Bedeutung des Biotopverbunds für die Zielart Esparsetten-Widderchen
Die nationalen Verbundkonzepte der Schweiz und Deutschlands werden mit vorliegender Arbeit für die Region Hegau Randen Klettgau konkretisiert. Dieser Raum verfügt aus landesweiter Sicht über einen überdurchschnittlich hohen Anteil an Trockenhabitaten. Nach dem Zielartenansatz wurden mit dem Esparsetten-Widderchen (Zygaena carniolica) sowohl faunistische Gesichtspunkte als auch mit der Gewöhnlichen Küchenschelle (Pulsatilla vulgaris) floristische Aspekte untersucht.
Das Esparsetten-Widderchen wurde als Zielart für die Untersuchung ausgewählt, da diese Art über ein mittleres Ausbreitungsvermögen verfügt und in Landschaften mit fragmentierten Habitaten vorkommt. Die potenziellen Habitate des Widderchens wurden sowohl quantitativ als auch qualitativ untersucht. Verbundraumanalysen ermöglichen Aussagen über die Erreichbarkeit der Flächen für die Arten. Im Gelände wurden die Verbreitung sowie die Habitatqualität für die Art erfasst und anschließend mit einer logistischen Regressionsanalyse ausgewertet.
Für das Esparsetten-Widderchen ist sowohl die Habitatqualität als auch die Konnektivität der Flächen entscheidend. Über eine Distanz von bis zu 750 m findet eine Besiedlung der Flächen am wahrscheinlichsten stattfindet. Für die zweite Zielart Pulsatilla vulgaris hingegen, welche über ein geringes Ausbreitungs- und Wiederbesiedlungspotenzial verfügt, ist die zeitliche Kontinuität der Habitate entscheidend. Zusammenfassend belegt die Untersuchung die Notwendigkeit, den räumlichen und zeitlichen Verbund der Standorte mit hoher Habitatqualität zu sichern.
Significance of Biotope Networks for the Target Species Zygaena carniolica and Pulsatilla vulgaris in the Border-Region Hegau-Randen-Klettgau
Facing an increasing landscape fragmentation, the concept of habitat networks has become more and more important. The national habitat network concepts of Switzerland and Germany are specified on a regional level. From a national perspective the region Hegau Randen Klettgau (Border area in Southwest Germany and Northwest Switzerland) has an above average proportion of dry habitats. Using the target species approach the study investigated faunistic aspects using Zygaena carniolica (blood droplet moth) as well as floristic aspects with Pulsatilla vulgaris (common pasque flower).
Zygaena carniolica was selected as a target species due to its medium dispersal capacity and its occurence in landscapes with fragmented habitats. Based on data of the nature conservation agencies, the potential habitats of Zygaena carniolica were analysed using qualitative and quantitative methods. The connectivity analysis allows statements about the reachability of the patches. In the field, both occurrences and habitat quality for Zygaena carniolica were mapped and analysed using logistic regression. For this species the habitat quality as well as the connectivity is decisive. The distance of 750 metres was identified as a point of reference within which colonization is most likely. On the other hand for Pulsatilla vulgaris, which has a low dispersal and re-colonization capacity, the temporal continuity of the habitats has proven decisive. In conclusion, the study points out the necessity to ensure both spatial and temporal connectivity of patches with a high habitat quality.
- Veröffentlicht am

1 Einleitung
Seit Mitte des 20. Jahrhunderts sind nährstoffarme, trockene Standorte der Offenlandschaft in Mitteleuropa selten geworden. Primäre und sekundäre Trocken- und Halbtrockenrasen sind in ihrem Bestand sowohl durch direkte Flächenverluste, wie z.B. durch Gesteinsabbau oder Aufforstung, als auch durch qualitative Veränderungen infolge von Nutzungsaufgabe oder Nutzungsintensivierung stark gefährdet. Beispielsweise sind im Nordwest-schweizerischen Jura zwischen 1950 und 1997 mehr als 75 % der nährstoffarmen Kalkmagerrasen zerstört worden (Stöcklin & Fischer 2000). Der Schutz der oft sehr kleinen Restflächen reicht für das Überleben der in den Trockenrasen lebenden Arten nicht aus. Aus diesem Grund wurde bereits in den 1970er Jahren die Notwendigkeit eines Biotopverbunds als regionalplanerisches Konzept formuliert (Kaule & Jürging 1977). Seit den 1990er Jahren ist der Biotopverbund mit der Fauna-Flora-Habitat-Richtline (FFH-Richtlinie) auf europäischer Ebene und damit auch in Deutschland mit den Naturschutzgesetzen auf Bundes- und Länderebene rechtlich vorgeschrieben. Ziel des Biotopverbunds ist es nach Jedicke (1994), den Kontakt insoweit wieder zu rekonstruieren, dass er die verbliebenen, voneinander isolierten kleinen Teilpopulationen in die Lage versetzt, sich untereinander auszutauschen. Das heißt, Einzelindividuen der einen Population muss es möglich sein, im Zuge üblicher Ortsveränderungen eine andere Population zu erreichen. Dies verdeutlicht, dass es bei der Funktion des Biotopverbunds um den Austausch von Arten zwischen Flächen geht. Die räumliche Anordnung der Flächen, der strukturelle Verbund, liefert lediglich die Kulisse für den Austausch von Individuen. Konkrete Aussagen zum Biotopverbund sind nur über den Habitatverbund für bestimmte Arten möglich. Doch was bedeutet dies für die Naturschutzpraxis? Wie kann der Biotopverbund umgesetzt werden? Für welche Tier- und Pflanzenarten ist er von Bedeutung? Mit der vorliegenden Untersuchung wurden Fragen zur Bedeutung von Flächengröße, Isolation und Habitatansprüchen von Arten, welche bereits in den 1990er Jahren bearbeitet wurden, wieder aufgegriffen (Amler et al. 1999).
In einem Workshop zum länderübergreifenden Biotopverbund, der mit Teilnehmern aus der Naturschutzverwaltung und -forschung der Schweiz, Frankreich und Deutschland im März 2005 in Basel stattfand, wurde die deutsch-schweizerische Grenzregion Hegau Randen Klettgau als prioritär für den Verbund von Trockenbiotopen identifiziert. Diese Region ist in den nationalen Verbundkonzepten der Schweiz und Deutschlands als potenzieller Trockenhabitatverbund ausgewiesen (Berthoud et al. 2004, Fink et al. 2005). In dem vorliegenden Beitrag werden die landesweiten Verbundkonzepte im regionalen Maßstab unter Berücksichtigung floristischer und faunistischer Aspekte konkretisiert, wobei der Schwerpunkt auf den Besonderheiten der Region liegt und sich die Darstellung weitgehend auf die Vorgehensweise für eine Falterart beschränkt, für die der räumliche Biotopverbund von großer Bedeutung ist. Die Pflanzenart wird nachrangig behandelt, da die Ergebnisse zeigten, dass der räumliche Verbund hier von geringerer Bedeutung ist.
2 Untersuchungsgebiet
2.1 Überblick
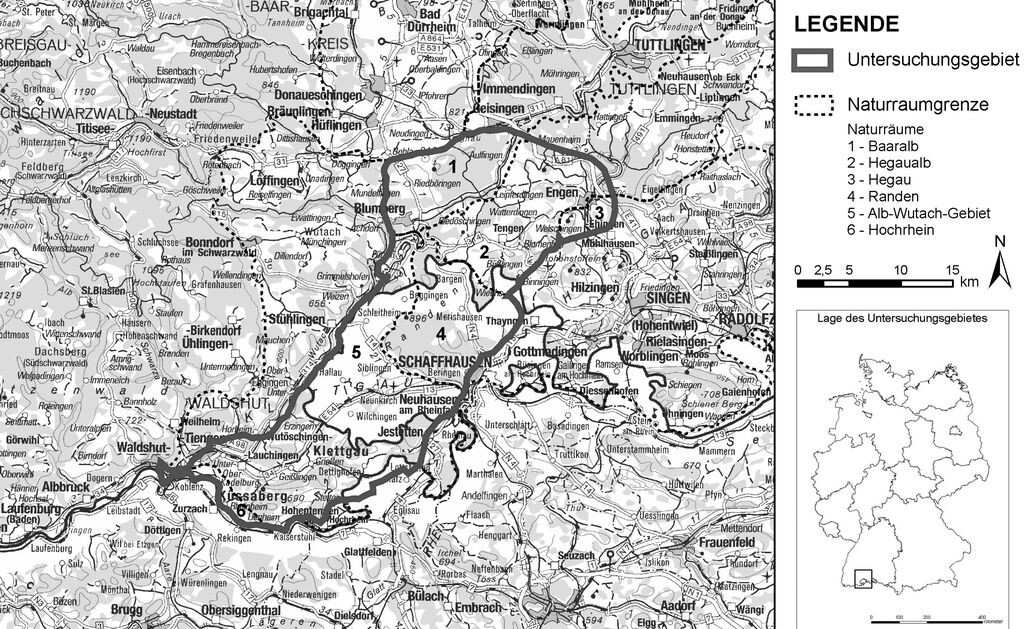
Das Untersuchungsgebiet (Abb. 1) umfasst den nordwestlich von Schaffhausen gelegenen Schweizer Randen und schließt Richtung Nordosten bei Engen an die Ausläufer der Schwäbischen Alb an. Nach Süden wird es durch den Rhein begrenzt.
Großräumig betrachtet liegt das Untersuchungsgebiet in dem Bogen des anstehenden Juragesteins, das sich vom Französischen über den Schweizer bis hin zum Schwäbisch-Fränkischen Jura erstreckt. Im Untersuchungsgebiet überwiegen die Gesteinsschichten des Weißen Jura, welche stellenweise von tertiären und quartären Schichten überlagert sind. Der Randen gehört geologisch zu der Weißjura-Tafel der Schwäbischen Alb. Mit der Einführung der Dreifelderwirtschaft wurde im 19. Jahrhundert der Ackerbau auf den gering rentablen Hochflächen des Randens aufgegeben. Es entstanden lichte Föhrenwälder im Wechsel mit mageren Trockenwiesen, während in der Nähe der Dörfer die Landwirtschaft intensiviert wurde. Die auf den ehemaligen Ackerflächen entstandenen Trockenrasen waren für ein Jahrhundert die typischen Heuwiesen und bedeckten eine große Fläche. Seit den 50er Jahren des 20. Jahrhunderts führte die Intensivierung der Landwirtschaft zu einem erheblichen Rückgang der charakteristischen Trockenarten (Keel 1995, Zoller 1958). Im Gegensatz zum Randen war im Hegau und der Hegaualb die Schafbeweidung weiter verbreitet und die Nutzung der Landschaft heterogener.
2.2 Trockenrasen im Untersuchungsgebiet
Für die Untersuchung des Verbunds von Trockenstandorten besteht die Notwendigkeit, den Begriff der Trockenrasen relativ weit zu fassen. Da die ehemals nur in den Volltrockenrasen vorkommenden Arten aufgrund der Landschaftsentwicklung heute teilweise auch in den trockenen Wiesen des Wirtschaftsgrünlands vorkommen, ist es aus naturschutzfachlicher Sicht sinnvoll, die mageren Wiesen bei den Trockenrasen mit einzubeziehen. Daher sind Gegenstand der vorliegenden Untersuchung die unter dem Begriff Trockenrasen zusammengefassten, trockenen und nährstoffarmen Standorte des Offenlandes (Beispiele in Abb. 2). Primäre, natürliche Trockenrasen-Biotope sind in Mitteleuropa nur kleinflächig ausgeprägt, während die sekundären Trockenrasen als Ergebnis historischer Bewirtschaftungsweisen früher oftmals landschaftsprägend waren.
Die Trockenrasen im Untersuchungsgebiet (Abb. 2) sind relativ niederwüchsige und mehr oder weniger lückige, meist sonnen-exponierten Rasen auf flachgründigen, kalkhaltigen und trockenen Böden. Den flächenmäßig größten Anteil an den Trockenrasen im Untersuchungsgebiet haben die sekundären Halbtrockenrasen (Mesobrometen). Pflanzensoziologisch sind die Trockenrasen des Schweizer Juras und Südbadens u.a. bei Zoller (1954, 1958) und Witschel (1980) beschrieben. Nach Witschel wird die floristische Zusammensetzung der Halbtrockenrasen hauptsächlich durch die Art der Bewirtschaftung bestimmt, entweder der Beweidung oder Mahd oder einer Kombination der beiden. Während Teile des Hegaus und des Alb-Wutach-Gebietes im Bereich der traditionellen Wanderschafhaltung liegen (Jacobeit in Beinlich 1995), ist für den Randen die Wiesennutzung typisch. Im Kanton Schaffhausen wurde 1937 lediglich 1 % der futterbaulichen Fläche als Weideland bewirtschaftet (Kummer 1937), während die Heuwiesen weite Teile der Hochfläche einnahmen. Nach den Ergebnissen von Bewirtschaftungsexperimenten in den Halbtrockenrasen des Randens hat die Bewirtschaftung nur einen geringen Einfluss auf die Artenzahlen und das Artenspektrum (Keel 1995, Langenauer et al. 2000). Weitere Ergebnisse der Untersuchungen zeigen, dass seit Mitte der 50er Jahre die Verhältnisse etwas stickstoffreicher sind und der Gefährdungsgrad der Arten der regelmäßig, aber spät gemähten, sehr nährstoffarmen und (wechsel-)trockenen Flächen noch stärker zugenommen hat (Keel 1995, Zoller 1954).
Die Reliktföhrenwälder stellen eine Besonderheit im Untersuchungsgebiet dar. Da sie auf primären Standorten vorkommen, können sie als Zeuge eines früher weiter verbreiteten Vegetationstyps angesehen werden. Da der Großteil des Untersuchungsgebiets im Würm eisüberdeckt war, stocken die meisten Reliktföhrenwälder auf einst vergletscherten Böden. Die Kiefernbestände und die begleitenden Pflanzenarten sind also in dem Sinne Relikte, dass sie seit ihrer postglazialen Etablierung nicht der Konkurrenz anderer Arten gewichen sind, sondern überdauert haben. Charakteristisch für die primären Trockenstandorte ist, dass sie einen mosaikartigen Vegetationskomplex aus lückigen Grasfluren, Felstriften, Trockenrasen, thermophilen Saum- und Gebüschgesellschaften und lichten Trockenwäldern bilden. Für diesen Vegetationskomplex prägte Gradmann den Begriff der Steppenheide.
3 Zielartenansatz und Auswahl der Zielart
Da eine Bearbeitung aller Arten praktisch nicht möglich ist, werden im Naturschutz häufig einzelne Arten ausgewählt. Der Zielartenansatz ist für die Bearbeitung des Biotopverbunds geeignet, da mit ihm komplexe Sachverhalte artspezifisch dargestellt werden können und gleichzeitig konkrete Anforderungen an die Landschaftsentwicklung abgeleitet werden können. Hypothetisch sollen durch den Schutz der ausgewählten Arten, welche die empfindlichsten und schutzbedürftigsten der relevanten Anspruchstypen umfassen, weitere Arten mit gesichert werden. Dahinter steht der Ansatz, durch die Förderung ausgewählter Arten einen möglichst großen Teil der gesamten biologischen Vielfalt zu erhalten (Reck 2004). Für die praktische Bearbeitung ist es von Vorteil, wenn die untersuchten Arten nicht zu selten vorkommen. Für die ausgewählten Zielarten wurde überprüft, inwieweit der überörtliche Austausch zwischen den Populationen in der fragmentierten Landschaft noch möglich ist.
Als Zielart wurde das relativ flugfreudige Esparsetten-Widderchen (Zygaena carniolica) ausgewählt, welches als eine gute Indikatorart für Trockenrasen gilt (u.a. Hofmann 1994). Für das Esparsetten-Widderchen als einen Vertreter der Artengruppe der flugfähigen Insekten mit einer aktiven Ausbreitung ist die Bedeutung überregionaler Korridore generell als hoch einzuschätzen (Reck et al. 2004). Ein weiteres Kriterium für die Auswahl des Esparsetten-Widderchens war die Unverwechselbarkeit der Art und die relativ einfache Erfassung der Imagines. Die Gefährdung zusammen mit der guten Kenntnis über die Ökologie vom Esparsetten-Widderchen sprechen zudem dafür, dass es sinnvoll ist, diese Zielart zu verwenden. Die vorliegende Untersuchung verfolgt den Ansatz, über die Vorkommensverteilung des Esparsetten-Widderchens auf die funktionalen Beziehungen zwischen den Trockenrasenfragmenten zu schließen.
Charakterisierung des Esparsetten-Widderchens (Zygaena carniolica)
Taxonomisch zu den Nachtfaltern gehörend, sind die tagaktiven Rotwidderchen mit der rot-schwarzen Färbung der Imagines und der gelb-schwarzen Färbung der Raupen sehr auffällige Arten. Von den im süddeutschen Raum vorkommenden Rotwidderchenarten ist das Esparsetten-Widderchen (Abb. 3) die Art mit den extremsten Ansprüchen und einer geringen Standortsamplitude (Wagner 2006) und sie ist daher als Zielart für Trockenrasen besonders gut geeignet. Ein eigentümliches Verhalten vom Esparsetten-Widderchen ist das Bilden von so genannten Parkstationen, bei denen am Spätnachmittag oft fünf, zehn oder mehr Falter an einer Blüte sitzen (Hofmann 1994, SBN 1997).
Die Raupen vom Esparsetten-Widderchen ernähren sich oligophag von der Futter-Esparsette (Onobrychis viciifolia) und dem Gewöhlichen Hornklee (Lotus corniculatus). Die Falter saugen bevorzugt an blauvioletten Blüten, wie die Tauben-Skabiose (Scabiosa columbaria), Wiesen-Knautie (Knautia arvensis), Wiesen-Flockenblume (Centaurea jacea) und Skabiosen-Flockenblume (Centaurea scabiosa). Pflanzensoziologisch sind die Verbreitungsschwerpunkte der Larvalhabitate des Esparsetten-Widderchens den Halbtrockenrasen (Mesobrometen) und Übergangsbereichen zum Volltrockenrasen (Xerobrometen) zuzuordnen. Die Imaginalhabitate leiten mit dem Vorkommen der Falternahrungspflanzen zu den Saumgesellschaften über (Hofmann 1994). Die aus Wiederfangversuchen ermittelten Flugweiten der Falter liegen bei 200 300 m für einen Aktionsradius, in dem sich fast alle Individuen bewegen. Eine zweite Häufung der Flugweiten liegt bei 800 1000 m, während bei einzelnen Individuen Extremwerte von bis zu 5 km nachgewiesen wurden (Hübner 2002, Kreusel 1999, Smolis und Gerken 1987).
Das Esparsetten-Widderchen steht auf der Roten Liste der gefährdeten Tierarten. In Baden-Württemberg ist die Art mit dem Gefährdungsgrad 3 (gefährdet) eingestuft. Die Gefährdung besteht hauptsächlich aufgrund ihrer Abhängigkeit von den stark gefährdeten Trocken- und Halbtrockenrasen (Hofmann 1994, SBN 1997).
4 Methode
4.1 Auswertung der vorliegenden landesweiten Kartierungen der Trockenrasen
Wichtigste Datengrundlagen für die Bearbeitung des Trockenrasenverbundes sind für Baden-Württemberg die überwiegend in den Jahren 1994 bis 1996 durchgeführte §-32-Kartierung und die Waldbiotopkartierung (WBK). Für die Schweiz bildet die bundesweite Erhebung der Trockenwiesen und -weiden (TWW) aus dem Jahr 2004 die Datengrundlage.
Ausgehend von den Habitatansprüchen der Zielart werden von der landesweiten §-32-Kartierung und Waldbiotopkartierung Baden-Württembergs die Biotoptypen der Magerrasen und ihrer Säume berücksichtigt. Aus dem TWW-Projekt fanden die als echte Halbtrockenrasen (Mesobrometen) kartierten Flächen unterschiedlicher Ausprägung Eingang, d.h. auch solche mit Ruderalzeigern und Anteilen von trockenen Saumgesellschaften, während Flächen mit Fettzeigern für die Zielarten weniger geeignete Habitate darstellen und daher nicht mit einbezogen wurden.
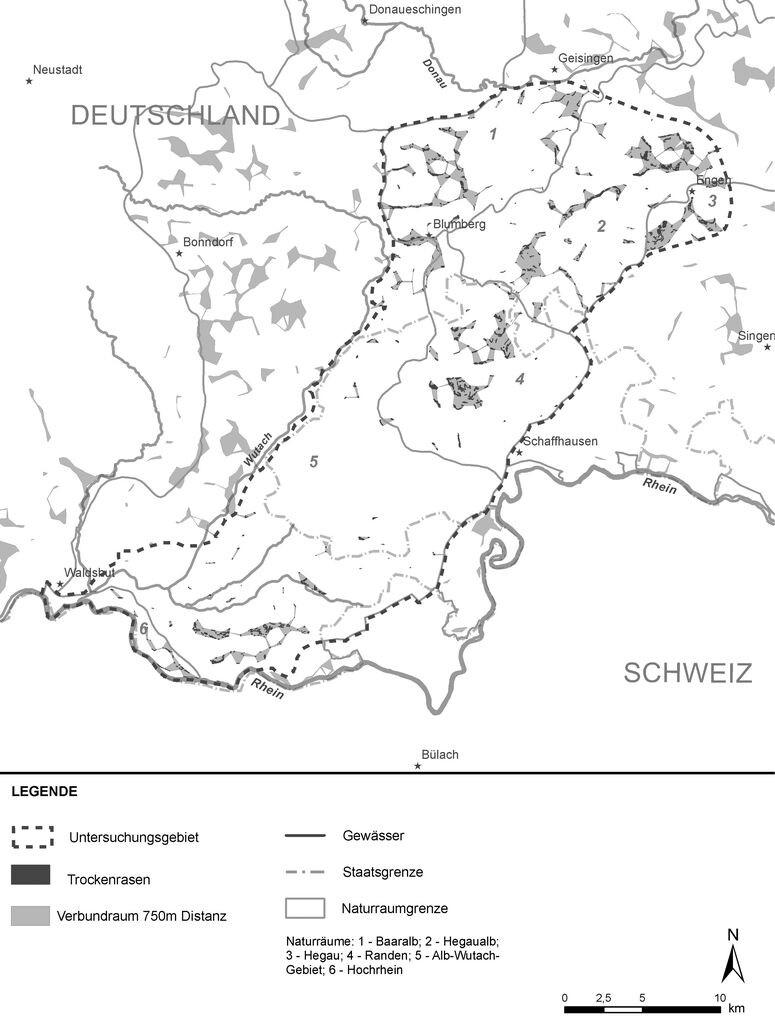
Obwohl aus landesweiter Sicht im Untersuchungsgebiet ein Schwerpunkt der Trockenrasen-Vorkommen liegt, nehmen sie weniger als 0,5 % der Fläche im Untersuchungsgebiet ein, nämlich 298 ha, verteilt auf 849 Teilflächen. Während in den Gebieten der Baaralb, Hegaualb, Hegau, Randen und dem südlichen Alb-Wutach-Gebiet verbreitet Trockenrasen vorkommen, fehlen diese in dem Gebiet zwischen Randen und Klettgau heute weitgehend. Die Flächen im Hegau inklusive der Hegaualb sind im Schnitt deutlich kleiner als die in der Baaralb oder im Randen.
4.2 Rasterkartierung des Esparsetten-Widderchens
Die Erhebung des Esparsetten-Widderchens erfolgte zunächst mit einer Präsenz-Absenz- Übersichtskartierung auf Rasterbasis. Die Kartierung wurde von Kissling im Jahr 2005 durchgeführt. Einen ersten Überblick auf Basis der Quadranten der TK 1 : 25 000 liefert das Grundlagenwerk des Artenschutzprogramms Baden-Württemberg (Ebert 1994). Ziel der Übersichtskartierung ist es, innerhalb einer Flugzeit, welche ca. fünf Wochen dauert (Hofmann 1994), einen Überblick über die aktuelle Verbreitung des Esparsetten-Widderchens im Untersuchungsgebiet zu bekommen. Da am Anfang und Ende der Flugzeit weniger Individuen anzutreffen sind, beschränkt sich der Kartierungszeitraum auf etwa drei Wochen der Hauptflugzeit. Das Untersuchungsraster besteht aus den Sechszehntel Quadranten der TK-25-Blattschnitte, wodurch sich eine Kantenlänge der einzelnen Raster von ca. 3 km ergibt. Das Untersuchungsgebiet wurde damit in 98 Rasterzellen unterteilt. Die Auswahl der Rasterzellengröße orientiert sich an der in einer Flugzeit erfassbaren Menge an Rasterzellen. Auch wenn eine weitere Unterteilung des Rasters aus Sicht der durchschnittlichen Flugweiten vom Esparsetten-Widderchen sinnvoll wäre, wurde darauf verzichtet, da sonst die Bearbeitung des gesamten Untersuchungsgebietes in einem Jahr nicht möglich gewesen wäre.
In den potenziellen Habitaten wurden die Blüten der Falternahrungspflanzen, insbesondere Tauben-Skabiose (Scabiosa columbaria), Wiesen-Knautie (Knautia arvensis) und Wiesen-Flockenblume (Centaurea jacea), nach imaginalen Esparsetten-Widderchen abgesucht. Die Dauer einer Begehung betrug maximal 30 min (Hermann 1992). Konnte bis dahin kein Vorkommen festgestellt werden, wurde für die Untersuchungsfläche ein Nicht-Vorkommen angenommen. Da das Esparsetten-Widderchen eine sehr auffällige Art ist und die Begehungen ausschließlich in der Hauptflugzeit durchgeführt wurden, kann davon ausgegangen werden, dass die Fehlerquote durch übersehene Individuen sehr gering ist (Hafner, Hermann mdl.). Hinzu kommt, dass die Trockenrasen im Untersuchungsgebiet zum Großteil sehr kleinflächig sind, so dass sie innerhalb einer kurzen Zeit vollständig abgelaufen werden können. Für den Fall, dass ein Nachweis des Esparsetten-Widderchens erfolgte, blieben die anderen potenziellen Habitate des Rasters unberücksichtigt. In der Zeit vom Ende Juli bis Mitte August 2005 wurden insgesamt 153 Flächen aufgesucht, auf denen in 33 Fällen das Esparsetten-Widderchen vorkam. Die Rasterkartierung zeigte, dass das Esparsetten-Widderchen besonders im Norden und Süden des Untersuchungsgebietes verbreitet ist. Die ökologische Spannweite der Flächen, in denen das Esparsetten-Widderchen vorkommt, reicht von Halbtrockenrasen trockener und typischer Ausprägung bis zu den mesophileren Salbei-Glatthaferwiesen. Damit ist das Esparsetten-Widderchen in der Region nicht streng stenök, sondern besiedelt insbesondere in den wärmebegünstigten Räumen ein relativ weites Spektrum an Biotoptypen. Da das Untersuchungsgebiet im Verbreitungszentrum der Art liegt, sind die schon von Hofmann (1994) beschriebenen euryöken Eigenschaften nicht außergewöhnlich.
4.3 Konnektivitätsanalysen
Konnektivitätsanalysen sollen Aussagen über die Erreichbarkeit der Flächen für einzelne Arten ermöglichen. In der vorliegenden Untersuchung wurde durch die Analyse des Vorkommens des Esparsetten-Widderchens der Aktionsradius der Art eingegrenzt. Mittels eines Geographischen Informationssystems (GIS) wurde über die Modellierung der räumlichen Daten die Verbundsituation der Trockenrasen visuell dargestellt. Den in der Literatur erwähnten Wanderungsdistanzen des Esparsetten-Widderchens zufolge wurden mit dem Verfahren der Verbundraumanalyse (Jooss 2006) für das Untersuchungsgebiet für Distanzen von 0 bis 1 000 m im Abstand von 25 m Verbundräume um die Trockenrasen erzeugt. Das von Jooss entwickelte Verfahren der Verbundraumanalyse verbindet alle Flächen (Patches), die näher als ein bestimmter Schwellenwert beieinander liegen, zum kleinstmöglichen Umrisspolygon mit Eckpunkten in den enthaltenen Flächen. Mit einer logistischen Regressionsananlyse wurde die Verbunddistanz von 750 m als der Wert ermittelt, welche das Vorkommen des Esparsetten-Widderchens am besten erklärt.
4.4 Detailerhebung zur Habitatqualität für das Esparsetten-Widderchen
In dem der Rasterkartierung folgenden Jahr wurde für ausgewählte Flächen die Habitatqualität für das Esparsetten-Widderchen erhoben. Die Kartierung verfolgt das Ziel, für Flächen von unterschiedlicher Flächengröße und Konnektivitätsgraden die Habitatqualität zu erfassen, um anschließend mit statistischen Auswertungen die Bedeutung der Einflussfaktoren auf das Vorkommen des Esparsetten-Widderchens zu ermitteln. Da bei Untersuchungen im regionalen Maßstab eine flächendeckende Bearbeitung nicht möglich ist, erfolgte eine Flächenauswahl mit einer geschichteten Zufallsstichprobe. Das Verfahren der geschichteten Zufallsstichprobe stellt sicher, dass unterschiedliche Gruppen von Flächen berücksichtigt werden, wobei keine weitere Vorauswahl getroffen wird, welche das Ergebnis beeinflussen würde. Die Auswahl der Stichproben erfolgte innerhalb der nach den Kriterien Verbundenheit und Flächengröße getrennten Gruppen. Dabei gelten als gut verbunden diejenigen Flächen, welche maximal eine Distanz von 800 m zum nächsten potenziellen Habitat aufweisen.
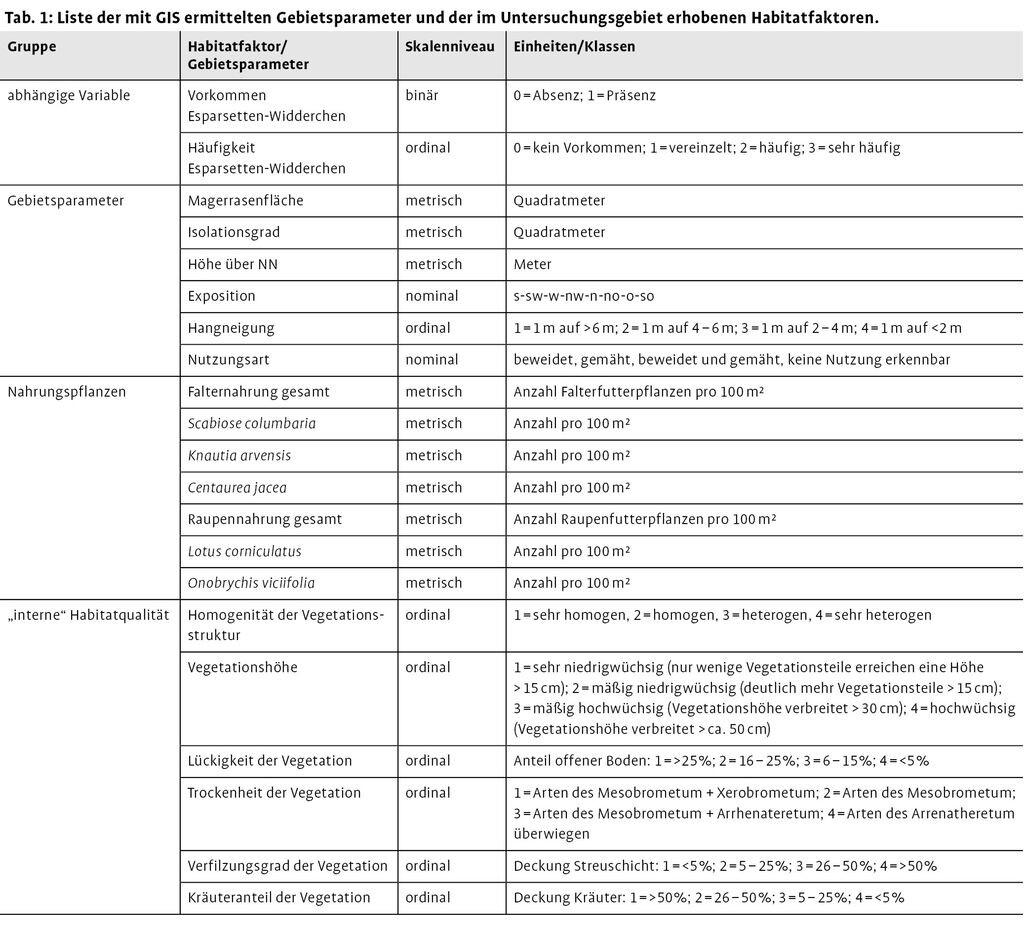
Für die Habitatqualität wurden die für das Esparsetten-Widderchen in der Literatur als relevant erachteten Umweltparameter mit einem Erhebungsbogen kartiert. Die Erhebungsmethode orientiert sich an Wagner (2002, 2006) und dem Wissen von Experten (Hafner, Hermann mdl.). Der Großteil der für die Art relevanten Gebietseigenschaften kann über die Vegetation erfasst werden. Die Dichte der wichtigsten Raupen- und Falternahrungspflanzen wurde über die Anzahl der Blütenköpfe notiert. Weitere Vegetationsparameter sind Vegetationshöhe, Trockenheit/Magerkeit, Lückigkeit, Verfilzung und Kräuterreichtum der Flächen. Hier wurde eine vierstufige Skala verwendet, wobei die Anordnung der Wertstufen so erfolgte, dass die Wertstufe 1 jeweils die optimale Bedingung für das Esparsetten-Widderchen darstellt. Die floristische Artenausstattung der Flächen wird über typische Vertreter von Artengruppen, die als Zeigerarten für bestimmte Standorteigenschaften stehen, erfasst. Die im Gelände erkennbare Nutzung wird in Beweidung und Mahd unterschieden. In Tab. 1 sind die weiteren im Gelände erhobenen Parameter aufgeführt.
Von Ende Juli bis Mitte August (28.07. bis 17.08.2006) wurden von den 849 Trockenrasenflächen insgesamt 108 Flächen erfasst, davon 80 zufällig ausgewählte Flächen der unterschiedlichen Größen- und Isolationskategorien und 28 Flächen, auf denen das Esparsetten-Widderchen im Jahr 2005 nachgewiesen wurde. Mit einem Stichprobenanteil von über 12 % sind die Ergebnisse statistisch abgesichert.
4.5 Statistische Habitatmodellierungen
Das Vorkommen und die Abundanz des Esparsetten-Widderchens stehen in einem engen Zusammenhang mit den Habitateigenschaften der Trockenrasen (vgl. Wiens 2002). Das Ziel von Habitatmodellierungen ist, die Beziehung zwischen den Umweltbedingungen und den Habitatansprüchen von Arten zu formalisieren (Kleyer et al. 2000, Schröder 2000). Bei Präsenz-Absenz-Daten kommt für die Habitatmodellierung als mathematisches Verfahren die logistische Regression in Frage (Hosmer & Lemeshow 2000, Jongman et al. 1995). Mit diesem Verfahren werden die Zusammenhänge zwischen dem Vorkommen des Esparsetten-Widderchens und den Habitatfaktoren quantifiziert. Darauf aufbauend werden die Trockenrasen hinsichtlich ihrer Vorkommenswahrscheinlichkeit der Art klassifiziert. Für das Esparsetten-Widderchen wurden in anderen Untersuchungsgebieten ebenfalls mit logistischen Regressionsanalysen Habitatmodelle erstellt (Binzenhöfer et al. 2005). Um die Bedeutung der Konnektivität für das Vorkommen der Art zu analysieren, wurden in der vorliegenden Untersuchung Modelle mit und ohne Berücksichtigung der Konnektivität miteinander verglichen. In Tab. 1 sind die im Gelände erhobenen und mit dem GIS ermittelten Gebietsparameter zusammengestellt.
5 Ergebnisse
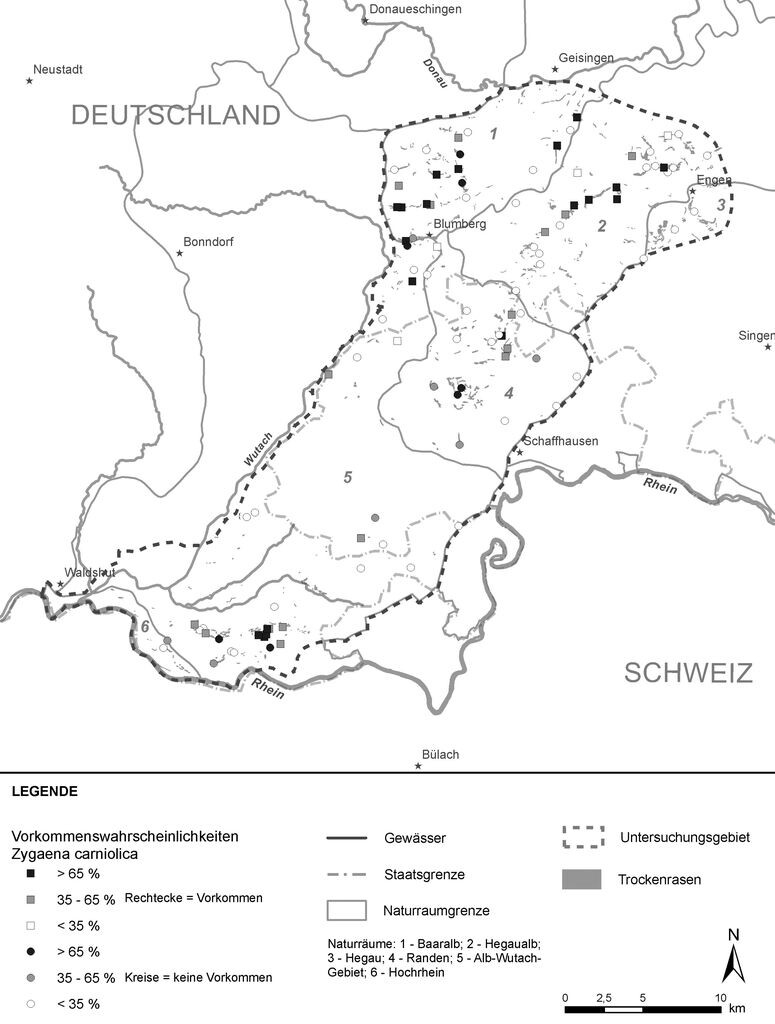
In Abb. 4 sind die Verbundräume für die 750 m-Distanz dargestellt. Diese Distanz liegt in einem Bereich, in dem sich Metapopulationsstrukturen von Widderchen am wahrscheinlichsten abbilden lassen (vgl. Jooss 2006, Wagner 2002). Die Karte zeigt, dass in der Region Hegau Randen Klettgau der Großteil der potenziellen Habitate für das Esparsetten-Widderchen in einer Verbundsituation vorliegt, in welcher ein Teil der Individuen die nächstgelegenen Nachbarflächen erreichen können.
Die Verbundsituation der Trockenrasen im Untersuchungsgebiet hat einen entscheidenden Einfluss auf das Vorkommen des Esparsetten-Widderchens und wurde im Modell nach dem Verfilzungsgrad und dem Kräuteranteil der Vegetation als drittwichtigste Variable ermittelt. In kräuterreichen, wenig verfilzten Beständen ist die Vorkommenswahrscheinlichkeit höher als in ungenutzten, grasreichen Flächen. Der Verfilzungsgrad der Flächen ist von der Nutzung abhängig, da in ungenutzten Flächen krautige Pflanzen gegenüber Gräsern konkurrenzschwächer sind. In ungenutzten, versaumenden Flächen wurde das Esparsetten-Widderchen im Untersuchungsgebiet vereinzelt nachgewiesen. Zu deren Vorkommen in verfilzten, ruderalisierten Flächen bemerkte Hofmann (1994), dass diese Standorte zum Imaginalhabitatspektrum in Baden-Württemberg gehören, aber mit Sicherheit nicht zum eigentlichen Larvalhabitat.
Einen wesentlichen Einfluss auf das Artvorkommen haben die Futterpflanzen, wobei im Untersuchungsgebiet die Raupennahrung eine wichtigere Rolle spielt als die Nahrung der Falter. In dem Vorhersagemodell sind die Raupenfutterpflanzen, gefolgt von den Falterfutterpflanzen und der Trockenheit der Vegetation, die nächst wichtigen Faktoren. Nach Gruppenvergleichstests war das Vorkommen des Esparsetten-Widderchens in größeren Flächen häufiger als in den kleineren Flächen. Die multivariaten Analysen zeigten jedoch, dass nicht die Flächengröße per se, sondern von der Flächengröße abhängige Habitateigenschaften für das Vorkommen entscheidend sind. Die kleinen Flächen werden häufig vom Menschen nicht bewirtschaftet und verfügen daher aufgrund ihres hohen Verfilzungsgrades über eine geringere Habitatqualität.
In Abb. 5 ist die berechnete Vorkommenswahrscheinlichkeit für das Esparsetten-Widderchen mit einem aus sechs Variablen erstelltem Modell sowie die im Gelände kartierten Präsenz-Absenz-Informationen dargestellt. Für das gesamte Untersuchungsgebiet ist mit einem Anteil von über 80 % korrekter Prognosen der Großteil der Vorhersagen richtig.
In den Flächen des südlichen Randens kamen trotz hoher Vorkommenwahrscheinlichkeit keine Esparsetten-Widderchen vor. Die Habitatqualität ist in den mageren Trockenrasen der Hochflächen beispielsweise am Mösli als geeignet einzustufen. Das Fehlen der Art könnte auf die erhöhte Isolation, bedingt durch die Waldgebiete, zurückgeführt werden. Die klimatische Ungunst auf den Hochflächen ist als Erklärung unwahrscheinlich, da in der Baaralb das Esparsetten-Widderchen auch in Höhenlagen über 800 m NN vorkam. Die einfachste Erklärung ist, dass die Art im südlichen Randen einfach nicht verbreitet ist und aufgrund der relativ abgeschiedenen Lage bislang keine Besiedlung stattfand. Die relative Seltenheit des Esparsetten-Widderchens in dem Gebiet um Hemmental ist historisch durch die Aufzeichnungen von Ries, welcher um 1920 die Falter des Randens beobachtete, belegt (Vogelsanger 1936/37).
6 Diskussion: Auswirkungen der Fragmentierung auf die Zielarten
Die Ergebnisse bestätigen, dass die Konnektivität für das Vorkommen des Esparsetten-Widderchens eine entscheidende Rolle spielt. Eine gute Verbundsituation alleine reicht aber für das Vorkommen der Art nicht aus. Die Isolation und die Habitatqualität sind wichtige Faktoren für das Überleben von Tagfaltern, die in Metapopulationsstrukturen vorkommen (vgl. Thomas et al. 2001). Bei den überwindbaren Distanzen scheint die Entfernung der Flächen von geringerer Bedeutung zu sein, solange sie unterhalb von 700 bis 800 m liegen. Der Wert entspricht den in Untersuchungen von Hübner (2002), Kreusel (1999), Smolis & Gerken (1987) und Wagner (2002) ermittelten Flugdistanzen. Das Esparsetten-Widderchen kommt auch in kleinen Flächen vor, wenn die Habitatqualität geeignet ist. Die Auswertungen haben gezeigt, dass für das Vorkommen der Art die Konnektivität der Trockenrasen und die Habitatqualität wichtiger sind als die Flächengröße per se. Daraus folgt, dass sowohl kleinere Flächen als auch nicht besetzte, aber potenzielle Habitate für den Biotopverbund des Esparsetten-Widderchens eine wichtige Rolle spielen. Neben den Maßnahmen in den Trockenrasen, welche die Vorrangflächen darstellen, ist eine flächendeckende Extensivierung anzustreben, um die Isolationswirkung der intensiven Nutzökosysteme zu mindern.
Bei der zweiten, hier nur kurz behandelten Art Gewöhnliche Küchenschelle (Pulsatilla vulgaris) ist die Größe der aktuellen Populationen entscheidend, da aufgrund der biologisch-ökologischen Merkmale eine Wiederbesiedlung neuer Standorte so gut wie ausgeschlossen werden kann. Für die Gewöhnliche Küchenschelle ist die Kontinuität der Standorte und damit der zeitliche Verbund der Habitate entscheidend, welche durch die Florenkonstanz in den Reliktföhrenwäldern sichergestellt werden kann (vgl. Wulf 1997). Beide Arten zusammen decken die wesentlichen Merkmale der Biotopstruktur der Trockenrasen im Untersuchungsgebiet ab.
Dank
Für die Durchsicht des Manuskripts und konstruktive Anregungen danke ich Herrn Prof. Dr. Giselher Kaule (ILPOE).
Literatur
Amler, K., Bahl, A., Henle, K., Kaule, G., Poschlod, P., Settele, J. (Hrsg., 1999): Populationsbiologie in der Naturschutzpraxis. Isolation, Flächenbedarf und Biotopansprüche von Pflanzen und Tieren. Ulmer, Stuttgart, 336 S.
Beinlich, B. (1995): Die historische Entwicklung der Schäferei in Südwestdeutschland. Beih. Veröff. Naturschutz Landschaftspfl. Bad.-Württ. 83.
Berthoud, G., Righetti, A., Lebeau, R.P. (2004): Nationales ökologisches Netzwerk REN. Schlussbericht. Schr.-R. Umwelt 373, BUWAL, Bern, 131 S.
Binzenhöfer, B., Schröder, B., Strauss, B., Biedermann, R., Settele, J. (2005): Habitat models and habitat connectivity analysis for butterflies an burnet moths The example of Zygaena carniolica and Coenonympha arcania. Biol. Conserv. 126, 229-243.
Ebert, G., Rennwald, E. (Hrsg., 1994): Die Schmetterlinge Baden-Württembergs, Bd. 3: Nachtfalter. Ulmer, Stuttgart, 518 S.
Fink, P., Riecken, U., Ullrich, K. (2005): Europäische Dimension des Biotopverbundes in Deutschland. Natur und Landschaft 80 (8), 364-368.
Gradmann, R. (1950): Das Pflanzenleben der Schwäbischen Alb. Bd. 1: Allgemeiner Teil. Verlag des Schwäbischen Albvereins, Tübingen, 401 S.
Hermann, G. (1992): Tagfalter und Widderchen. Methodisches Vorgehen bei Bestandsaufnahmen zu Naturschutz- und Eingriffsplanungen. In: Trautner, J., Hrsg., Arten- und Biotopschutz in der Planung: Methodische Standards zur Erfassung von Tierartengruppen, J. Margraf, Weikersheim, 219-238.
Hofmann, A. (1994): Zygaeninae. In: Ebert, G., Rennwald, E., Hrsg., Die Schmetterlinge Baden-Württembergs, Bd. 3: Nachtfalter, Ulmer, Stuttgart, 196-355.
Hosmer, D.W., Lemeshow, S. (2000): Applied Logistic Regression. Wiley, New York, 375 pp.
Hübner, G. (2002): Widderchen im Trockenbiotopverbund. Naturschutz und Landschaftsplanung 34 (6), 189-190.
Jedicke, E. (1994): Biotopverbund. Grundlagen und Maßnahmen einer neuen Naturschutzstrategie. Ulmer, Stuttgart, 287 S.
Jongman, R.H.G., ter Braak, C.J.F., van Tongeren, O.F.R. (1995): Data analysis in community and landscape ecology. Cambridge Univ. Press, Cambridge, 299 pp.
Jooss, R. (2006): Schutzverantwortung von Gemeinden für Zielarten in Baden-Württemberg. Empirische Analyse und naturschutzfachliche Diskussion einer Methode zur Auswahl von Vorranggebieten für den Artenschutz aus landesweiter Sicht. Diss. Univ. Stuttgart, 262 S.
Kaule, G., Jürging, P. (1977): Biotopkartierung für die Landschaftsplanung dargestellt am Beispiel der Donauebene, Region 10, Ingolstadt. Schr.-R. Naturschutz Landschaftspfl. 8, 7-22.
Keel, A. (1995): Vegetationskundlich-ökologische Untersuchungen und Bewirtschaftungsexperimente in Halbtrockenwiesen (Mesobromion) auf dem Schaffhauser Randen. Veröff. Geobot. Inst. Eidg. Techn. Hochschule, Stiftung Rübel 124, Zürich, 181 S.
Kissling, S. (2010): Trockenrasenverbund in der Region Hegau-Randen-Klettgau. Diss. Univ. Stuttgart, 196 S.
Kleyer, M., Kratz, R., Lutze, G., Schröder, B. (2000): Habitatmodelle für Tierarten: Entwicklung, Methoden und Perspektiven für die Anwendung. Zeitschrift für Ökologie und Naturschutz 8, 177-194.
Kreusel, B. (1999): Dispersionsdynamik von Widderchen zwischen verinselten Kalkmagerrasen in Süddeutschland unter besonderer Berücksichtigung von Naturschutzaspekten. Natur und Landschaft 74 (6), 255-265.
Kummer, G. (1937-1947): Die Flora des Kantons Schaffhausen mit Berücksichtigung der Grenzgebiete. Lieferung 1-7. Kuehn, Schaffhausen, 936 S.
Langenauer, R., Köhler, B., Gigon, A. (2000): Ergebnisse 20-jähriger Bewirtschaftungsversuche in Halbtrockenwiesen bei Merishausen. Mitt. Naturforsch. Ges. Schaffhausen 45, 37-47.
Reck, H. (2004): Das Zielartenkonzept. Ein integrativer Ansatz zur Erhaltung der biologischen Vielfalt? In: Wiggering, H., Müller, F., Hrsg., Umweltziele und Indikatoren, Springer, Berlin/Heidelberg, 311-343.
Schiess-Bühler, C. (1993): Tagfalter im Schaffhauser Randen. Neujahrsblatt der Naturforsch. Ges. Schaffhausen 45, 73 S.
Schröder, B. (2000): Habitatmodelle für ein modernes Naturschutzmanagement. In: Gnauck, A., Hrsg., Theorie und Modellierung von Ökosystemen: Workshop Kölpinsee 2000, Shaker, Aachen, 201-224.
SBN (Schweizerischer Bund für Naturschutz, Lepidopterolgogen-Arbeitsgruppe, Hrsg., 1997): Schmetterlinge und ihre Lebensräume. Arten, Gefährdung, Schutz. Bd. 2. Fotorotar AG, Egg/ZH, 679 S.
Smolis, M., Gerken, B. (1987): Zur Frage der Populationsgröße und der intrapopularen Mobilität von tagfliegenden Schmetterlingen, untersucht am Beispiel der Zydaenidenarten (Lepidoptera: Zygaenidae) eines Halbtrockenrasens. Decheniana 140, 102-117.
Stöcklin, J., Ryf, M., Fischer, M. (2000): Small size of remnants of nutrient-poor calcarous grassland (Mesobromion) in the Swiss Jura puts many plant species at the risk of local extinction. Zeitschrift für Ökologie und Naturschutz 9, 109-118.
Thomas, J.A., Bourn, N.A., Clarke, R.T, Stewart, K.E., Simcox, G., Pearman, S., Curtis, R., Goodger, B.A. (2001): The quality and isolation of habitat patches both determine where butterflies persist in fragmented landscapes. Proceedings of the Royal Society B: Biological Sciences 268, 1791-1796.
Vogelsanger, T. (1936/37): Aus dem entomologischen Tagebuch von Dr. med. F. Ris. Mitt. Naturforsch. Ges. Schaffhausen 13, 159-201.
Wagner, W. (2002): Einnischungsmechanismen bei Rotwidderchen (Lepidoptera: Zygaenidae) auf Kalkmagerrasen der Schwäbischen Alb (Baden-Württemberg). Diss. Univ. Ulm, 146 S.
(2006): Präimaginalökologie mitteleuropäischer Zygaena-Arten schwerpunktmäßig untersucht auf Magerrasen der Schwäbischen Alb. In: Fartmann, T., Hermann G., Larvalökologie von Tagfaltern und Widderchen in Mitteleuropa, Abh. Westf. Mus. Naturkde. 68 (3/4), 171-196.
Wiens, J. (2002): Predicting Species Occurence: Progress, Problems, and Prospects. In: Scott, J.M., Heglund, P.J., Morrison, M.L., eds., Predicting Species Occurance: Issues of Accuracy and Scale, Island Press, Washington, 739-749.
Witschel, M. (1980): Xerothermvegetation und dealpine Vegetationskomplexe in Südbaden. Vegetationskundliche Untersuchungen und die Entwicklung eines Wertungsmodells für den Naturschutz. Landesanstalt für Umweltschutz Baden-Württemberg, Karlsruhe, 212 S.
Wulf, M. (1997): Plant species as indicators of ancient woodland in northwestern Germany. J. Veg. Science 8, 635-642.
Zoller, H. (1954): Die Arten der Bromus erectus-Wiesen des Schweizer Juras, ihre Herkunft und ihre Areale mit besonderer Berücksichtigung der Verbreitung in ursprünglicher Vegetation. Veröff. Geobot. Inst. Rübel in Zürich 28, 283 S.
(1958): Die Vegetation und Flora des Schaffhauser Randen mit Vorschlägen zu ihrem Schutze. Mit. Naturforsch Ges. 26, 181-216.
Anschrift der Verfasserin: Dr. Stephanie Kissling, Institut für Landschaftplanung und Ökologie (ILPOE), Universität Stuttgart, Keplerstraße 11, D-70174 Stuttgart, E-Mail sk@ilpoe.uni-stuttgart.de .
-
Abb. 1: Geographische Lage des Untersuchungsgebietes. -
Abb. 2: Trockenrasen im Untersuchungsgebiet: (a) Reliktföhrenwald Langenhaag, (b) Voll- bis Halbtrockenrasen am Steinbruch bei Zimmerholz, (c) Heuwiesen auf dem Randen Mösli, (d) Voll- bis Halbtrockenrasen nordöstlich von Blumberg. Fotos: Stephanie Kissling -
Abb. 3: Zygaena carniolica. (a) Einzelindividuum Beggingen, Heeresbuck, 08.08.2005; (b) Parkstation Küssaburg, Im Allmend, 06.08.2006. -
Abb. 4: Verbundräume für die 750-m-Distanz im Untersuchungsgebiet. -
Abb. 5: Modellierung und Vorkommen von Zygaena carniolica. -
-



Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.