, in denen Räuber sie nicht so gut erbeuten können (Kuijper et al. 2013, Laundré et al. 2001). Diese Habitate können je nach Jagdtechnik des Prädators sehr unterschiedlich sein. Während Pirschjäger, wie der Luchs, Deckung zum Anschleichen benötigen, brauchen Hetzjäger, wie der Wolf, freie Flächen, um erfolgreich zu jagen. Als weitere Strategien können Beutetiere große Gruppen bilden, um Raubtiere früher zu erkennen oder abzuwehren (Belotti et al. 2014, Creel & Winnie 2005) oder mehr Zeit mit Sichern verbringen, um ihre Umgebung abzusuchen (Creel et al. 2014, Périquet et al. 2010). Die nicht-letalen Effekte können stärker sein als die, die durch das Töten der Beutetiere hervorgerufen werden (Creel & Christianson 2008).
Durch große Beutegreifer hervorgerufene letale und nicht letale Effekte können sich über den Einfluss auf die Herbivoren und Mesoprädatoren (direkte Effekte) im Ökosystem fortsetzen (indirekte Effekte), wenn z.B. Wölfe den Kojotenbestand (Canis latrans) limitieren und dadurch die Beutetierpopulationen der Kojoten ansteigen. Erstrecken sich Effekte über mehr als eine trophische Ebene, spricht man von einer trophischen Kaskade. Die Bedeutung trophischer Kaskaden für ökosystemare Prozesse wird immer noch kontrovers diskutiert, insbesondere, weil die meisten Studien, die sich mit trophischen Interaktionen beschäftigen, Gemeinschaften von wirbellosen Tieren betreffen (Borer et al. 2005). Deshalb besteht noch immer Unsicherheit darüber, wie häufig trophische Kaskaden tatsächlich auftreten und wie stark sie sind (Ray et al. 2005).
Ein gut dokumentiertes Beispiel stammt aus einer Region in Venezuela (Lago Guri). Dort wurde ein riesiges Gebiet im Rahmen eines Staudammprojekts überflutet. Auf den Inseln, die aus der Wasseroberfläche ragten, ging die gesamte Raubtierzönose, bestehend aus Jaguar (Panthera onca), Puma (Puma concolor) und kleineren Prädatoren, verloren. Das hatte weitreichende Folgen auf das gesamte Ökosystem, namentlich auf die Bestäuber, Samenverbreiter, Samenprädatoren, Mesoprädatoren, Blattfresser, das Vogelvorkommen, das C/N-Verhältnis der Böden und auf die Gehölzverjüngung (Terborgh et al. 2001). Bislang konnten für sieben von 31 auf der Erde vorkommenden großen Beutegreifern (Körpergewicht >15kg) trophische Kaskaden nachgewiesen werden: für Löwe, Leopard (Panthera pardus) Dingo (Canis lupus dingo), Seeotter (Enhydra lutris), Eurasischer Luchs (Lynx lynx), Wolf und Puma (Ripple et al. 2014a).
5 Was wissen wir über den Einfluss von großen Beutegreifern auf Huftiere in Europa?
In Europa gibt es nur zwei große Beutegreifer, die sich vor allem von Huftieren ernähren, der Wolf (Canis lupus) und der Eurasische Luchs (Lynx lynx). Braunbär (Ursus arctos) und Rotfuchs (Vulpes vulpes) spielen nur als Prädatoren von Jungtieren eine Rolle, Vielfraß (Gulo gulo) und Goldschakal (Canis aureus) haben nur eine beschränkte Verbreitung. Deshalb werden diese Arten hier nicht weiter betrachtet. Bei den Huftieren liegt der Schwerpunkt auf den in Mitteleuropa am weitesten verbreiteten Arten Reh (Capreolus capreolus), Rothirsch (Cervus elaphus) und Wildschwein (Sus scrofa).
5.1 Eurasischer Luchs
Der Eurasische Luchs hat ein riesiges Verbreitungsgebiet, das sich von den Pyrenäen über die gesamte Taiga bis nach Kamtschatka erstreckt. In weiten Teilen Westeuropas wurde er in den letzten Jahrhunderten ausgerottet (Breitenmoser 1998).
Aufgrund ihrer einzelgängerischen Lebensweise und ihres geringen Gewichts jagen Luchse vor allem kleine Huftiere. Bevorzugt wird das Reh, bei dem Luchse alle Altersklassen und Geschlechter erbeuten können. Dadurch ergibt sich ein starker Einfluss auf Rehpopulationen, der nicht kompensatorisch ist. Dort, wo sie vorkommen, werden auch Rentiere und Gämsen häufig erbeutet. Insbesondere in Waldgebieten fallen Gämsen (Rupicapra rupicapra) dem Luchs leicht zum Opfer, da ihre Feindvermeidungsstrategie auf Lebensräume oberhalb der Waldgrenze angepasst ist. Bei Rothirschen erbeuten Luchse vor allem Kälber und einjährige Weibchen, während sie Wildschweine meiden (Belotti et al. 2014, Hayward et al. 2012, Mattisson et al. 2014, Podolski et al. 2013).
Im Durchschnitt reißen Luchse etwa 50 Huftiere pro Jahr (Belotti et al. 2015, Molinari-Jobin et al. 2002, Okarma et al. 1997). Dabei unterscheiden sich die Werte stark zwischen Geschlechtern und Altersklassen. Während Weibchen mit Jungen etwa 80 Huftiere pro Jahr zur Strecke bringen, liegt der Wert für subadulte Luchse bei etwa 45 Beutetieren. Die Prädation je Flächeneinheit lag in Polen bei 1,1 bis 1,8 Rehen und 0,4 bis 0,7 Rothirschen je km² (Okarma et al. 1997) und im Böhmerwald bei 1,2 Rehen und weniger als 0,1 Rothirschen je km² und Jahr (Belotti et al. 2015). In der Schweiz betrug sie 0,5 Rehe und 0,9 Gämsen je km² (Molinari-Jobin et al. 2002). Der Anteil der Luchsprädation an der jährlichen Mortalität der Rehe und Gämsen lag dabei zwischen 9 und 56% (Breitenmoser & Breitenmoser-Würsten 2008, Jedrzejewski et al. 2010).
Dass Luchse lokal einen sehr starken Einfluss auf ihre Beutetiere haben können, zeigt die Entwicklung der Beutetierpopulationen in einem schneereichen Hochtal im Schweizer Wallis. Dort hatte der Luchs direkt nach seiner Wiederansiedlung den Gämsenbestand von 800 auf 300 bis 400 Tiere reduziert und den geringen Rehbestand von 70 Tieren vorübergehend ganz zum Verschwinden gebracht (Breitenmoser & Haller 1993).
Auch großflächig können Luchse zu einem Rückgang ihrer Beutetiere beitragen, was ebenfalls in den Schweizer Alpen dokumentiert wurde. Zwischen 1990 und 1999 stieg dort der Luchsbestand, was zu einem Rückgang des Rehabschusses von 1000 auf unter 300 Stück führte. In dieser Zeit erbeuteten die Luchse fast 40% des geschätzten Frühlingsbestands der Rehe (Breitenmoser et al. 2010). Allerdings gibt es auch Untersuchungen aus den Schweizer Alpen und dem Schweizer Jura, bei welchen der Anteil der Luchsprädation am Rehbestand geringer als 10% war und damit der Einfluss auf seine Beutetiere als gering eingestuft wurde (Breitenmoser & Haller 1987, Molinari-Jobin et al. 2002).
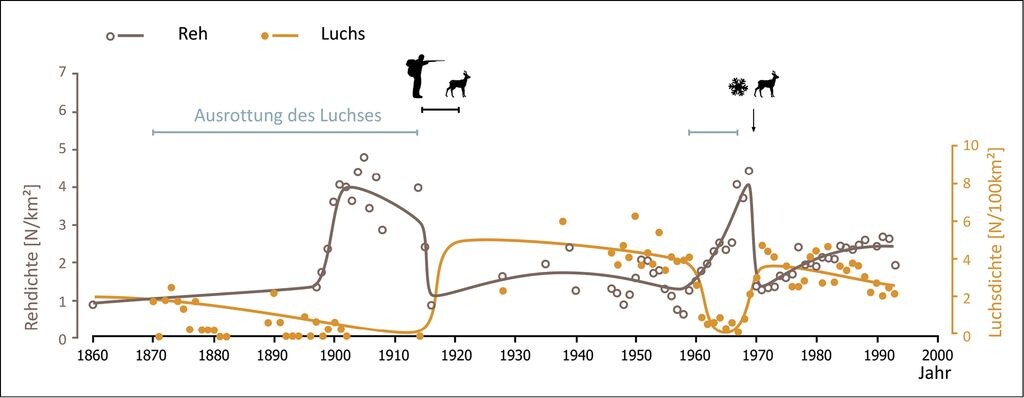
Aufschlüsse erlauben auch die über 140 Jahre langen Zeitreihen zur Bestandsentwicklung von Luchs und Reh aus dem Wald von Bialowieza. Diese Datenreihen zeigen deutlich, dass Luchse in der Lage sind, ihre Beutetierbestände zu limitieren: In den Jahren, in denen Luchse ausgerottet waren, lag die Rehdichte mit 4 Tieren/km² doppelt so hoch wie in den Zeiträumen, in denen Luchse vorkamen. Diese Ergebnisse konnten auch in einer weiteren Studie im gleichen Untersuchungsgebiet zwischen 1984 und 1993 bestätigt werden. Damals wurden 74% aller tot aufgefundenen Rehe vom Luchs erbeutet (Jdrzejewska & Jdrzejewski 1998, Jedrzejewski et al. 1993). Auch in Schweden wurde nach der Rückkehr der Luchse ein starker Rückgang der Rehpopulation nachgewiesen. Parallel dazu gingen die Rehabschüsse von 0,5 auf 0,2 Tiere je km² um mehr als die Hälfte zurück (Andrã & Liberg 2015). Ins Bild passen auch die Ergebnisse aus dem Bayerischen Wald, wo etwa 50% der tot aufgefundenen Rehe durch den Luchs erbeutet wurden und die Überlebensrate der Rehe nach Rückkehr der Luchse signifikant niedriger lag (Heurich et al. 2012). Darüber hinaus konnte in Norwegen und Frankreich ein geringeres Wachstum der Rehpopulationen in Gebieten mit Luchsvorkommen nachgewiesen werden (Nilsen et al. 2009).
Luchse sind darüber hinaus auch in der Lage, Mesoprädatoren zu limitieren (Melis et al. 2009, Pasanen-Mortensen et al. 2013). Als Konsequenz können Veränderungen in der Luchspopulation weitreichende Auswirkungen auf das Ökosystem haben. So wurde für Finnland nachgewiesen, dass der Anstieg der Luchspopulation einen Rückgang der Rotfuchsbestände zur Folge hatte, was wiederum eine Erholung der Bestände von Birk- und Auerhuhn (Tetrao tetrix und T. urogallus) sowie des Schneehasen (Lepus timidus) bewirkte (Elmhagen et al. 2010). Pasanen-Mortensen et al. (2013) konnten in einer europaweiten Studie ebenfalls nachweisen, dass Luchse die Bestände von Rotfüchsen limitieren.
Bislang gibt es nur wenige Arbeiten, die sich mit der Frage beschäftigen, ob sich die durch den Eurasischen Luchs ausgeübte Prädation auch auf die Habitatnutzung seiner Beutetiere auswirkt. Samelius et al. (2013) haben dazu die Situation vor und nach der Rückkehr der Luchse in Mittelschweden verglichen und konnten keine Hinweise darauf finden, dass Rehe Habitate mit einem hohen Prädationsrisiko weniger häufig aufsuchen. In einer Studie aus Norwegen kommen Lone et al. (2014) bei der Betrachtung von Habitatstrukturen zu dem Ergebnis, dass in dichteren Waldbeständen das Risiko für Rehe, gerissen zu werden, höher ist. Folgerichtig konnten Ewald et al. (2014) im Bayerischen Wald zeigen, dass Rehe in Gebieten mit Luchsvorkommen vor allem an Orten wiederkäuen, die ihnen eine gute Sicht bieten, so dass sie einen sich anpirschenden Luchs besser entdecken können.
Indirekte Einflüsse auf die Vegetation sind bei dem starken Prädationsimpakt des Luchses zwar wahrscheinlich, wurden aber bislang noch nicht schlüssig nachgewiesen. Nur aus dem Schweizer Oberland gibt es Hinweise, dass der kombinierte Einfluss von Luchs und Jagd zur einer Verjüngung des Waldes beigetragen hat (Rüegg et al. 1999). Für Pumas, die eine ähnliche ökologische Nische besetzen, wurden ebenfalls indirekte Effekte auf die Vegetationsentwicklung festgestellt. Im Yosemite und im Zion Nationalpark limitieren Pumas Maultierhirsche (Odocoileus hemionus), so dass der Verbissdruck auf Gehölzpflanzen sank (Ripple & Beschta 2006, 2008). Darüber hinaus kann das von Pumas verursachte Prädationsrisiko dazu führen, dass sich Hirsche in die Nähe von menschlicher Infrastruktur zurückziehen, um nicht erbeutet zu werden. Das führt zu hohen Hirschdichten in menschlicher Umgebung und einer höheren Verbissbelastung (Ripple & Beschta 2006, 2008). Auch die Landschaftsstruktur beeinflusst die Stärke der Effekte des Pumas auf seine Beutetiere. So reißen Pumas in 73% der Fälle ihre Beutetiere in der Nähe von Waldrändern und nur 6% auf offenen Flächen. In einer Landschaft mit großen Freiflächen haben also Maultierhirsche einen Vorteil, während Pumas in Landschaften mit vielen Grenzlinien leichter Beute machen können (Laundré & Hernández 2003).
5.2 Wolf
Wölfe sind aufgrund ihrer weiten Verbreitung, dem Jagen in Rudeln und ihrer ganzjährigen Aktivität die bedeutendsten Jäger von Huftieren in der nördlichen Hemisphäre (Peterson et al. 2003). Sie sind in der Lage, alle in ihrem Verbreitungsgebiet wild vorkommenden Huftierarten zu erbeuten, vom Reh bis zum Wisent, und je nach Verfügbarkeit von Beutetieren kann sich ihr Nahrungsspektrum ändern. Allerdings bevorzugen sie Rothirsche, auch wenn andere Arten im gleichen Gebiet leben (Gazzola et al. 2005, Jedrzejewski et al. 2010, Jdrzejewski et al. 2000). Bei dieser Art erbeuten Wölfe vor allem Kälber und alte oder verletzte Tiere (Wrigth et al. 2006).
Beim Wildschwein ergibt sich ein nach Mittel- und Südeuropa differenziertes Bild. Während Wildschweine in Mitteleuropa eher als Beute gemieden werden, werden sie in Südeuropa bevorzugt (Jdrzejewski et al. 2000, Mattioli et al. 2004). Dies hängt vermutlich damit zusammen, dass den Wölfen in Mitteleuropa mehr Alternativbeute zur Verfügung steht, insbesondere die bevorzugten Rothirsche. Rehe sind für Wölfe indifferent, sie werden weder bevorzugt gerissen noch gemieden, nur in der Lausitz stellen sie ein bevorzugtes Beutetier dar (Ansorge et al. 2006, Jdrzejewski et al. 2010).
Die Prädationsrate von Wölfen wurde bislang in Skandinavien, Polen und Italien bestimmt. Dabei fraß ein Wolf in Skandinavien im Durchschnitt 29 Elche und 1,5 Rehe pro Jahr. In Polen waren es 27 Rothirsche, 12 Wildschweine und 2 Rehe (Jdrzejewski et al. 2002, Sand et al. 2005). Auf die Fläche bezogen entspricht das 0,6 bis 1,1 Rothirsche, 0,04 bis 0,3 Rehe und 0,2 bis 0,4 Wildschweine je km² und Jahr (Jdrzejewski et al. 2002). In Italien wurden 0,2 bis 0,7 Rothirsche, 0,2 bis 1,2 Rehe und 0,07 bis 0,3 Gämsen je km² und Jahr erbeutet (Gazzola et al. 2005). Der Anteil der Wolfsprädation an der jährlichen Mortalität der Beutetiere lag dabei zwischen 7 und 40% (Jdrzejewski et al. 2010).
Für Mitteleuropa gibt es nur wenige Studien, die sich mit dem Einfluss von Prädatoren auf ihre Beutetiere beschäftigen. Die Ergebnisse, die aus dem Wald von Bialowieza vorliegen, zeigen, dass sich Prädation durch Wölfe und menschliche Jagd addieren und zu einem Rückgang der Rothirschpopulation führen können (Jdrzejewski et al. 2000). Die weitaus meisten Forschungsergebnisse zum Einfluss von Wölfen auf die Huftierbestände stammen aus Nordamerika und hier vor allem aus dem Yellowstone Nationalpark, wo zwischen 1995 und 1996 Wölfe wiederangesiedelt wurden. Anschließend verringerte sich der Wapitibestand innerhalb eines Jahrzehnts um die Hälfte. Dies war aber nicht nur eine Folge der Wolfsprädation, sondern auch der Bejagung der Wapitis außerhalb der Parkgrenzen und dem Rückgang der Niederschläge in der Vegetationsperiode, der zu einer schlechteren Ernährung der Tiere beigetragen hat (Hamlin et al. 2008, Hebblewhite et al. 2005, Vucetich et al. 2005, White & Garrott 2005). Auch die Habitatnutzung der Wapitis änderte sich durch die Rückkehr der Wölfe. Während sich die Wapitis in den Zeiten vor der Wiederansiedlung vor allem auf offenen Flächen aufhielten, wechselten sie danach in Wälder, die ihnen bessere Fluchtmöglichkeiten geben (Hernández & Laundré 2005). Darüber hinaus halten sich die Wapiti nach Rückkehr der Wölfe im Winter vor allem in den schneeärmeren Tallagen außerhalb des Nationalparks auf, da sie dort schlechter gejagt werden können (White et al. 2012).
Durch den Rückgang der Beutetiere wurden viele indirekte Effekte im Yellowstone Nationalpark ausgelöst. Am auffälligsten ist der Rückgang des Aspenverbisses von 100% im Jahr 1998 auf 20% in 2010 mit dem Ergebnis, dass die Bäume mittlerweile eine Größe erreicht haben, in der sie nicht mehr verbissen werden können und sich kleine Wäldchen ausbilden (Ripple & Beschta 2012b). Ausgelöst durch das verstärkte Wachstum der Weiden und Aspen, stieg die Anzahl der aktiven Biberkolonien zwischen 1996 und 2009 um mehr als 50% (Smith & Tyers 2012) und es konnte auch eine Reaktion der Singvögel beobachtet werden (Baril et al. 2011). Durch den nachlassenden Äsungsdruck der Wapitis konnten sich auch die beerentragenden Zwergsträucher besser entwickeln, was wiederum dazu geführt hat, dass sich ihr Anteil an der Nahrung der Grizzlybären vervielfacht hat (Beschta & Ripple 2012, Ripple et al. 2014b). Allerdings bleibt der kausale Zusammenhang zwischen Wiederansiedlung der Wölfe und der Erholung der Vegetation nicht unwidersprochen, da die Erholung der Vegetation schon vor der Wiederansiedlung der Wölfe eingesetzt haben soll (Kauffman et al. 2010). Unzweifelhaft haben Wolfsrisse einen wichtigen Einfluss auf die Verfügbarkeit von Aas als Nahrungsgrundlage für Scavenger. Im Yellowstone Nationalpark konnte gezeigt werden, dass Wölfe zu einer dauerhaften Verfügbarkeit von Aas beitragen und diese nicht mehr auf den Ressourcenimpuls im Spätwinter beschränkt ist. An den Rissen konnten 30 verschiedene Säugetier- und Vogelarten sowie mehr als 57 Käferarten nachgewiesen werden. Sie haben also auch einen positiven Einfluss auf die Biodiversität des Ökosystems (Wilmers et al. 2003).
Parallel zum Rückgang der Wapitis wurde ein Rückgang der Kojotenpopulation um 39% beobachtet (Berger & Gese 2007), der ebenfalls weitreichende indirekte Effekte zur Folge hatte. In Gebieten mit Wolfsvorkommen war die Überlebenswahrscheinlichkeit von Kitzen der Pronghornantilope (Antilocapra americana) viermal höher als in Gebieten ohne Wölfe (Berger et al. 2008). Da Kojoten die Umgebung von Wurfplätzen der Wölfe meiden, kam es in einer Entfernung von bis zu 3km auch zu einem Anstieg von Kurzschwanzmäusen, der in einer größeren Entfernung nicht zu beobachten war (Miller et al. 2012).
Dass die Beobachtungen aus dem Yellowstone Nationalpark kein Einzelfall sind, zeigen weitere Studien aus dem Westen der Vereinigten Staaten (Beschta & Ripple 2009). Durch ein großflächiges Reduktionsprogramm für Wölfe ergab sich im Yukon ein quasi experimenteller Ansatz, um den Einfluss der Wölfe auf ihre Beutetiere zu untersuchen. Bereits nach fünf Jahren führte die Reduktion der Wolfsbestände um 80% zu einem Anstieg der Elch- und Rentierpopulationen (Hayes et al. 2003). Im Isle Royal Nationalpark im Oberen See werden seit mehr als 50 Jahren die Entwicklungen der Wolf- und der Elchpopulation verfolgt. Die Untersuchungen zeigen, dass Wölfe einen von oben nach unten gerichteten Einfluss auf das Ökosystem ausüben können und es der Tanne erlauben, sich in den Phasen mit hoher Wolf- und niedriger Elchpopulation zu verjüngen (McLaren & Peterson 1994). Auch in den stärker von Menschen beeinflussten Gebieten in Wisconsin im Osten der Vereinigten Staaten üben Wölfe einen starken Einfluss auf Weißwedelhirsche (Odocoileus virginianus) aus, der in Gebieten mit einer hohen Wolfsdichte zu einer höheren Artenzahl an Kräutern und Sträuchern führt. Ein Einfluss auf die Gehölzvegetation konnte jedoch nicht festgestellt werden (Callan et al. 2013).
Aus Mitteleuropa gibt es nur zwei Studien, die sich mit nicht letalen und indirekten Effekten durch Wölfe beschäftigen. Beide stammen aus dem Wald von Bialowieza und kommen zu widersprüchlichen Ergebnissen. Theuerkauf & Rouys (2008) fanden heraus, dass Huftiere die am stärksten durch Wölfe genutzten Bereiche nicht mieden und im Gegenteil forstliche Eingriffe und Jagd die Huftierdichte stärker beeinflussten als das Prädationsrisiko durch Wölfe. Kuijper et al. (2013) verglichen den Verbissdruck in Kerngebieten von Wolfsrudeln mit deren Umgebung und fanden dort mit 50,5% einen signifikant geringeren Verbissdruck als außerhalb, wo 58,3% der Verjüngung verbissen wurde. Dies führten sie auf die fünfmal höhere Prädation innerhalb der Kerngebiete zurück (Jdrzejewski et al. 2002).
5.3 Kombinierter Prädationsdruck
Insbesondere der kombinierte Prädationsdruck mehrerer Raubtierarten kann Huftiere limitieren. So zeigen die Studien aus Bialowieza, dass Rehe vor allem von Luchsen erbeutet werden. 52 bis 85% des jährlichen Zuwachses wird von Luchsen und nur 8 bis 10% von Wölfen genutzt (Jdrzejewska & Jdrzejewski 1998). Auch in der Studie von Melis et al. (2009) war die Rehdichte in Gebieten mit Vorkommen von Luchs und Wolf mit etwa 1,8 Tiere/km² viel geringer als in Gebieten, in denen nur Wölfe lebten (10,5/km²) oder weder Luchs noch Wolf jagten (15/km²). Beim Rothirsch wurden im Wald von Bialowieza 32 bis 47% des jährlichen Zuwachses von Wölfen und 21 bis 43% von Luchsen erbeutet. Bei Wildschweinen waren es 10 bis 17%. Bei Elch und Bison ist der Prädationsdruck sehr gering und für Luchse sind diese Tierarten zu groß und wehrhaft (Jdrzejewska & Jdrzejewski 1998).
In Nordamerika und Eurasien waren die Huftierdichten in Gebieten ohne Wolfsvorkommen im Durchschnitt sechsmal höher als in Gebieten mit Wölfen (Ripple & Beschta 2012a). Entsprechend den Annahmen der EES konnten Ripple & Beschta (2012a) zeigen, dass die Huftierdichten mit zunehmender Primärproduktion nur geringfügig anstiegen, wenn Wölfe und Bären gemeinsam vorkommen. Übereinstimmend kommt auch Crête (1999) zu dem Ergebnis, dass die Huftierbiomasse in einem Produktivitätsgradienten in Nordamerika nicht ansteigt, wenn Wölfe vorkommen, sich aber in Gebieten ohne Wölfe um den Faktor fünf erhöht. Aus Nordamerika ist auch bekannt, dass der kombinierte Prädationsdruck von Wolf und Bär Huftierpopulationen limitieren kann, während Bären alleine dazu nicht in der Lage sind (Gasaway et al. 1992b, Mech & Peterson 2003, Messier 1994, Orians et al. 1997). Bären fressen immer wieder an von Wölfen und Luchsen erbeuteten Tieren. Das hat zur Folge, dass die beraubten Prädatoren mehr Beutetiere reißen müssen, als sie selbst benötigen (Krofel et al. 2012). Darüber hinaus können Bären auch einen hohen Anteil neugeborener Huftiere fressen (Zager & Beecham 2006). Vor allem bei niedrigen Huftierdichten ist der Einfluss der Bären additiv, da sie die Jungtiere fressen, bevor sie sich vermehren können. Wenn die Bären noch durch zusätzliche Nahrung in Form von Raubtierrissen, insbesondere in Zeiten mit Nahrungsengpässen, profitieren, kann das eine höhere Bärendichte zur Folge haben, was wiederum die Prädation an neugeborenen Huftieren erhöht. Damit wirken Bären quasi wie ein Verstärker, der den Effekt von Wölfen und Luchsen steigert.
5.4 Interaktion mit dem Klima
Neben dem Vorkommen von Prädatoren sind die Produktivität des Ökosystems, die Winterstrenge und die Waldbedeckung die wichtigsten Faktoren, die die Rehdichte in Europa bestimmen (Melis et al. 2009). Dabei beeinflussen die Produktivität des Lebensraums und die Winterstrenge die limitierenden Effekte der Räuber. In den produktiven Gebieten liegen die Rehdichten bei Vorkommen von Beutegreifern 20 bis 40% niedriger als in ähnlichen Gebieten ohne Beutegreifervorkommen. In weniger produktiven Lebensräumen ist der Einfluss wesentlich stärker. Der gleiche Effekt konnte in Bezug auf die Winterstrenge beobachtet werden. Je strenger die Winter, desto stärker der Einfluss von großen Beutegreifern (Melis et al. 2009). Diese Ergebnisse werden durch die Analyse der Datenreihe, die für den Wald von Bialowieza vorliegt, bestätigt. Dort konnte gezeigt werden, dass während kühler, weniger produktiver Perioden die Beutetierbiomasse durch die Luchse 40 bis 50% unter der Lebensraumkapazität gesenkt wurde, während es in wärmeren produktiveren Perioden nur 10 bis 20% waren (Jdrzejewska & Jdrzejewski 2005). Es zeigt sich also, dass der Einfluss der Prädatoren in ungünstigeren Lebensräumen stärker ist als in günstigen. Anders stellt sich die Situation bei den Wildschweinen dar. Hier kamen Melis et al. (2006) zu dem Ergebnis, dass Winterstrenge und Lebensraumproduktivität die Hauptfaktoren sind, die die Variation der Populationsdichten in Europa bestimmen. Das Vorhandensein von Räubern hatte nur einen geringen, nicht signifikanten Einfluss (Melis et al. 2006). Über die Faktoren, die die Rothirschpopulationen in Europa beeinflussen, liegen noch keine Forschungsergebnisse vor.
6 Schlussfolgerungen
In den letzten Jahren wurden zunehmend Forschungsergebnisse publiziert, die darauf hindeuten, dass große Beutegreifer Schlüsselarten sind, die trotz ihrer geringen Biomasse das Potenzial haben, die Bestände von Huftieren und Mesoprädatoren zu limitieren, also deren Bestände unterhalb der Kapazität ihres Lebensraums zu halten und darüber hinaus deren Verhalten wesentlich zu beeinflussen. Die Ergebnisse deuten auch darauf hin, dass dadurch weitreichende Effekte im Ökosystem, wie die Verjüngung von Wäldern, angestoßen werden können. Damit stellt das Vorkommen großer Beutegreifer nach Meinung vieler Wissenschaftler einen wichtigen Beitrag für den Erhalt naturnaher Ökosysteme dar (Estes et al. 2011, Ripple et al. 2014a, Terborgh & Estes 2010). Der Ausschluss von großen Beutegreifern aus der Landschaft kann deshalb zu verarmten Lebensgemeinschaften und einer veränderten Biodiversität führen und wird neben der Klimaveränderung als einer der bedeutendsten Eingriffe des Menschen in die Natur angesehen (Ripple et al. 2014a).
Dabei ist es wichtig festzustellen, dass die Effekte, die große Beutegreifer auf ihre Beutetiere und Mesocarnivoren ausüben, nicht von Menschen nachgeahmt werden können. Selbst wenn es gelingt, die Huftierbestände durch Managementmaßnahmen zu kontrollieren, kann es zu unerwünschten Folgen kommen, die den Schutzzielen entgegenlaufen (Möst et al. 2015). Die natürliche Rückkehr und die aktive Wiederansiedlung von großen Beutegreifern sind ein wichtiger Ansatz zur Renaturierung von Ökosystemen, der z.B. dazu beitragen kann, die Resilienz von Waldökosystemen gegenüber dem Klimawandel zu erhöhen. Insbesondere in Großschutzgebieten sollte es das Ziel sein, die natürliche Artenausstattung an Prädatoren wieder herzustellen, um einen sinnvollen Prozessschutz zu gewährleisten (Günther & Heurich 2013).
Allerdings weisen die bislang vorgelegten Studien oft methodische Schwächen auf, die durch die großen räumlichen und zeitlichen Skalen der beobachteten Prozesse begründet sind, so dass deren Aussagen vorsichtig interpretiert werden müssen. Auch stellt sich die Frage, ob die Ergebnisse ohne weiteres auf Mitteleuropa übertragen werden können. Dies liegt erstens daran, dass die Fallstudien aus naturnahen Ökosystemen, meist großen Nationalparken mit geringem menschlichem Einfluss stammen, Gebiete, die es in Mitteleuropa nicht mehr gibt. Zweitens liegen die meisten Untersuchungsgebiete in unproduktiven und harschen Lebensräumen, wie dem borealen Nadelwald oder Gebirgsökosystemen, da es in den letzten Jahrzehnten nur dort freilebende Wölfe und Luchse gab.
Im Vergleich dazu sind die Ökosysteme in Mitteleuropa wesentlich produktiver und die Landschaften stärker fragmentiert, gleichzeitig ist der menschliche Nutzungsdruck weitaus höher. Ein Beispiel dafür ist die intensive Landwirtschaft, die gepaart mit den hohen Stickstoffeinträgen aus dem Verkehr zu massiven Veränderungen der Lebensraumkapazität führt. Diese Einflüsse sind weitaus stärker als natürliche Prozesse. Das gilt auch für menschliche Störungen, wie Jagd und Erholungsverkehr, die einen größeren Einfluss auf das Verhalten der Huftiere ausüben können als die tierischen Jäger (Ciuti et al. 2012, Mech 2012). Auch in Schutzgebieten ist davon auszugehen, dass sich die indirekten Effekte von den in Nordamerika beobachteten unterscheiden, insbesondere, weil die Schutzgebiete kleiner sind, eine geringere Heterogenität aufweisen und Randeffekte eine große Rolle spielen.
Doch nicht nur die Verteilung der Untersuchungsgebiete ist ungünstig, um Schlüsse auf Mitteleuropa zu ziehen, auch das Untersuchungsdesign ist teilweise problematisch, da die Stichprobenzahlen oft sehr gering sind und keine Wiederholungen vorgenommen werden können. Beispielsweise vergleichen Ripple & Beschta (2006) zwei Canyons im Zion Nationalpark. Der erste Canyon wird stark von Nationalparkbesuchern frequentiert, während der andere nur sporadisch besucht wird. Als Folge kommen im ersten Canyon weniger Pumas vor, was dazu führt, dass mehr Schwarzwedelhirsche auftreten. Dies hat wiederum einen höheren Verbiss zur Folge. Ob letztendlich die unterschiedlich hohen Pumabestände die Ursache für die schlechtere Verjüngung sind, kann bei einem Vergleich von nur zwei Gebieten nicht eindeutig belegt werden, da auch weitere Faktoren von Bedeutung sein können, wie z.B. die touristische Nutzung. Darüber hinaus gibt es auch viele korrelative Studien, die Entwicklungen über die Zeit analysieren. Ein Beispiel dafür sind die Veränderungen im Yellowstone-Ökosystem, die auf die Wiederansiedlung der Wölfe zurückgeführt werden.
Ursächlich können aber auch andere Faktoren sein, die sich unabhängig von der Rückkehr der Wölfe in ähnlicher Weise verändert haben, wie beispielsweise die zunehmende Trockenheit. Diese Problematik kann auch anhand der Entwicklung des Leittriebverbisses im Nationalpark Bayerischer Wald verdeutlich werden. Dort wird seit Beginn der Verbisserhebungen 1987 ein Rückgang des Leittriebverbisses bei der Tanne von 42 auf 10% beobachtet, obwohl der Rehabschuss im gleichen Zeitraum von über 200 Tieren auf 0 zurückgenommen wurde. Parallel zu dieser Entwicklung verlief die Wiederansiedlung und Etablierung des Luchses im Nationalparkgebiet. Damit könnte man den Rückgang des Verbisses auf die Rückkehr des Luchses zurückführen, wenn nicht im gleichen Zeitraum die Massenentwicklung des Buchdruckers stattgefunden hätte, die mehr Äsung und eine höhere Verjüngungsdichte zur Folge hatte. Da beide Entwicklungen parallel abliefen, lässt sich rückwirkend nicht mehr klären, welcher Erklärungsbeitrag der Rückkehr der Luchse und welcher der Borkenkäfermassenvermehrung zukommt.
Tatsächlich haben Großraubtiere selbst dort, wo sie in Europa noch vorkommen oder wieder zurückkehren, oft nur einen geringen Einfluss auf die Ökosysteme. Dies liegt vermutlich nicht am fehlenden Potenzial zur Limitierung ihrer Beutetiere, sondern an fehlender menschlicher Toleranz. Legale und vielerorts auch illegale Abschüsse führen zu einer Absenkung ihrer Bestände, so dass eine Reaktion der Räuber auf ansteigende Beutetierbestände nur eingeschränkt möglich ist (Liberg et al. 2012, Müller et al. 2014). Ein gutes Beispiel dafür war die Situation in den Schweizer Alpen Mitte der 1990er-Jahre. In Folge eines hohen Luchsbestandes gingen die Abschüsse bei den Rehen stark zurück, gleichzeitig stiegen die Schäden bei Nutztieren an. Als Reaktion auf diese Entwicklung nahmen die illegalen Luchsabschüsse stark zu und führten zu einem Rückgang des Luchsbestands (Breitenmoser et al. 2010). Hier hat also der Spitzenprädator Mensch in das System eingegriffen und so den Einfluss der Luchse limitiert. Deshalb ist es auch unklar, ob der von Melis et al. (2009) beobachtete geringere Einfluss von Prädatoren in produktiven Lebensräumen nicht durch andere Faktoren beeinflusst wird, zumal er nicht im Einklang mit den Vorhersagen der EHS steht.
Es bestehen also noch große Wissensdefizite darüber, in welchem Ausmaß große Beutegreifer Beutetierpopulationen und Ökosysteme in Mitteleuropa beeinflussen und ob sie einen signifikanten B
-
Abb. 1: Veranschaulichung der Vorhersagen der „Exploitation Ecosystems Hypothesis“ (EEH) für Ökosysteme mit zwei bzw. drei trophischen Ebenen (nach Oksanen 1992, Oksanen et al. 1981). Dargestellt wird die Dichte der großen Beutegreifer, der Pflanzenfresser und die Schäden an Pflanzen entlang eines Produktivitätsgradienten in Abhängigkeit vom Vorkommen großer Beutegreifer für Ökosysteme mit einer Nettoprimärproduktion von mehr als 0,7kg/m²/Jahr (verändert nach Ripple & Beschta 2012a).Visualisation of the predictions of the ”Exploitation Ecosystems Hypothesis“ (EEH) for ecosystems with two or three trophic levels (according to Oksanen 1992, Oksanen et al. 1981). The figure shows the density of the large predators, of the herbivores and the damages of plants along a gradient of productivity depending on the occurrence of large predators for ecosystems with a net primary production of more than 0.7 kg/m²/year (altered according to Ripple & Beschta 2012a). -
Abb. 2: Der Zusammenschluss zu größeren Gruppen ist eine bewährte Feindvermeidungsstrategie beim Rothirsch. Viele Augen, Ohren und Nasen können die Umgebung dann besser überwachen als ein einzelnes Tier, so dass es für ein Raubtier sehr schwierig wird, sich unbemerkt zu nähern. Darüber hinaus können sich die Huftiere auch beim Sichern abwechseln, so dass sie mehr Zeit für die Nahrungsaufnahme zur Verfügung haben. © Rainer SimonisThe consolidation to larger groups is an established method for the protection against enemies for the red deer. Many eyes, ears and noses can better observe the surrounding than it is possible for the individual animal, which makes it more difficult for a predator to approach unnoticed. Additionally the hoof animals can also alternate with being attentive, which allows them to take more time for foraging. -
Abb. 3: Dort, wo Rehe vorkommen, sind sie die beliebtesten Beutetiere des Luchses. © Rainer SimonisIf roe deer does occur it is the favourite prey of the lynx. -
Abb. 4: Entwicklung von Reh- und Luchspopulation im Wald von Bialowieza (Ostpolen) über einen Zeitraum von mehr als 140 Jahren. Durch die Ausrottung der Luchse um 1900 und in den 1960er-Jahren ergaben sich zwei unbeabsichtigte Experimente, um den Einfluss von Prädatoren zu untersuchen. Die Rehpopulation befindet sich in den Jahren, in denen Luchse im System vorkommen, deutlich unter der Habitatkapazität, die nur in den Jahren erreicht wird, in denen die Luchse ausgerottet waren (verändert nach Jedrzejewska & Jedrzejewski 1998).Development of populations of roe deer and lynx in the forest of Bialowieza (Eastern Poland) over a period of more than 140 years. The extinction of the lynx around 1900 and in the 1960s lead to two unintended experiments to investigate the influence of predators. During the occurrence of the lynx in the system the roe deer population remains clearly under the capacity of the habitat. This capacity was only reached in the years when the lynx was exterminated (Jedrzejewska & Jedrzejewski 1998). -
Abb. 5: Wölfe sind die bedeutendsten Jäger von Huftieren in der nördlichen Hemisphäre. © Rainer SimonisWolves are the most important hunters of hoofed mammals in the northern hemisphere. -
Welche Effekte haben große Beutegreifer auf Huftierpopulationen und Ökosysteme? -
Welche Effekte haben große Beutegreifer auf Huftierpopulationen und Ökosysteme?





Bedienhilfen
Barrierefreiheit
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.