Wildpflanzensaatgut im Spannungsfeld des Naturschutzes
Abstracts
Der Einsatz kommerziellen Wildpflanzensaatguts ist zentraler Bestandteil vieler Kompensationsmaßnahmen. Aus genetischen Studien lassen sich Abstände zwischen Quell- und Zielstandort für Saatgut von mindestens 300km bei ähnlichen Standortfaktoren als zulässig ableiten. Kleinräumig stabil nebeneinander bestehende Ökotypen erlauben den Rückschluss, dass am Ausbringungsort von regionalem Saatgut in den angrenzenden Populationen keine erheblichen Schäden durch Hybridisierung auftreten.
In einer Studie der Universität Hannover wird in einem Artenfilter ein großer Teil der heimischen Arten für Ansaaten ausgeschlossen. Die Autoren dieser Studie wirkten auch bei Rezepturen für 72 Mischungen, den RSM-Regio, als bundesweitem Standard mit. Diese Mischungen bestehen auf Basis des Artenfilters aus wenigen, meist ubiquitären Arten und werden vom Verband deutscher Wildpflanzenproduzenten e.V. (VWW) abgelehnt.
Am Beispiel der Feuchtwiesen wird im vorliegenden Beitrag die mangelnde Differenzierung der RSM-Regio aufgezeigt und der weitreichende Artenausschluss durch den Artenfilter hinterfragt. Der VWW empfiehlt für den Naturschutz artenreiche standortangepasste Mischungen und den Aufbau einer Artendatenbank mit genetischen Informationen für den Saatguteinsatz.
Seeds from Wild Plants from a Nature Conservation Point of View Critical annotations on current suggestions of regulations
The application of commercial seed material of wild plants has become a central component of many compensation measures. Genetic studies allow the derivation of acceptable distances at least 300 km between source and destination of the seed of with comparable site conditions. The fact that adjacent ecotypes firmly coexist side by side allows the conclusion that the application of regional seed material does not cause significant hybridisation effects in adjacent populations. A study at the University of Hannover excludes a large number of native species for seed measures by applying a species filter. The authors of this study also contributed to the recipes for 72 mixtures, the RSM-Regio, which was established as a standard on federal level. Based on the species filter these mixtures consist of few, mostly ubiquitary species. The Association of German Wild Seed and Wild Plant Producers (VWW) has refused their application. Using the example of moist grassland, the study illustrates the lacking differentiation of the RSM-Regio and it questions the exclusion of species. For nature conservation purposes the VWW recommends species-rich mixtures adapted to the site and the establishment of a species data bank providing genetic information for the application of the seed material.
- Veröffentlicht am

1 Vorbemerkung
Regionales Wildpflanzensaatgut wird im Bundesnaturschutzgesetz (BNatSchG; Deutscher Bundestag 2009) in § 39 erstmalig als Mittel für naturschutzfachlich geeignete Maßnahmen genannt. Zu beachten ist, dass dort alle Arten, die sich für mehrere Generationen in Deutschland vermehren, als heimisch und Arten, die außerhalb ihres natürlichen Vorkommensgebietes auftreten, als gebietsfremd bezeichnet werden (§7). Nicht gebietsfremde Arten können im Umkehrschluss nach Kowarik & Seitz (2003) als gebietseigen bezeichnet werden; sie verbinden mit diesem Begriff die Bedingung, dass die betreffende Art im Gebiet auch indigen sein muss. Diese ist allerdings nicht zwingend aus dem BNatSchG ableitbar.
Die Geschichte der Sammlung, Vermehrung und Verwendung von Wildpflanzen zu Ansaatzwecken ist dagegen deutlich älter. Wildgräseranbau tritt in Europa schon im 18. Jahrhundert auf (L.C. Nungesser Saaten 1981). Differenzierte Mischungen z.B. zur Wiesenanlage mit trockenem Boden werden bereits 1883 von der Fa. Conrad Appel (1989) angeboten. Diese Mischungen bestehen aber überwiegend aus Gräsern. Noch bis in die 1960er-Jahre werden große Partien von artenreinen Wildgrassammlungen aus den Haubergen von Taunus, Westerwald und Lahn-Dill-Bergland bei der Fa. Samen Schneider aufbereitet und zur Vermehrung weiterverkauft (R. Schneider, Haiger, mdl. Mitt. 2015).
Mit der im BNatSchG von 1976 erstmalig verankerten Eingriffsregelung (Jessel 2003) wird die breite Anwendung von Wiesenansaaten für Ausgleichs- und Ersatzvorhaben unterstützt. Diese nun kräuterreichen Mischungen gewinnen in den ausgeräumten Agrarlandschaften rasch eine zunehmende Popularität. Durch steigenden Bedarf an Saatgut zur Neuanlage von Blumenwiesen verstärken die wenigen mit Wildpflanzen vertrauten Saatgutfirmen, wie Blauettikett Bornträger, Conrad Appel und die L.C. Nungesser KG, in dieser Zeit den Zukauf von Kräutersamen und legen landwirtschaftliche Vermehrungsflächen an. Heute sind in Deutschland ca. 120 Betriebe mit der Saatgutgewinnung und der Produktion von Einzelpflanzen oder Mischungsprodukten befasst.
Mit der steigenden Zahl der Betriebe, dem zunehmenden Informationsbedarf der Saatgutanwender und den vielen offenen Fragen zu naturschutzfachlichen und saatguttechnischen Belangen wird 2005 der Verband der deutschen Wildsamen- und Wildpflanzenproduzenten e.V. (VWW) zur Bündelung und Bearbeitung dieser Themen gegründet. Neben Beratung bei Gesetzgebung und Verwaltung wird vom VWW mit den Zertifikaten VWW-Regiosaaten® und VWW-Regiogehölze® ein Herkunftsnachweis für Wildpflanzen eingeführt.
Zentrales Thema des Verbandes ist außerdem die Erarbeitung von wissenschaftlichen Grundlagen für den Einsatz von Wildpflanzen. Hierzu wird in einem ersten Vorschlag von Hacker & Hiller (2003) Deutschland in 13 Regionen unterteilt. In einer darauf folgenden Studie wird von Prasse et al. (2010) ausgehend von einem bundesweiten Arbeitskreis eine Gliederung in 22 Regionen vorgenommen. Ein weiterer Teil dieser Studie enthält eine Klassifizierung der Ansaateignung aller Wildpflanzenarten (ohne Gehölze) in einem Artenfilter. Seit 2014 werden von der Forschungsgesellschaft Landschaftsbau und Landschaftsentwicklung (FLL) auf der Basis des Artenfilters die RSM-Regio herausgegeben (FLL 2014): eine Sammlung von konkreten Mischungsrezepturen aus Wildsaatgut, differenziert für 22 Regionen. In Abschnitt 3 des vorliegenden Beitrags sollen unter der Prämisse eines praktikablen Wildsaatguteinsatzes das Regionenkonzept, der Artenfilter und die RSM-Regio kritisch betrachtet werden.
2 Genetische Aspekte
2.1 Ausgangssituation
Neben den massenhaft eingesetzten Zuchtsorten der Landwirtschaft bestehen auch die verbreiteten Landschaftsansaaten jahrzehntelang fast ausschließlich aus Zuchtsortenmischungen, z.B. Berliner Tiergarten-Mischung und Regelsaatgutmischungen (RSM). Seit Mitte der 1970er-Jahre werden erstmals kräuterreiche Mischungen erprobt, die überwiegend der natürlichen Zusammensetzung von Pflanzengesellschaften nachgeahmt sind. Die Präzision, mit der standörtlich angepasste Mischungen mit prozentgenauer Artenzusammensetzung erstellt werden können, führt bei fachgerechter Anwendung zur Etablierung von klassischen Pflanzengesellschaften, die sogar die Merkmale für Lebensraumtypen nach FFH-Richtlinie (Rat der EU 1992) erfüllen. Dabei handelt es sich aber immer nur um Anfangsstadien langfristig zu entwickelnder Lebensgemeinschaften (Joas et al. 2007). Diese neue Machbarkeit im Naturschutz wird von vielen Projektträgern und Naturschutzverwaltungen aufgegriffen und führt zu weitreichenden Ansaatvorhaben mit Wildpflanzen, z.B. Ausgleichsflächen für Flughäfen mit Anlage großflächiger Magerrasen, Begrünung von Autobahnböschungen oder die gezielte Ansaat für Artenschutzprojekte, z.B. von wechselfeuchten Wiesen mit Großem Wiesenknopf (Sanguisorba officinalis) zur Förderung der Ameisenbläulinge (Maculinea spec.) (HMWVL 2005). Das Angebot an Wildpflanzensaatgut umfasst heute in Deutschland Hunderte von Mischungen aus ca. 400 Arten.
Mit dem verbreiteten Saatguteinsatz entsteht eine Debatte über die naturschutzfachliche Zulässigkeit der Saatgutausbringung. Vor allem Fragen zur Genetik stehen dabei im Vordergrund und werden von einer zunehmenden Zahl von Studien beleuchtet. Im Folgenden sollen kurz die beiden wichtigsten Untersuchungsmethoden und die häufigsten Kritikpunkte beim Saatguteinsatz diskutiert werden.
2.2 Untersuchungsmethoden
Die Ergebnisse zahlreicher vorliegender molekulagenetischer Markeruntersuchungen (Allozym, RAPD, SSR, AFLP, cpDNA, SNP) zeigen Differenzierungen am Erbgut zwischen Populationen, die ohne Selektion, also zufällig auftreten (genetische Drift). Merkmale, die einem ähnlichen Selektionsdruck unterliegen, verändern sich dagegen wenig oder gar nicht (Vander Mijnsbrugge et al. 2010). Verpflanzungsexperimente (Common-Garden-Versuche) testen dagegen auch quantitative, selektionsabhängige Merkmale, z.B. die Wuchsleistung, die Fertilität und die Ausprägung relevanter Organe. Sie erfassen aber praktisch nie die Summe der Standortfaktoren, z.B. Witterungsanomalien, wie starke Fröste und Trockenperioden, die über eine lange Zeit zur Ausbildung des lokalen Ökotyps beigetragen haben.
2.3 Biodiversität
Als mögliche nachteilige Effekte für die innerartliche und zwischenartliche Vielfalt durch Saatgutausbringung werden folgende Aspekte diskutiert:
(1) Auftreten invasiver heimischer Arten (kryptische Invasion): Verdrängung örtlicher Sippen durch eingebrachte Sippen oder deren Hybriden infolge von Konkurrenzvorteilen. Zusätzlich kann eine Gefährdung von Artengemeinschaften entstehen, wenn die neue Sippe ihr ökologisches Spektrum erweitert. Das Auftreten invasiver Sippen aus dem heimischen Artenspektrum ist extrem unwahrscheinlich, wenn auch durch neu entstandene Ökotypen möglich. Diskutiert wird das z.B. von Joshi (2015) für die Schlanksegge (Carex acuta), die in die Strandschmielen-Gesellschaft am Bodensee einwandert. Ein Teil der heimischen Arten, die in vom Menschen beeinflusste Standorte vordringen (Apophyten), ist ebenfalls auf dieses Phänomen zu prüfen, z.B. das Einwandern von Schmalblättrigem Weidenröschen (Epilobium angustifolium) in Bergwiesen (Sukopp 2008). Auf kontinentaler Ebene wurde invasives Verhalten bei Schilfrassen (Phragmites australis) beobachtet (Saltonstall 2002). Die genannten Beispiele werden nicht auf Effekte durch Saatgutausbringung zurückgeführt!
(2) Inzuchtdepression: Die Population auf dem neu besiedelten Standort ist durch fehlende genetische Diversität so wenig heterozygot, dass schädliche Allele den Fortbestand der Art an diesem Standort gefährden. Zudem kann die Art wegen fehlender Variabilität nur wenig auf sich ändernde Umweltfaktoren mit Anpassung reagieren.
Dieses ernstzunehmende Phänomen ist für die Saatguterzeugung von Bedeutung. Die Produzenten müssen darauf achten, eine genetische Verengung durch eine zu geringe Anzahl besammelter Standorte oder durch technische Selektion im Anbau zu vermeiden.
(3) Auszuchtdepression: Die Population auf dem neu besiedelten Standort kreuzt sich mit der vorhandenen standörtlichen Population. Die Nachkommen besitzen nicht mehr die am Standort vorteilhaften Eigenschaften und verlieren durch gegenseitige Beeinflussung von Genen (Epistase) und Geneinschleusung (Introgression) Konkurrenzvorteile. Es entsteht im Extremfall ein Verlust an innerartlicher Vielfalt durch Verlust an Ökotypen.
Dieser Vermutung widerspricht (bei Einsatz regionaler Ökotypen) die Tatsache, dass kleinräumig viele (Teil-)Population nebeneinander existieren. Selbst Fremdbefruchter können noch bei erheblichem genetischem Austausch (Genfluss) innerhalb kleinräumiger Skalen isolierte Populationen aufbauen. Mix et al. (2006) zeigen an Ferkelkraut (Hypochaeris radicata) in den Niederlanden einen Abstand von 3,5km, bei dem zwei benachbarte Populationen voneinander als isoliert gelten können. Waser & Price (1991) ermitteln in Befruchtungsversuchen an einer ausdauernden Wiesen-Ritterspornart (Delphinium nelsonii) einen optimalen Abstand von 10m. Näher wachsende Individuen erzeugen Inzuchteffekte, weiter entfernte (30m) bewirken Auszuchtdepression, obwohl die Pflanzen von Hummeln und Kolibris bestäubt werden (Waser & Price 1990). Die kleinräumige Existenz zahlreicher Ökotypen nebeneinander belegt, dass selbst bei Auskreuzung und hohem Genfluss erhebliche Stabilität für die Existenz der Populationen besteht (vgl. auch Ernst 1990). Daher führen neue Genotypen in der Nachbarschaft nicht gleich zur genetischen Vermischung und schon gar nicht zur Verdrängung eines ganzen Ökotyps, da auch die Einbringung, anders als bei landwirtschaftlichen Kulturformen, immer nur lokal und punktuell stattfindet (dazu auch Durka in Prasse et al. 2010: 89ff.). Noch beständiger bleibt die Trennung zwischen Selbstbefruchtern oder sogar apomiktischen Sippen (Gregor 2013). Frankham et al. (2011) stellen sogar das Auftreten von Auszuchtdepression bei kleinräumiger Diversität generell in Frage. Räumlich getrennte Populationen entwickeln demnach bei ähnlichen Umwelteinflüssen auch nach mehreren tausend Generationen oder nach bis zu 500 Jahren Trennung ohne Genfluss keine Auszuchtdepression bei Kreuzung.
Zwar gibt es bisher kaum Untersuchungen zu Auszuchtdepression in situ als bestandsgefährdendem Effekt (Mischalski & Durka 2012), jedoch zeigen die bisherigen Nachweise von Verdrängung lokaler Ökotypen durch massenhafte Ausbringung von Zuchtformen (Fischer & Stohr 2000, Lumaret 1990) oder durch Invasion überregionaler Herkünfte (Saltonstall 2002), dass verdrängende Sippen gegenüber der lokalen Sippe besonders abweichende Eigenschaften erfordern.
(4) Verfälschung von Arealen: Das Verbreitungsgebiet eines Ökotyps kann durch Ansaat vergrößert werden. Vorhandene Arealgrenzen, die z.B. die nacheiszeitliche Wiedereinwanderung nachzeichnen, werden verwischt.
Tatsächlich werden durch menschliche Aktivitäten Pflanzenareale im großen Stil verändert (Seitz et al. 2007), z.B. bei Forstpflanzen, entlang von Verkehrswegen, bei Transporten von Verkippungsmaterial und Einsatz von landwirtschaftlichem Saatgut. Auch durch Einsatz von regionalem Wildpflanzensaatgut können Ökotypen in ein für sie neues Areal gelangen. Die Verwischung von (meist nicht bekannten Arealgrenzen) kann daher als Nachteil für die Forschung betrachtet werden, die Vorteile neu begründeter Biodiversität in der Landschaft (Bosshard & Kuster 2001, Haberreiter 2006) sind dem aber gegenüberzustellen.
2.4 Definition im Bundesnaturschutzgesetz
Es muss davon ausgegangen werden, dass für einen Großteil der Wildpflanzenarten genetische Differenzierung und lokale Anpassungen in voneinander mehr- oder weniger isolierten, auch kleinräumigen Populationen bestehen (Michalski & Durka 2012). Die Abstände liegen dabei zwischen vielen km und wenigen Zehner-Metern (Mix et al. 2006, Waser & Price 1991). Die juristische Einordnung derartiger Populationsstrukturen gemäß Artbegriff im BNatSchG ist aber unklar. Zu beachten ist, dass dort alle Arten, die sich für mehrere Generationen in Deutschland vermehren, als heimisch, und Arten (inkl. Unterarten und Populationen), die außerhalb ihres natürlichen Vorkommensgebietes auftreten, als gebietsfremd bezeichnet werden (§ 7). Nicht gebietsfremde Arten können im Umkehrschluss nach Kowarik & Seitz (2003) als gebietseigen bezeichnet werden; sie verbinden mit diesem Begriff die Bedingung, dass die betreffende Art im Gebiet auch ursprünglich vorhanden (indigen) sein muss. Dies ist allerdings nicht zwingend aus dem BNatSchG ableitbar.
Obwohl die innerartliche Vielfalt als Schutzgut unstrittig ist (z.B. Schumacher & Werk 2010), dürfen lokale, häufig durch Lebensraumfragmentierung entstandene Teilpopulationen nicht dazu führen, die Wiederbesiedlung geeigneter Standorte mit Herkünften aus Nachbarräumen zu unterbinden. Kleinste Populationen könnten dann sogar den Einsatz lokaler Mähgutübertragung verbieten. Der Naturschutz würde damit einen wesentlichen Teil seiner Ausgleichsinstrumente verlieren. Es fehlt eine handhabbare Abgrenzung von Populationen mit eigenem Verbreitungsareal im Sinne des BNatSchG (§7 Abs. 2) und hinreichender eigenständiger Differenzierung gegenüber genetisch erkennbaren Teilpopulationen ohne vergleichbare Bedeutung, z.B. bei genetischer Anpassung in wenigen Generationen (Jump et al. 2008). Dieses gilt natürlich nicht für sehr seltene oder versprengte Arten, deren ggf. reliktisches Auftreten mit hoher genetischer Differenzierung verbunden sein kann und die einzelnen, gezielten Artenhilfsmaßnahmen vorbehalten bleiben müssen (z.B. Lütt 2009).
3 Naturschutzfachliche Vorgaben
3.1 Vorbemerkungen
Die unterschiedlichen Auffassungen im wissenschaftlichen Naturschutz führen zu zwei Herangehensweisen für den Einsatz von Wildpflanzensaatgut:
(a) konservierend: Um eine Florenverfälschung zu vermeiden, soll möglichst keine oder nur eine geringe Verwendung von regionalem Saatgut erlaubt werden. Falls eine Ansaat unvermeidbar ist, sollen möglichst wenige Arten ausgebracht werden, weitere geeignete Arten sollen aus natürlicher Zuwanderung hinzutreten. Zwischen Entnahme und Ausbringung dürfen für viele Arten nur kleinste Entfernungen liegen, z.B. Ausbringung nur auf Gemeindeebene (Zahlheimer 2009). Ansaatinstrumente sind dann Mähgutübertragung oder Wiesendrusch (zwangsläufig begrenzt auf Gebiete, in denen geeignete Spenderflächen und die technische Infrastruktur zur Verfügung stehen; ackerbaulich vermehrtes Saatgut steht auf dieser kleinräumigen Ebene meist nicht zur Verfügung).
(b) gestaltend: Die dagegenstehende Auffassung empfiehlt vor dem Hintergrund ausgeräumter Agrarlandschaften und großflächiger Flächenverluste den Einsatz artenreicher Mischungen zur schnellen Entfaltung großer ökosystemarer Synergien (Bosshard & Kuster 2001, Haberreiter 2006, Schmidt 2014) in weiter gefassten Ausbringungsgebieten. Die potenzielle genetische Belastung wird geringer eingeschätzt als die Summe der Vorteile, die durch neue artenreiche Bestände entstehen.
Beiden Haltungen liegt ein regionales Verständnis für die Verwendung von Wildpflanzensaatgut zugrunde, das aber mit unterschiedlichen räumlichen Skalenweiten arbeitet. Die Verantwortung, keine Arten über ihre natürlichen Areale hinaus zu verbreiten, ist für beide Haltungen gleich.
Auf Initiative von Saatgutproduzenten, Wissenschaftlern und administrativem Naturschutz wurde mit Förderung der DBU von Prasse et al. (2010) in einer Studie ein bundesweites Modell entwickelt, um den Einsatz von Wildsaatgut naturverträglich und praxisnah zu regeln. Der VWW war beratend im Projekt beteiligt, hatte aber keinen Einfluss auf die Methode, die den Ergebnissen zugrunde liegt. Die Studie ist dabei eher dem konservierenden Ansatz zuzuordnen und weist für zwei Bereiche Regelungsvorschläge aus, die hier näher betrachtet werden sollen: Abgrenzung von Regionen (Abschnitt 3.2) und Auswahl von zugehörigen Arten (Artenfilter; Abschnitt 3.3).
3.2 Regionen
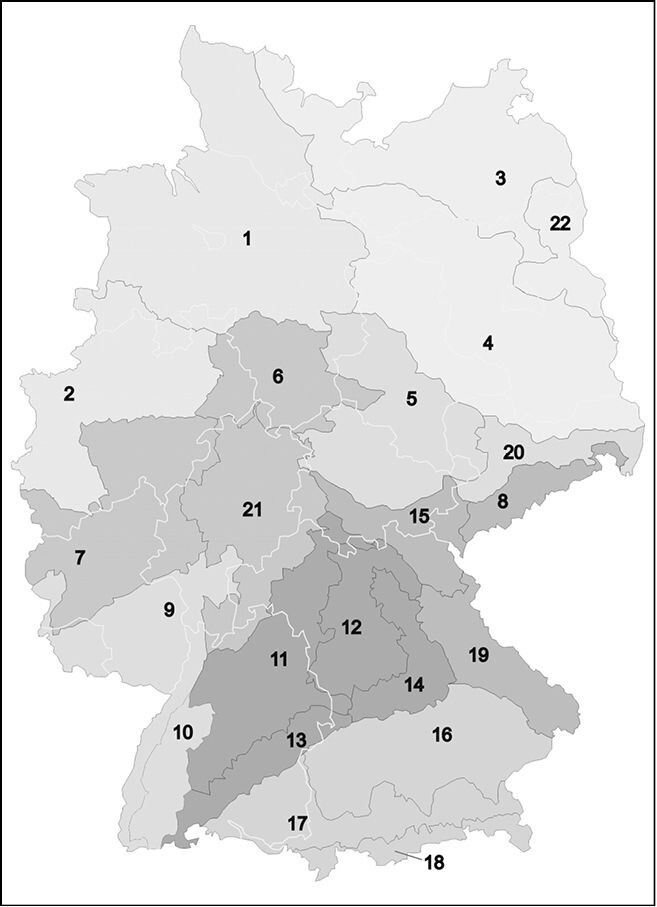
Deutschland wird in der genannten DBU-Studie anhand von klimatischen und bodenkundlich-geologischen Kriterien auf der Basis von aggregierten Naturräumen in 22 Regionen und acht Saatgut-Produktionsräume eingeteilt. Die Regionen sind in ihrer Ausdehnung auf ca. 200km begrenzt (Abgrenzung s. Abb. 4). Sofern darin Ballungsräume mit hohem Nachfragepotenzial auftreten, sind Räume in dieser Dimension zu klein. Landwirtschaftliche Saatgutvermehrung erreicht ohne hinreichenden Saatgutabsatz in Kontinuität und Umfang keine wirtschaftliche Basis oder kann keine ausreichende Palette an Arten zur Verfügung stellen. Der gewählte Maßstab ist auch aus genetischen Erkenntnissen nicht ableitbar. Die meisten Untersuchungen beziehen sich auf großräumige Effekte, z.B. den Vergleich von Ökotypen kontinentaler, subatlantischer und atlantischer Herkünfte (Joshi 2001, Keller & Kollmann 1999, Vander Mijnsbrugge et al. 2010). Dabei deutet sich an, dass im großen Maßstab (Distanzen >500km) mit zunehmender Entfernung die Eignung eines Ökotyps am neuen Standort abnimmt (Leimu & Fischer 2008). Im kleineren Maßstab sind die örtlichen Standortfaktoren, z.B. Beweidung oder Mahd, für eine genetische Differenzierung ausschlaggebender als die räumliche Entfernung, die sich genetisch häufig nicht abbilden lässt (z.B. bei Lauterbach 2012).
Keller et al. (2000) stellen fest, dass im kontinentalen Maßstab Saatguttransfers zu Fitness-Einbußen führen, eine Transfergrenze lässt sich daraus aber nicht definieren. McKay et al. (2005) kommen entsprechend zu dem Schluss: Es ist unmöglich (und kontraproduktiv), eine geographische Standarddistanz als Maßstab für lokale Anpassung vorzugeben (z.B. eine 50-km-Regel). Dementsprechend hätten für Deutschland statt 22 auch 18 oder acht Räume gebildet werden können. Auch die Gewährleistung der Eigenschaft gebietseigen lässt sich nicht über Einhaltung von Räumen ableiten, da die Betrachtung derjenigen Ökotypen, die aus unterschiedlichen Standorten derselben Region stammen (basisch-sauer, feucht-trocken usw.), gänzlich fehlt. Solche Ökotypen können ihre eigenen Regionen aufweisen, z.B. beweidungstolerante Sippen in großräumig von Beweidung geprägten Landschaften, die nicht mit den Grenzen der 22 Regionen übereinstimmten (Abgrenzung s. Abb. 3).
Der VWW betrachtet die acht Produktionsräume als tragfähigen Kompromiss für Saatguttransfergebiete, bei denen die Wahrscheinlichkeit für Schäden an örtlichen Ökotypen sehr gering ist. Er unterstützt aber die Einhaltung der 22 Regionen, soweit dazu Saatgut verfügbar ist, um die Durchdringung von Arealen kleinräumiger Sippen möglichst zu verringern.
3.3 Artenfilter
3.3.1 Funktion
Der Artenfilter wurde als ein Instrument zum Auffinden von geeigneten Ansaatarten in einer Region entwickelt. Er gilt für Anwender, denen keine ausreichenden Kenntnisse über Standort und Arten zur Verfügung stehen. Diesem Ziel wird der Filter gerecht, indem er restriktiv alle Arten ausschließt, deren Verwendung im Einzelfall problematisch sein können. Dabei werden allerdings so viele wesentliche Sippen ausgeschlossen, dass in vielen Regionen nur noch fragmentarische Mischungen möglich sind, die dem Anspruch an Biodiversität und an eine ökologisch fachgerechte Gestaltung kaum mehr genügen. Der Einsatz des Filters stellt sich damit selbst in Frage, da er als ein Instrument zur naturschutzfachlichen Verbesserung gerade das wesentliche Merkmal naturschutzfachlicher Wertigkeit, nämlich regional differenzierten Artenreichtum, ausschließt.
3.3.2 Kritische Ausschlusskriterien
Indigenat: Eingewanderte Arten, die gemäß Konvention seit 1492 als Neophyten bezeichnet werden, sind pauschal nicht als regionales Saatgut zugelassen. Dieses Kriterium wird dem oft jahrhundertelangen Anpassungsprozess vieler eingewanderter und heute etablierter Arten nicht gerecht. Zurzeit sind nur wenige Arten, die diesem Kriterium unterliegen, für Ansaaten relevant, z.B. Nachtviole (Hesperis matronalis). Der Verzicht auf Arten, die in den letzten 100 Jahren eingewandert sind, wäre ausreichend, um die Gefahr invasiver Ausbreitung von Arten zu vermeiden. Die Fokussierung auf Neophyten ist unter dem Aspekt einer potenziellen Verdrängung heimischer Arten ohnehin zweifelhaft. Wie Sukopp (2008) an zahlreichen Arten belegt, können durch geeignete Umstände und genetische Anpassung auch heimische Arten zu Invasoren zuvor artenreicher Gesellschaften werden. Zudem werden vom Bundesamt für Naturschutz alle bekannten invasiven und potenziell invasiven Arten mit laufender Aktualisierung aufgeführt und beschrieben und können so für Ansaaten sicher ausgeschlossen werden.
Gefährdung: Rote-Liste-Arten (ausgenommen Vorwarnstufe) sind von der Verwendung ausgeschlossen, obwohl gerade sie einer Förderung bedürfen und in geeigneten Habitaten neue Lebensräume gewinnen könnten. Viele dieser Arten waren früher weit verbreitet. Mit Berücksichtigung dieser Kenntnisse könnten für zahlreiche Rote-Liste-Arten Ansaaten ermöglicht werden. Einzelne Arten sind bereits in den Sortimenten der Vermehrer vorrätig, die meisten sind aber wegen ihrer geringen Nachfrage in Ansaatprojekten zurzeit nicht als Saatgut erhältlich. Einige Bundesländer haben bewusst dem Ausschluss einzelner Rote-Liste-Arten widersprochen, um eine Förderung zu ermöglichen, z.B. für Sumpfschafgarbe (Achillea ptarmica) in Region 1, Natternkopf (Echium vulgare) in Region 3 oder Heidenelke (Dianthus deltoides) in Region 4. Die besonderen Arten der Kategorien 0, 1, 2 und R in Roten Listen sollten davon aber ausgenommen werden, um sie grundsätzlich nur in speziellen Artenschutzprojekten zu fördern.
Seltenheit und geringe Verbreitung (Arealflächenanteil): Dieses Kriterium schließt die meisten der ansaatrelevanten Arten im Artenfilter aus. Die Erfassungstiefe der floristischen Kartierungen ist zwischen den Bundesländern sehr unterschiedlich. So führen mangelhafte Verbreitungsdaten zum Ausschluss von Arten, obwohl diese eine hinreichend geschlossene Verbreitung aufweisen. Verbreitete Arten, wie die Rundblättrige Glockenblume (Campanula rotundifolia), sind durch den Artenfilter eingeschränkt (nur in 11 von 22 Regionen freigegeben), obwohl die Datenerfassung (heute) ein anderes Bild zeichnet. Hier fehlen zumindest Überarbeitungen, um aufgrund verbesserter Daten aktuelle Einstufungen abzuleiten.
Eine besondere Problematik besteht in regionalen Verbreitungslücken einiger Arten innerhalb ihres Verbreitungsgebietes. Für wärmeliebende Arten können die Hochlagen der Mittelgebirge eine Besiedlungsgrenze darstellen oder säurezeigende Arten fehlen in großen Teilen von Kalkgebieten. In den meisten Fällen führen im Artenfilter solche Verbreitungslücken zum Gesamtausschluss der Art für die betroffene Region. Obwohl also Samen ständig über Wind- oder Tierverbreitung in die Verbreitungslücken eingetragen werden und dort wegen nicht geeigneter Standortfaktoren nicht zur Etablierung gelangen, wird der vergleichbare Vorgang bei einer Ansaat ausgeschlossen.
In Landschaften mit prägender Intensivlandwirtschaft führt der Artenfilter zu der sinnentstellenden Maßgabe, früher häufige Arten nicht mehr ansäen zu dürfen, da sie zu große Areallücken aufweisen.
Natürlich gibt es genügend seltene und sehr seltene Arten mit so geringer Ausbreitungsenergie, dass sie nur noch durch künstliche Ausbringung verbreitet werden könnten. Dieses ist aber nicht das Anliegen der Produzenten im VWW, deren Arbeit vorrangig den häufigen Arten gilt. Bei seltenen oder stärker gefährdeten Arten gelten bei der Saatgutausbringung oder Pflanzung uneingeschränkt die Leitlinien zur Ausbringung heimischer Wildpflanzen (Anonym 1981):
Ausbringung innerhalb des Verbreitungsgebietes,
Saat- oder Pflanzgut von einem nahegelegenen Standort,
Übereinstimmung von Quell- und Zielstandorteigenschaften,
Dokumentation und wissenschaftliche Begleitung der Ausbringung,
Abstimmung mit den zuständigen Landesämtern/-Stellen.
3.3.3 Datenlücken
Bei Abfragen im Datenfilter zeigen sich zahlreiche Datenlücken, die auch den heterogenen Datenquellen der einzelnen Bundesländer geschuldet sein können. Viele Arten werden für einzelne Regionen nicht aufgeführt, obwohl sie dort nachweislich vorkommen. In diesen Fällen ist auch eine Beurteilung, ob die Art als regionales Saatgut geeignet ist, nicht möglich. Zufällig aufgefundene Beispiele sind: Kleiner Odermennig (Agrimonia eupatoria) fehlt für Region 19, Wiesenfuchsschwanz (Alopecurus pratensis) fehlt für Region 12 und 14, Acker-Ochsenzunge (Anchusa arvensis) fehlt in Region 16, Wiesenkerbel (Anthriscus sylvestris) fehlt in Region 10 und 13, Gewöhnlicher Pastinak (Pastinaca sativa) fehlt in Region 3, Bittersüßer Nachtschatten (Solanum dulcamara) fehlt vollständig.
3.4 Die RSM-Regio Praxisempfehlungen der FLL
3.4.1 Ziele
Für die Anwender regionalen Saatgutes ist es i.d.R. schwierig, die richtige Mischung für ihren Standort festzulegen. Hier hilft eine Beratung mit den Saatgutanbietern, die auf die Erfahrung von oft hunderten bereits erfolgter Ansaaten zurückgreifen können. Neben individuellen Mischungen gibt es eine Reihe von auch regional angepassten Standardmischungen bei den Anbietern. Manche Länderbehörden geben Mischungen vor, z.B. für die Anlage von mehrjährigen Blühflächen auf Äckern (HMUKLV 2015, SMUL 2015). Ein besonders weitreichender Versuch, standardisierte Mischungen am Markt vorzuschlagen, sind die RSM-Regio, die seit Frühjahr 2014 von der Forschungsgesellschaft Landschaftsbau und Landschaftsentwicklung (FLL) herausgegeben werden (FLL 2014).
Seit 1978 werden von der FLL (2015) die RSM Rasen jährlich veröffentlicht. Bis heute werden darin Zuchtgräsermischungen wie die RSM 7.1-4 für den Einsatz in der freien Landschaft empfohlen bzw. wird nicht auf deren Anwendungsausschluss in der freien Landschaft hingewiesen, obwohl die Ausbringung von Zuchtformen gegen das BNatSchG (§40 Abs. 4) verstößt und Beispiele zeigen, dass Zuchtformen heimische Formen verdrängen können (Lumaret 1990, Seitz et al. 2007).
Die neuen RSM-Regio mit Wildpflanzen sollen für ingenieurbiologische Zwecke eingesetzt werden und sind geeignet, den Einsatz von Zuchtformen zu reduzieren, indem sie die bisherigen RSM ersetzen könnten. Dabei erhebt das Regelwerk den Anspruch, für alle Regionen in Deutschland Mischungen zu beschreiben, die als Mindeststandard für Begrünungen in der freien Natur für fast alle Standorte geeignet sind. Der VWW hält diese Mischungen bis auf wenige Ausnahmen für nicht geeignet, einen Mindeststandard für naturnahe Begrünungen zu erfüllen, da sie vorwiegend aus ubiquitären, euryöken Arten bestehen und die standörtlichen Möglichkeiten nicht ausschöpfen. Dennoch werden die RSM-Regio immer häufiger über den ihnen zugedachten Zweck hinaus in naturschutzfachlich anspruchsvollen Bereichen eingesetzt, obwohl hier regional abgestimmte, individuelle Mischungen weitaus zielführender sind. Der Einsatz artenarmer Mischungen trägt schließlich auch zu weiteren Lebensraumverlusten für die Fauna bei und verhindert die gesetzliche Funktion dieser Flächen oder erfüllt sie nur in geringem Maße (vgl. z.B. Schmidt 2014).
3.4.2 Differenzierung der Mischungen untereinander
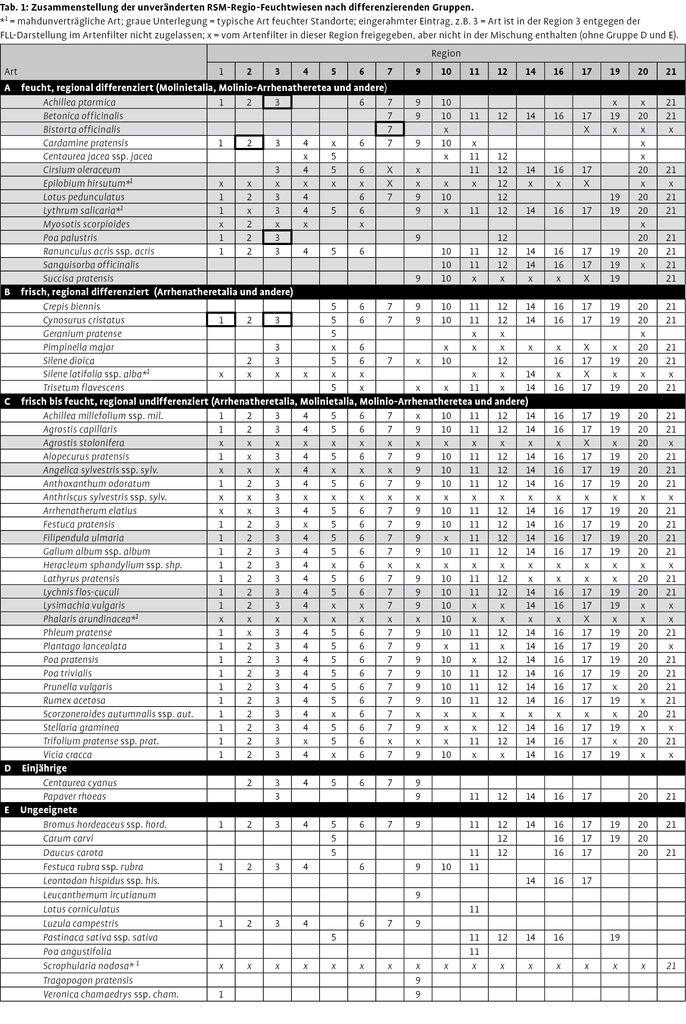
Einer der auffallendsten Aspekte des neuen Regelwerks ist der hohe Differenzierungsgrad der Mischungen. Mit 72 Mischungen für Deutschland wird scheinbar eine große Vielfalt und damit eine Anpassung an die Individualität der Landschaften erreicht. Betrachtet man die Mischungen aber im Detail, so sind nur wenige und kaum regional charakteristische Arten an Differenzierungen beteiligt. Beispielhaft werden alle 17 Mischungen für Feuchtstandorte in Tab. 1 nebeneinandergestellt und nach Artengruppen differenziert (von der FLL wurden nicht für alle 22 Regionen Feuchtstandortmischungen erstellt). Der Eintrag x in den Gruppen A bis D steht für eine Freigabe einer Art in der jeweiligen Region im Artenfilter (Prasse et al. 2010), sie ist aber von den Autoren der RSM-Regio in dieser Region nicht in der Mischung vorgesehen. Vorgesehen sind dagegen die Einträge mit der Nummer der Region.
Die Arten der untersten Gruppe (E) können als ungeeignet für eine Feuchtwiesenansaat eingestuft werden (da im Regelwerk von einer ein- bis dreimaligen Mahd ausgegangen wird, wird hier das Ziel Feuchtwiese angenommen.). Es handelt sich überwiegend um trockener stehende oder sehr konkurrenzschwache Arten, wie Feldhainsime (Luzula campestris), oder stark verdämmende Arten, wie Ausläuferrotschwingel (Festuca rubra ssp. rubra).
Die Gruppe der beiden annuellen Arten (D) beteiligt sich nur im ersten Jahr am Gesellschaftsaufbau und ist für die Betrachtung nicht relevant.
Die Gruppe C enthält 27 Arten, die im Artenfilter für alle 26 Regionen freigegeben sind. Sie können damit nicht zur regionalen Differenzierung zwischen den Mischungen beitragen, auch wenn die Autoren der RSM-Regio durch Auslassungen künstliche Unterschiede zwischen den Mischungen erzeugen; das gilt natürlich auch für die in dieser Gruppe auftretenden Feuchtezeiger. Es handelt sich überwiegend um ubiquitäre Wirtschaftswiesenarten (Molinio-Arrhenatheretea), einige Kennarten aus den Ordnungen der Glatthaferwiesen (Arrhenatheretalia) und aus den Feuchtwiesen (Molinietalia).
Die Gruppe B besteht aus sieben frischezeigenden Arten, die laut Artenfilter aber nicht in allen Regionen eingesetzt werden dürfen. Damit sind zwar regionale Differenzierungen möglich, die Arten sind aber für Feuchtwiesen untypisch, da sie überwiegend der Ordnung der Glatthaferwiesen angehören.
Erst die Gruppe A mit regelmäßig in Feuchtwiesen anzutreffenden Arten, die in einigen Regionen durch den Artenfilter ausgeschlossen werden, erlaubt eine Differenzierung zwischen den Regionen. Allerdings fallen von 14 Arten zwei mahdunverträgliche Arten auf (ebenso wie in Gruppe B + C), die nicht in das Konzept einer Feuchtwiesenansaat passen. Von den übrigen Arten weisen nur acht eine soziologische Zuordnung zu Ordnung oder Verbänden der Feuchtwiesen auf. Dagegen sind Scharfer Hahnenfuß (Ranunculus acris), Wiesenflockenblume (Centaurea jacea ssp. jacea) und Wiesenschaumkraut (Cardamine pratensis) nur Klassenkennarten der Wirtschaftswiesen und zur Charakterisierung von Feuchtwiesen wenig geeignet.
Die acht typischen Feuchtwiesenarten genügen nicht, um 17 regional differenzierte Feuchtwiesenmischungen aufzubauen, obgleich weitere Arten auch bei Anwendung des Artenfilters zur Verfügung stehen würden und auch im Handel erhältlich sind, z.B. Sumpfdotterblume (Caltha palustris), Wolfstrapp (Lycopus europaeus), Sumpfstorchschnabel (Geranium palustre) und Wiesensilge (Silaum silaus).
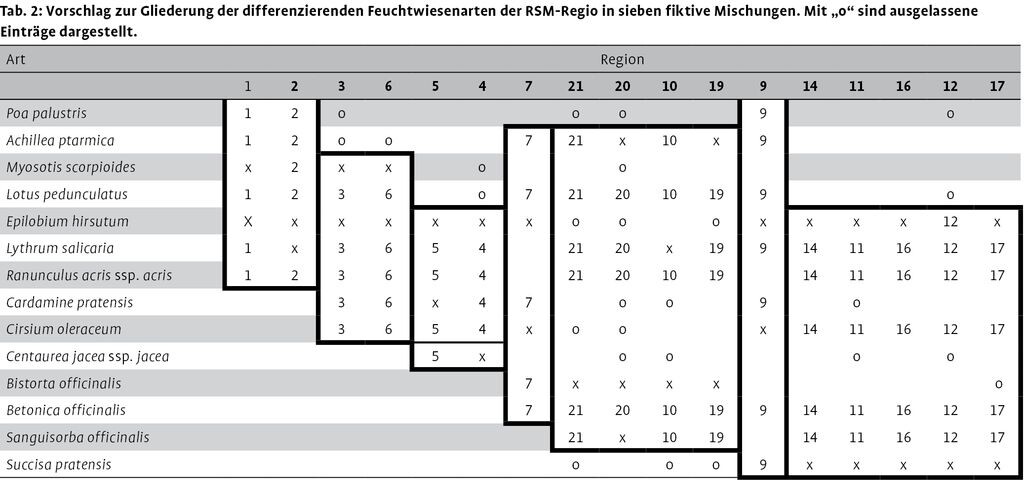
Beispielhaft sind in Tab. 2 mit den Arten der Gruppe A durch Sortierung und Entfernen einzelner Arteinträge sieben Gruppen, bestehend aus einer bis fünf Regionen, gebildet worden. Jede Gruppe enthält nur eine Mischung für die zugehörigen Regionen. Diese Gliederung spiegelt den vorhandenen Artenpool wider und vermeidet eine Scheinvielfalt. Die Darstellung der Tab. 2 ist nicht als realer Entwurf für alternative oder überarbeitete Mischungen zu verstehen, sondern dient zur Erläuterung.
Die RSM-Regio-Feuchtwiesen sollen durch Zusatz von ein oder zwei Brachearten in eine Hochstauden-Ufersaum verwandelt werden, in der Region 10 z.B. nur durch Zusatz von Wasserdost (Eupatorium cannabinum). Diese Wahlmöglichkeit unterstreicht die Scheinvielfalt der Mischungen: Entweder ist die Mischung eine Wiesenmischung, eine Ufermischung oder eine Übergangsmischung; daran ändert auch das Hinzufügen von Wasserdost nichts.
3.4.3 Mischungskonzept
Die Mischungen sind nur wenig nach den natürlichen Zusammensetzungen von Pflanzengesellschaften orientiert und sind widersprüchlich zusammengesetzt. So werden innerhalb des gleichen Standorttyps extreme Magerkeitszeiger und Nährstoffzeiger gemischt, z.B. in der Region 21 beim Feuchtstandort Teufelsabbiß (Succisa pratensis) mit Stickstoffzeigerwert 2 (Ellenberg 1991) und Rote Lichtnelke (Silene dioica) mit 8, was nach der Aussaat zusätzlich zu einem Verlust von Arten führt, je nachdem, ob ein Standort nährstoffreich oder arm ist. Zudem müsste die Beschreibung mager-sauer zu trocken-sauer präzisiert werden (und mager-basisch zu trocken-basisch), da viele der Arten hohe Stickstoff-Zeigerwerte besitzen, z.B. in der Region 14 Wegwarte (Cichorium intybus) miot 5, Gewöhnlicher Pastinak (Pastinaca sativa) mit 5, Gewöhnliches Wiesenrispengras (Poa pratensis) mit 6, Weiße Lichtnelke (Silene latifolia) mit 7.
3.4.4 Artenauswahl
Die Auswahl der Arten in den Mischungen widerspricht häufig den genannten Standorttypen. Die folgenden Beispiele wurden überwiegend anhand der Regionen 5, 9 und 10 ermittelt: Der nässeliebende Sumpfhornklee (Lotus pedunculatus) wird in der Grundmischung der Region 10 eingesetzt, der Basenzeiger Wilder Majoran (Origanum vulgare) in der Region 9 im sauren Bereich. Die Weiße Lichtnelke (Silene latifolia ssp. alba) ist eine Ruderalart nährstoffreicher Standorte und ist für trocken-mager z.B. in der Region 9 nicht geeignet. Die trocken stehende Wilde Möhre (Daucus carota) ist z.B. in der Region 16 im Feuchtstandort aufgeführt. Die Feuchtwiesenart Kuckuckslichtnelke (Lychnis flos-cuculi) wird in der Region 5 in der Grundmischung gelistet. Auch die in Region 5 nur an Magerstandorten vorkommenden Arten Wiesensalbei (Salvia pratensis), Kleiner Wiesenknopf (Sanguisorba minor) und Knöllchensteinbrech (Saxifraga granulata) werden in der Grundmischung aufgeführt. Vom Großen Wiesenknopf (Sanguisorba officinalis) gibt es im Schwarzwald (Region 10) zwei Ökotypen, eine verbreitete, im Juni blühende Form, und eine spätblühende in Tallagen und am Nord- bzw. Ostrand des Schwarzwaldes. Diese müssten getrennt gehandhabt oder besser hier ausgeschlossen werden.
3.4.5 Fehlende Arten
Die Auswahl der Arten für die Mischungen zeigt erhebliche Lücken bei typischen Arten. Wolliges Honiggras (Holcus lanatus) und Gewöhnlicher Hornklee (Lotus corniculatus) fehlen z.B. vollständig in den Mischungen für Region 9 und 10, sind hier aber typisch für frische Wiesen vom sauren bis in den basischen Bereich. Weitere fehlende Arten sind z.B. Wiesenkerbel (Anthriscus sylvestris), Kleiner Klappertopf (Rhinanthus minor) und Ackerwitwenblume (Knautia arvensis). Durch Anwendung des Artenfilters fallen weitere zentrale Arten, wie Wiesenflockenblume (Centaurea jacea ssp. jacea), Rundblättrige Glockenblume (Campanula rotundifolia), Horstrotschwingel (Festuca nigrescens) und Wiesenschaumkraut (Cardamine pratensis), vielfach aus. Auch besonders charakteristische, aber seltenere Arten, wie Knäuelglockenblume (Campanula glomerata) und Karthäusernelke (Dianthus carthusianorum), sind mehrfach im Artenfilter geblockt. Der Artenfilter führt damit insgesamt zu einer gravierenden Einschränkung für naturnahe Mischungen.
3.4.6 Einsatzbereich
Obwohl der Einsatz der neuen RSM-Regio z.B. auf erosionsgefährdeten Böschungen einer standortgerecht zusammengestellten Mischung unterlegen sein muss, empfiehlt das Regelwerk der FLL (S. 19) den Einsatz der neuen Mischungen ausdrücklich vor allem für Begrünungen mit vorwiegend ingenieurbiologischer Sicherungsfunktion. Husicka (2003) und GMU (2004) zeigen dagegen für artenreiche Extensivwiesen die höchste Erosionsfestigkeit anhand der Rheindämme in NRW und am Hochrhein.
Für Ausgleichs- und Ersatzmaßnahmen und naturschutzfachlich ähnlich anspruchsvolle Zwecke soll naturraumtreues Saatgut eingesetzt werden, das nur jeweils innerhalb einer der für Deutschland benannten 502 (!) naturräumlichen Haupteinheiten nach Meynen & Schmithüsen (1962) gewonnen und ausgebracht werden darf. Damit ist naturraumtreues Saatgut als Handelsware praktisch nicht existent. Niemand kann wirtschaftlich Saatgut in so kleinen Räumen produzieren und vorhalten, von einzelnen Vermehrungen für Großprojekte und natürlich Mähgutübertragung und Wiesendrusch, die in der Praxis nur einen geringen Umfang haben, einmal abgesehen. Das FLL-Regelwerk eröffnet daher auch für die RSM-Regio eine Möglichkeit für einen deutlich weiteren Einsatzbereich (S. 19): Wenn kein naturraumtreues Saatgut verfügbar ist [...], kann in Abstimmung mit den zuständigen Fachbehörden auf den nächstbenachbarten Naturraum oder auf Regiosaatgut des entsprechenden Ursprungsgebietes ausgewichen werden.
Damit können die RSM-Regio nicht nur für den begrenzten Bereich ingenieurbiologischer Maßnahmen, sondern für nahezu alle Begrünungen in der freien Natur eingesetzt werden.
4 Empfehlung für die Saatgutanwendung
(a) Zur Vermeidung von Hybridisierung genetisch sehr unterschiedlicher Ökotypen sollten Saatguttransferzonen auf naturräumlich-klimatisch ähnliche Gebiete beschränkt bleiben. Nach Durka (2009) sollte hierzu eine Obergrenze von ca. 300km eingehalten werden. Diesen Ansatz verfolgt der VWW mit den acht Produktionsräumen in Deutschland, deren Längenausdehnung in etwa dieser Dimensionierung entspricht. Aus den vorliegenden Untersuchungen zur Genetik lässt sich eine erhebliche negative Beeinflussung lokaler Ökotypen durch eingebrachte, züchterisch unveränderte Herkünfte desselben Großraums nicht ableiten. Als erheblich sind dagegen die positiven Effekte für die ökosystemare Vielfalt durch naturnahe Begrünungen einzustufen. Die 22 Herkunfts- und Ausbringungsgebiete nach Prasse et al. (2010) werden zur Vermeidung von Gebietsveränderungen von Ökotypen bei vorhandenem Saatgutangebot eingehalten. Hierzu entwickelt sich das Netz der Mitgliedsbetriebe im VWW ständig weiter, da jährlich neue produzierende Betriebe an neuen Standorten hinzukommen.
(b) Die Anwendung des Artenfilters sollte Projekten vorbehalten bleiben, bei denen naturschutzfachliche Anliegen nicht im Vordergrund stehen. Auch bei Begrünungsvorhaben ohne naturschutzfachliche Begleitung erfüllt der Artenfilter eine Sicherungsfunktion. Die restriktiven und zum Teil auch lückenhaften Vorgaben sind nicht ausreichend, um daraus standortangepasste Mischungen zu erstellen. Zudem ist der Artenfilter ohne Fortschreibung ein starres Instrument, das nicht auf neue Erkenntnisse reagieren kann. Alternativ sollte die Auswahl der Arten für eine Mischung nach folgenden Kriterien geprüft werden: Liegt der Ausbringungsort der Art im natürlichen Verbreitungsgebiet? Ist die Art klein- oder großräumig differenziert? Unterliegt die Herkunftsfläche ähnlichen Standortbedingungen wie die Zielfläche? Ist die Art bestimmungskritisch oder genetisch zu variabel? Zukünftig sollte im VWW oder bei einem anderen Träger eine kommentierte und fortzuführende Datenbank erstellt werden, die aus genetischen und sonstigen Vorbedingungen der Arten Hinweise für die Aussaatpraxis enthält.
(c) Aufgrund der Anwendung des Artenfilters unterliegen die RSM-Regio der FLL den beschriebenen Einschränkungen und sollten nicht auf Flächen mit naturschutzfachlichem Anspruch ausgebracht werden. Auch ihr Einsatz zu ingenieurbiologischen Zwecken bleibt kritisch zu hinterfragen. Alternativ sind erprobte oder individuell zu erstellende Mischungen der am Markt vertretenen Wildpflanzenanbieter zu verwenden.
(d) Die Vorgaben für Sammlung und Nachbau müssen den genetischen Erkenntnissen angepasst sein, um Gendrift mit mangelnder Anpassung und Inzuchtdepressionen zu vermeiden.
Literatur
Anonym (1981): Ausbringung von Wildpflanzen Leitlinien zur Ausbringung von Wildpflanzen. Natur und Landschaft 56 (10), 367-369.
Bosshard, A., Kuster, D. (2001): Bedeutung neu angelegter Extensivwiesen für Tagfalter und Heuschrecken.-Agrarforschung Umwelt 8 (7), 252-257.
Conrad Appel (1989): 200 Jahre Conrad Appel Samen und Pflanzen. Roetherdruck, Darmstadt.
Deutscher Bundestag (2009): Gesetz zur Neuregelung des Rechts des Naturschutzes und der Landschaftspflege. BGBl I (51), 06. August 2009.
Durka, W. (2009): Herkunftskontrollen von Saat- und Pflanzgut aus lokalen Herkünften. Machbarkeitsstudie im Auftrag des Instituts für Umweltplanung, Univ. Hannover, unveröff. Mskr.
Ellenberg, H. (1991): Zeigerwerte der Gefäßpflanzen (ohne Rubus). In: Ellenberg, H., Weber, H.E., Düll, R., Wirth, V., Werner, W., Paulissen, D., Zeigerwerte von Pflanzen in Mitteleuropa, 3. Aufl., Scripta geobotanica 18, 9-166.
Ernst, W.H.O. (1990): Mine vegetation in Europa. In: Shaw, A.J., ed., Heavy metal tolerance in plants: evolutionary aspects, CRC Press, Boca Raton, Florida, 21-37.
Fischer, P., Stohr, G. (2000): Neufunde von Arten und Unterarten des Festuca-ovina-Aggregates in Trockenrasen an der Mittleren Elbe und im angrenzenden Gebiet. Mitt. Florist. Kart. Sachsen-Anhalt 5, 167-174.
FLL (Forschungsgesellschaft Landschaftsbau und Landschaftsentwicklung, 2014): Empfehlungen für Begrünungen mit gebietseigenem Saatgut Regiosaatgut-Mischungen, RSM Regio Naturraumtreues Saatgut: Übertrag von Mähgut, Druschgut, Saatgut, Vegetationssoden, Oberboden. Broschüre, Bonn.
(2015): Regel-Saatgut-Mischungen Rasen RSM Rasen 2015. Broschüre, Bonn.
Frankham, R., Ballou, J.D., Eldridge, M.D.B., Lacy, R.C, Ralls, K., Dudash, M.R., Fenster, C.B. (2011): Predicting the Probability of Outbreeding Depression.-Conserv. Biol. 25 (3), 465-475.
Gregor, T. (2013). Apomicts in the vegetation of Central Europe. Tuexenia 33, 233-257.
Haberreiter, B. (2006): Neuanlage von artenreichen Wiesen und Weiden auf ehemaligen Ackerflächen Erfahrungsbericht mit Beispielen aus Niederösterreich. Unveröff. Studie, gefördert vom Land Niederösterreich. Internet: https://www.yumpu.com/de/document/view/10525448/neuanlage-von-artenreichen-wiesen-und-weiden-auf-ehemaligen-/47 (abgerufen am 10.05.2015).
Hacker, E., Hiller, A. (2003): Herkunftsproblematik bei krautigen Arten Regiosaatgut als Perspektive für die Einführung eines bundesweiten Mindeststandards. BfN-Skripten 96, 55-61.
HMUKLV (Hessisches Ministerium für Umwelt, Klimaschutz, Landwirtschaft und Verbraucherschutz, 2015): Hessisches Programm für Agrarumwelt- und Landschaftspflege-Maßnahmen HALM. Internet: https://umweltministerium.hessen.de/sites/default/files/media/hmuelv/richtlinienentwurf_02.04.2015.pdf (abgerufen am 10.05.2015).
HMWVL (Hessisches Ministerium für Wirtschaft, Verkehr und Landesentwicklung, 2005): Planfeststellungsbeschluss vom 22. Dezember 2005 (Neubau der Bundesautobahn 44, Teilabschnitt Hessisch Lichtenau-West Hessisch Lichtenau-Mitte). Internet: http://www.stueer.business.t-online.de/heli/051222.pdf (abgerufen am 10.05.2015).
Husicka, A. (2003) Vegetation, Ökologie und Erosionsfestigkeit von Grasnarben auf Flussdeichen am Beispiel der Rheindeiche in Nordrhein-Westfalen. Dissertationes Botanicae 379, Stuttgart.
Jessel, B. (2003): Die Eingriffsregelung: Bewahrung des Status quo von Natur und Landschaft mittlerweile ein Alltagsgeschäft? Schr.-R. DRL 75, 65-75.
Joas, C., Kiehl, K., Wiesinger, K. (2007): Konzept für naturraumbezogene Ansaaten am Beispiel der Münchner Ebene. Heideflächenverein Münchner Norden e.V., Eching. Internet: http://www.heideflaechenverein.de/service/Artikel_naturraumbezogene%20Ansaaten_070906.pdf (abgerufen am 03.05.2015).
Joshi, J. (2001): Local adaptation enhances performance of common plant species. Ecology Letters 4, 536-544.
(2015): Keimlingsetablierung der invasiven Segge Carex acuta limitierender Faktor bei der Invasion seltener Strandrasengesellschaften? Internet: http://www.uni-potsdam.de/ibb/arbeitsgruppen/ordentliche-professuren/biodiv/stellen/keimlingsetablierung-der-invasiven-segge-carex-acuta.html (abgerufen am 03.05.2015).
Jump, A.S., Penuelas, J., Rico, L., Ramallo, E., Estiarte, M., Martinez-Izquierdo, J.A., Lloret, F. (2008): Simulated climate change provokes rapid genetic change in the Mediterranean shrub Fumana thymifolia. Global Change Biology 14, 637-643.
Keller, M., Kollmann, J. (1999): Effects of seed provenance on germination of herbs for agricultural compensation sites. Agriculture, Ecosystems and Environment 72, 87-99.
, Kollmann, J., Edwards, P.J. (2000): Genetic introgression from distant provenances reduces tness in local weed populations. J. Appl. Ecol. 37, 647-659.
Kowarik, I., Seitz, B. (2003): Perspektiven für die Verwendung gebietseigener (autochthoner) Gehölze. Neobiota 2, 3-26.
L.C. Nungesser KG, Saaten (1981): Samen von Wildpflanzen. Broschüre, Darmstadt.
Lauterbach, D. (2012): Population genetic structure and plant fitness of natural and ex situ populations in Silene chlorantha (WILLD.) EHRH. and Silene otites (L.) WIBEL. Diss. FU Berlin.
Leimu, R., Fischer, M. (2008): A Meta-Analysis of Local Adaptation in Plants. PLoS ONE 3, e4010.
Lütt, S. (2009): (Wieder-) Ansiedlungsprojekte von gefährdeten Pflanzenarten in Schleswig-Holstein. Not. Pflanzenkd. 36 (2), 119-129.
Lumaret, R. (1990): Invasion of natural pastures by cultivated grass (Dactylis glomerata L.) in Galicia, Spain process and consequence on plant-cattle interactions. In: Di Castri, F., Hansen, A. J., Debussche, M., eds., Biological invasions in Europe and the Mediterranean Basin, Monographiae biologicae 65, 392-397.
McKay, J.K., Christian, C.E., Harrison, S., Rice, K.J. (2005): How Local Is Local? A Review of Practical and Conceptual Issues in the Genetics of Restoration. Restoration Ecology 13 (3), 432-440.
Meynen E., Schmithüsen, J. (1962): Handbuch der naturräumlichen Gliederung Deutschlands. Bundesanstalt für Landeskunde, Bad Godesberg.
Michalski, S.G., Durka, W. (2012): Assessment of provenance delineation by genetic differentiation patterns and estimates of gene ow in the common grassland plant Geranium pratense. Conserv. Genet. 13, 581-592.
Mix, C., Arens, P.F.P., Rengelink, R., Smulders, M.J.M., Van Groenendael, J.M., Ouborg, N.J. (2006): Regional gene flow and population structure of the wind-dispersed plant species Hypochaeris radicata (Asteraceae) in an agricultural landscape. Molecular Ecology 15, 1749-1758.
Prasse, R., Kunzmann, D., Schröder, R. (2010): Entwicklung und praktische Umsetzung naturschutzfachlicher Mindestanforderungen an einen Herkunftsnachweis für gebietseigenes Wildpflanzensaatgut krautiger Pflanzen. Univ. Hannover, Abschlussbericht DBU-Projekt (Aktenzeichen: 23931).
Rat der EU (1992): Richtlinie 92/43/EWG des Rates vom 21. Mai 1992 zur Erhaltung der natürlichen Lebensräume sowie der wildlebenden Tiere und Pflanzen. Amtsblatt Nr. L 206 vom 22.07.1992, 7-50.
Saltonstall, K. (2002): Cryptic invasion by a non-native genotype of the common reed, Phragmites australis, into North America. Proc. Nat. Acad. Science USA 99 (4), 2445-2449.
Schmidt, A. (2014): Bedeutung von mehrjährigen, artenreichen Blühstreifen und Feldrainen für Tagfalter und Widderchen Fallbeispiele aus der Agrarlandschaft um Bernburg (Saale). Unveröff. Bachelorarb., Hochschule Anhalt, FB Landwirtschaft, Ökotrophologie und Landschaftsentwicklung, Bernburg.
Schumacher, A., Werk, K. (2010): Die Ausbringung gebietsfremder Pflanzen nach §40 Abs. 4 BNatschG. Natur und Recht 32, 848-853.
Seitz, B., Jürgens, A., Kowarik, I. (2007): Erhaltung genetischer Vielfalt: Kriterien für die Zertifizierung regionalen Saat- und Pflanzguts - Literaturstudie.- BfN-Skripten 208
SMUL (Staatsministerium für Umwelt und Landwirtschaft, Freistaat Sachsen, 2015): Ackermaßnahmen AL. 5c Mehrjährige Blühflächen. Internet: http://www.smul.sachsen.de/foerderung/download/Mischung_fuer_magere_Boeden.pdf (abgerufen am 03.05.2015).
Sukopp, H. (2008): Apophyten in der Flora von Mitteleuropa. Braunschweiger Geobot. Arb. 9, 443-458.
UMG Umweltbüro Grabher (2004): Auswirkung der Bewirtschaftung und der bodenkundlichen Verhältnisse auf die Böschungs-Grasnarbenstabilität der Rheindämme. Internet: http://www.umg.at/projekte/Rheindamm_Stabilitaet.pdf (abgerufen am 03.05.2015).
Vander Mijnsbrugge, K., Bischoff, A., Smith, B. (2010): A question of origin: Where and how to collect seed for ecological restoration. Basic and Applied Ecology 11, 300-311.
Waser, N.M., Price, M.V. (1990): Pollination efficiency and effectiveness of bumble bees and hummingbirds visiting Delphinium nelsonii. Collect. Bot. (Barcelona) 19, 9-20.
(1991): Outcrossing distance effects in Delphinium nelsonii: Pollen loads, pollen tubes, and seed set. Ecology 72, 171-179.
Zahlheimer, W. (2009): Autochthone Begrünung: Grundsätzliches und Aktuelles. Naturschutz in Niederbayern 6, Landshut.
Anschrift des Verfassers: Markus Wieden, Verband deutscher Wildsamen- und Wildpflanzenproduzenten e.V. (VWW), Wetzlarer Straße 11, 35581 Wetzlar, E-Mail info@natur-im-vww.de.
-
Abb. 1: Vermehrungsbestand von Schwarzer Königskerze (Verbascum nigrum) bei Wildsaaten in Wetzlar. Foto: M. Wieden (28.06.2009) -
Abb. 2: Vermehrungsbestand von Bergklee (Trifolium montanum) bei Wildsaaten in Wetzlar. Foto: M. Wieden (26.05.2011) -
Abb. 3: Karte der 22 Ursprungsgebiete für Regiosaatgut. Mit Graustufen sind die Produktionsräume markiert (verändert nach Prasse et al. 2010) -
Abb. 4: Ernte von Wiesenkerbel (Anthriscus sylvestris) bei Wildsaaten in Wetzlar. Foto: A. Wieden (26.06.2011) -
Wildpflanzensaatgut im Spannungsfeld des Naturschutzes -
Wildpflanzensaatgut im Spannungsfeld des Naturschutzes -
Wildpflanzensaatgut im Spannungsfeld des Naturschutzes




Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.