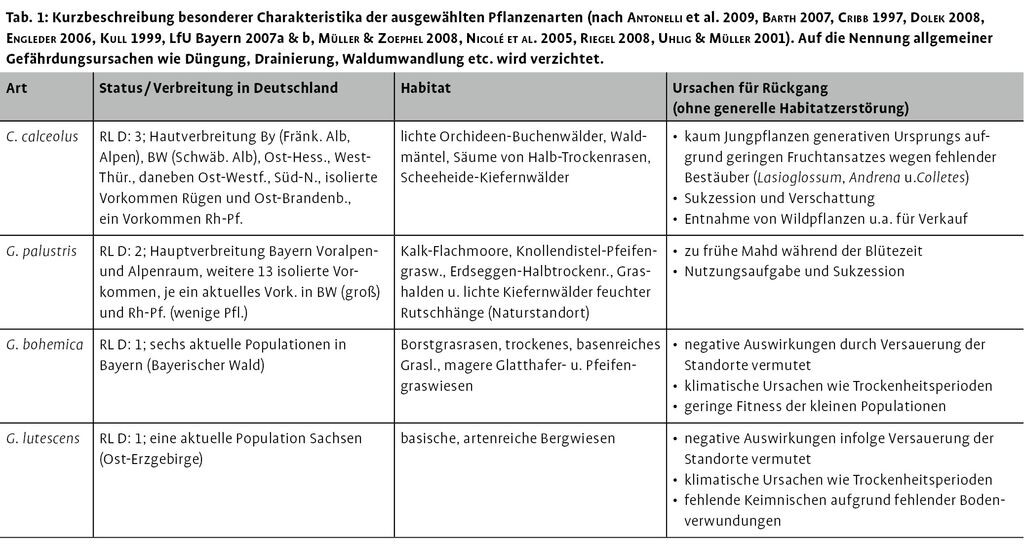
Ex-situ-Kultivierung und In-situManagement als Beitrag zum Artenschutz
Abstracts
In einem Forschungsvorhaben wurde u. a. untersucht, (1) inwieweit sich Ex-situ-Kulturen von ausgewählten Pflanzenarten der FFH-Richtlinie etablieren lassen, (2) welche In-situ-Maßnahmen zur Verbesserung der generativen Reproduktion geeignet sind und (3) unter welchen wirtschaftlichen Rahmenbedingungen In-situ-Management und Ex-situ-Kultivierung im Hinblick auf Eingriffskompensationen möglich ist.
Hierzu wurden bei G. palustris, G. bohemica und G. lutescens zunächst Keimungsexperimente im Klimaschrank unter Laborbedingungen sowie in Topfkulturen im Freiland durchgeführt. Hierbei zeigte sich u.a., dass G. palustris und G. lutescens besonders hohe Keimraten aufwiesen, wenn die Samen vorher einer längeren Kältestratifikation ausgesetzt waren. Wichtig für die Fitness der Jungpflanzen waren eine gute Durchfeuchtung der Substrate während der Sommermonate, eine moderate Düngung, gut durchlüftete Substrate sowie bei Gentianella die Zugabe von mit VA-Mycorrhiza infizierten Substraten. In multiplen Regressionsanalysen erwies sich jedoch die Tiefe der Pflanzgefäße als entscheidende Ressource für die Blütenanzahl der Pflanzen von G. lutescens und G. bohemica.
Bei In-situ-Maßnahmen an G. palustris und C. calceolus zeigte sich, dass bei ersterer in einem Aussaatexperiment signifikant mehr Jungpflanzen in Flächen nachgewiesen wurden, wo Vegetation und Streuschicht entfernt worden waren. Jungpflanzen von C. calceolus konnten ebenfalls in solchen Flächen vorgefunden werden. Cypripedium-Stöcke, um die herum Streuschicht und humoser Oberboden entfernt worden war, zeigten nach vier Jahren signifikant mehr Blüten als unbeeinflusste Stöcke.
Die Kostenkalkulation einer Gärtnerei ergab, dass Maßnahmen an einer Cypripedium-Population sowie die In-Kulturnahme von Pflanzenmaterial mit dem Ziel der Vermehrung für Ausbringungsmaßnahmen nach einer Laufzeit von acht Jahren insgesamt 15 180 kosten würde.
Der Einsatz der aufgezeigten In-situ-Maßnahmen sowie die Etablierung von Ex-situ-Kulturen für Ausbringungsmaßnahmen werden im Hinblick auf zu leistende Kompensationen bei Eingriffen in Natur und Landschaft diskutiert.
Ex-Situ Cultivation and In-Situ Management as Contribution to the Protection of Species The examples of Cypripedium calceolus, Gladiolus palustris, Gentianella bohemica and Gentianella lutescens
The research study investigated the following issues: 1)To what extent can ex-situ cultures be established of species listed in the appendices of the Habitats Directive of the EU? 2)Which simple in-situ measures work to enhance seedling-recruitment? 3)Which are the economic framework conditions of these measures regarding ecological compensation?
First of all, germination experiments for G. palustris, G. lutescens and G. bohemica were carried out in a climate chamber both in a laboratory and in outdoor pot-cultures. G. palustris and G. lutescens exhibited the highest germination rates when subjected to cold-stratification schemes. Fitness of juvenile plants was positively affected by moderate substrate moisture during summer months, moderate fertilization, good aeration of the substrate, and application of VA-Mycorrhiza-substrates. Multiple regression analysis revealed, however, that the number of flowers in Gentianella plants was significantly correlated with the pot depth.
In-situ measures showed that in sowing experiments significantly more seedlings of G. palustris could be recorded in plots where vegetation and litter layer were removed. Similarly, juvenile plants of C. calceolus could only be recorded in plots where the litter and topsoil layer had been removed three years before. If topsoil and litter layer were removed in a patch around Cypripedium plants, these plants showed significantly more flowers after three years than untreated individuals.
The calculation of costs carried out by a nursery showed that in-situ management and ex-situ cultivation of a Cypripedium population with the aim to yield material for re-introduction would cost 15,180 over a period of eight years.
Finally, the study discusses the in-situ measures investigated in this study as well as the establishment of ex-situ cultivation of highly endangered species and species listed in the Appendices of the Habitats Directive in the light of necessary ecological compensation measures.
- Veröffentlicht am

1 Einführung
Aus der FFH-Richtlinie (92/43/EWG) ergibt sich für die Tier- und Pflanzenarten der Anhänge II und IV unmittelbar, dass sich deren Bestandsituation nicht verschlechtern darf und bei nicht vermeidbaren Eingriffen und Beeinträchtigungen ihrer Habitate vorgezogene Ausgleichsmaßnahmen durchzuführen sind (EU-Kommission 2007, Louis 2007). Die Bestandsituation vieler Anhang-Arten verschlechtert sich jedoch weiterhin, auch ohne dass hierzu konkrete Beeinträchtigungen ihrer Habitate infolge von Eingriffen in Natur und Landschaft verantwortlich sind. Beispielsweise konstatiert das landesweite FFH-Artgutachten für den Frauenschuh (Cypripedium calceolus) in Hessen eine andauernde, z.T. dramatische Abnahme der Bestände, sowohl was die Anzahl der Populationen als auch die Anzahl von Pflanzen in den Populationen betrifft (Barth 2007). Dieser Rückgang wird auch für ganz Europa beschrieben (Kull 1999, Nicolé et al. 2005). Ebenso geht z.B. die FFH-Anhang-Art Böhmischer Enzian (Gentianella bohemica), die endemisch für das bayrisch-böhmische Grenzgebiet ist, trotz vielfältiger Bemühungen und Managementmaßnahmen in den Populationen auf bayerischer Seite bisher weiter zurück (Dolek 2008, Engleder 2006, Zipp 2008).
Viele der bisherigen In-situ-Management-Maßnahmen sind zum Teil berechtigterweise recht konservativ. So beschränkt sich das Management von Cypripedium-Populationen häufig auf die Freistellung der Bestände zur Verminderung der Beschattung (Entnahme von Bäumen). In Populationen seltener und gefährdeter Arten bestehen jedoch häufig Probleme hinsichtlich der Rekrutierung von Jungpflanzen, also bei der generativen Vermehrung (Barth 2007, Kull 1998). Dem insgesamt geringen Kenntnisstand zu praktikablen und wirksamen Management-Maßnahmen wird durch ein Forschungs- und Entwicklungs-Vorhaben Managementempfehlungen für Arten des Anhangs IV der FFH-Richtlinie zu begegnen versucht.
Bei Eingriffen in Natur und Landschaft ist eine Beeinträchtigung von Populationen der FFH-Anhangs-Arten wie auch anderer besonders geschützter Arten nicht immer vermeidbar. Hierzu sieht z.B. die FFH-Richtlinie vor, dass Eingriffe, die zur Beeinträchtigung der Habitate und/oder Populationen von Arten der FFH-Anhänge führen (können), durch so genannte CEF-Maßnahmen (measures to ensure Continued Ecological Functionality, im BNatschG: vorgezogene Ausgleichsmaßnahmen) im Rahmen eines Kohärenzausgleichs kompensiert werden können. Demnach ist für die Etablierung eines gleichwertigen Habitates mit einer entsprechenden funktionierenden Population der jeweiligen Art vom Vorhabensträger vor Durchführung des Eingriffs zu sorgen (EU-Kommisson 2007, Lüttmann 2007, Sobotta 2007). In vielen Fällen könnte demnach eine Umsiedlung oder Wiederansiedlung von Populationen von Anhang-Arten als Ausgleich oder Eingriffsminderung in Betracht gezogen werden, doch fehlt für solche Maßnahmen vielfach das nötige populationsökologische und biologisch-technische Wissen hinsichtlich der betroffenen Arten (Thum & Wätzold 2007, Wachter et al. 2004). Auch hinsichtlich geeigneter In-situ-Maßnahmen mangelt es im Hinblick auf Eingriffskompensation bei vielen Arten zum Teil am nötigen Wissen (Lüttmann 2007, Wachter et al. 2004). Ein weiteres großes Problem ist in dem Mangel an geeignetem Material (Individuen aus Nachzuchten, Saatgut etc.) für etwaige Populationsstützungs-Maßnahmen oder Wiederansiedlungen zu sehen. Dieses sollte den für die Region typischen und genetisch möglichst nahe gelegenen, noch vorhandenen Populationen ähneln (Donath & Eckstein 2008).
Um diesen Anforderungen Rechnung zu tragen, wird in jüngerer Zeit immer häufiger die Etablierung von Ex-situ-Kulturen entsprechender Arten diskutiert (Borner 2006, GSPC 2002 in BGCI 2007, Rauer et al. 2000, Welk et al. 2002). Diese Diskussion wird auch dadurch verstärkt, dass sich die Bundesrepublik in internationalen Konventionen (z.B. CBD) und entsprechenden Umsetzungsstrategien (z.B. Global Strategy of Plant Conservation (GSPC) 2002 in BGCI 2007) verpflichtet hat, Pflanzenarten, für die Deutschland eine besondere Verantwortung aufweist (vgl. Ludwig et al. 2007), zu schützen. Diese Arten, die innerhalb Deutschlands ihren Verbreitungsschwerpunkt haben, endemisch oder sub-endemisch sind und aus diesen Gründen zumeist in der bundesdeutschen Roten Liste als Vom Aussterben bedroht gelistet werden, sollen sowohl durch geeignete In-situ-Maßnahmen als auch durch eine ausreichende Etablierung von Ex-situ-Kulturen nachhaltig gesichert werden. Hier können z.B. botanische Gärten eine große Rolle spielen (Arbeitskreis Ex-situ-Kulturen botanische Gärten, mdl. Mitt. M. Burkhard). Die Kultur dieser seltenen und gefährdeten Arten ist jedoch häufig nicht einfach. Auch aus diesem Grund existieren nicht für alle der Arten, die Deutschland im Zuge der internationalen Konventionen und der Umsetzungsempfehlungen der GSPC vorrangig schützen muss, Ex-situ-Kulturen. So war es z.B. bis zu den Arbeiten im Rahmen dieses Vorhabens nicht gelungen, eine Ex-situ-Kultur des Karpaten-Enzians (Gentianella lutescens) aufzubauen, der aktuell nur noch eine Population in Deutschland aufweist (Müller & Zoephel 2008, Uhlig & Müller 2001). Auch sind auf Herkunftsregionen bezogene Ex-situ-Kulturen häufigerer FFH-Anhang-Arten nur lückenhaft vorhanden (mdl. Mitt. M. Burkhard).
Vor diesem Hintergrund hatte ein über die Bundesanstalt für Landwirtschaft und Ernährung (BLE) gefördertes Modell- und Demonstrationsvorhaben (MuD) im Bereich der Erhaltung der biologischen Vielfalt unter anderem folgende Ziele:
Etablierung von Ex-situ-Kulturen von ausgewählten Pflanzenarten der FFH-Richtlinie und der in Deutschland vom Aussterben bedrohten Art Gentianella lutescens (Tab. 1),
Untersuchung der Kulturbedingungen und Gewinnung von Saatgut für potenzielle Ausbringungsmaßnahmen,
Erprobung einfacher In-situ-Maßnahmen zur Verbesserung der generativen Reproduktion vor allem durch Aussaat-Experimente,
Untersuchung wirtschaftlicher Rahmenbedingungen für In-situ-Management und Ex-situ-Kultivierung im Hinblick auf Kompensationsmaßnahmen bei Eingriffen in Natur und Landschaft.
2 Methoden
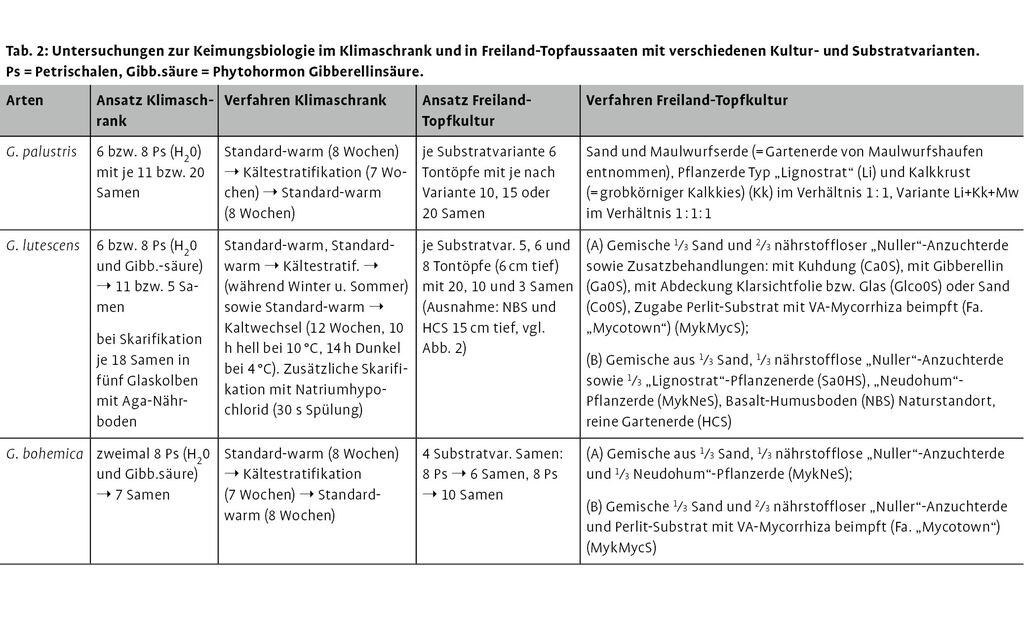
2.1 Ex-situ-Kultivierung: Keimungs- und Haltungsbedingungen von Gladiolus palustris, Gentianella lutescens und G. bohemica
Im Rahmen der Erprobung und Etablierung geeigneter Ex-situ-Kulturbedingungen wurden mit Ausnahme von C. calceolus, für den diese Methoden aus artspezifischen Gründen (besondere Keimungsbiologie) so nicht durchführbar sind zunächst Untersuchungen zur Ermittlung der optimalen Keimungsbedingungen im Labor in einem Klimaschrank durchgeführt. Mittels dieses Klimaschrankes lassen sich Tageslängen simulieren sowie Temperaturgänge (Warm-, Kaltphasen) steuern, die z.B. durch eine Kältestratifikation hartschalige Samen bei Frostkeimern zur Keimung bringen. Hierbei bedeutet z.B. das Verfahren Standard-warm: Tag (14 h bei 20 °C) / Nacht (10 h bei 10 °C)-Wechsel oder das Verfahren Kältestratifikation: Dauerdunkel bei Temperaturen von 0 bis 4 °C. Die Samen wurden auf feuchtem Filterpapier in Petrischalen in statistisch absicherbarer Anzahl (s.u.) in den Klimaschrank verbracht und bei unterschiedlicher Dauer (mehrere Wochen oder Monate) verschiedenen Behandlungen, z.B. Dauerdunkel, Dauerfrost etc., Befeuchtung mit dem keimungsunterstützenden Phytohormon Gibberellinsäure oder nur mit Wasser ausgesetzt (Tab. 2, s. Brunzel & Fischer 2009).
In ähnlicher Weise wurden Topfkulturen angelegt, bei denen die Samen in Tontöpfe gesät wurden, die dann zur Überwinterung bis ins nächste Frühjahr im Freiland verblieben. Auch hier wurde durch verschieden Substratvarianten und Behandlungen (z.B. mit und ohne Düngung) versucht, die optimalen Keimbedingungen (und später Wuchsbedingungen der Jungpflanzen) herauszufinden (vgl. Tab. 2). So wurden z.B. im Rahmen der experimentellen Untersuchungen zu Keimungsbiologie und Kulturbedingungen von G. lutescens 1 112 Samen der Art ausgebracht. An den hieraus gekeimten Jungpflanzen wurden nach Pikieren in unterschiedlich große Tontöpfe und Substrate verschiedene Fitnessparameter wie Keimraten, Mortalitätsraten, Jungpflanzengröße (Durchmesser der Blattrosette = Abstand zwischen Spitzen der zwei größten Blätter) und Anzahl der Blüten/Pflanze sowie die Samenzahl/Blüte gemessen.
2.2 In-situ-Management: Erprobung einfacher Maßnahmen zur Verbesserung der generativen Reproduktion von Cypripedium calceolus und Gladiolus palustris
Für Pflanzenarten vor allem in dynamischen Lebensräumen ist bekannt, das für ihre generative Reproduktion Störungen ihrer Habitate notwendig sind, damit geeignete Nischen für Keimung und Jungpflanzenetablierung entstehen können (Brunzel et al. 2009, Züghart 2002). Diese Zusammenhänge bilden auch den Ausgangspunkt der Überlegungen zur Verbesserung der generativen Reproduktion bei den in dieser Untersuchung behandelten Arten, obwohl diese in wenig dynamischen Lebensräumen vorkommen. Die Hypothese für die häufig geringe generative Reproduktion ist, dass ausgestreute Samen nicht in geeignete Keimnischen gelangen bzw. sich Keimlinge in der vorhandenen Vegetation oder dicht liegenden Streuschicht nicht etablieren können und absterben. So müssen z.B. die Samen von Cypripedium unter die Streuschicht an die Grenze von Humus zu mineralischem Boden gelangen, damit sie von der zur Keimung benötigten Mycorrhiza befallen werden können (Cribb 1997, Kull 1999, Rasmussen 1995).
Um diese Hypothese zu testen, wurden bei C. calceolus und G. palustris in 2007 Aussaatexperimente in einem randomisierten Blockdesign durchgeführt. Hierzu wurden strickleiterartig nebeneinander je acht Paare von 25 cm² großen Aussaatflächen gelegt, von denen die eine Hälfte bis auf den mineralischen Boden (Grenze Humusschicht mineralischer Boden) von Streu und Vegetation befreit wurden. In diese Variante wurde die gleiche Menge Samen gesät wie in die direkt angrenzende Variante, in der Vegetation und Streuschicht ungestört belassen wurden. Im Falle von G. palustris wurden in vier Populationen insgesamt acht Aussaaten vorgenommen und in jede Aussaatfläche zehn Samen gesät. Bei C. calceolus wurden acht Aussaatexperimente in fünf verschiedenen Populationen vorgenommen. Bei dieser Art konnten die mikroskopisch kleinen Samen nicht gezählt werden. Hier wurde ein kleiner medizinischer Laborlöffel jeweils gestrichen voll mit Samen gefüllt, die vor Ort kurz zuvor einer reifen Kapsel entnommen worden waren. Jede 25-cm²-Aussaatfläche wurde auch bei Nicht-Auffinden von Keimlingen maximal 15 min lang abgesucht.
Zusätzlich zu den Aussaatexperimenten im Blockdesign wurden bei C. calceolus weitere Störungsexperimente in vier Populationen durchgeführt. Hierbei wurden um einen Teil der Cypripedium-Stöcke einer Population herum die Streu und Vegetation auf 0,5 bis 1 m² Fläche entfernt. Die anderen Stöcke der Population blieben unbeeinflusst. Auf diese Weise wurden in den vier Populationen zwölf Stöcke gestört, um acht herum blieb die Vegetation und Streuschicht unbeeinflusst. Zum einen sollte durch dieses Experiment ebenfalls getestet werden, ob auf den vegetationslosen Flächen Samen keimen, die in diesem Fall auf natürliche Weise von dem in der Experimentfläche befindlichen fruchtenden Stock ausgestreut wurden. Zum anderen sollte geprüft werden, ob die von der Streuschicht und Konkurrenz befreiten Stöcke im Verlauf der Jahre eine größere Fitness als die Kontrollstöcke zeigen. Die Fitness wurde in Form der Blütenanzahl/Jahr ermittelt.
3 Ergebnisse
3.1 Ex-situ-Kultivierung: Keimungs- und Haltungsbedingungen von Gladiolus palustris, Gentianella lutescens und G. bohemica
Die Keimraten von G. palustris in den Petrischalen lagen bei über 90 % (Keimung nach vier bis sechs Tagen) und zeigten zwischen den Populationen keine signifikanten Unterschiede.
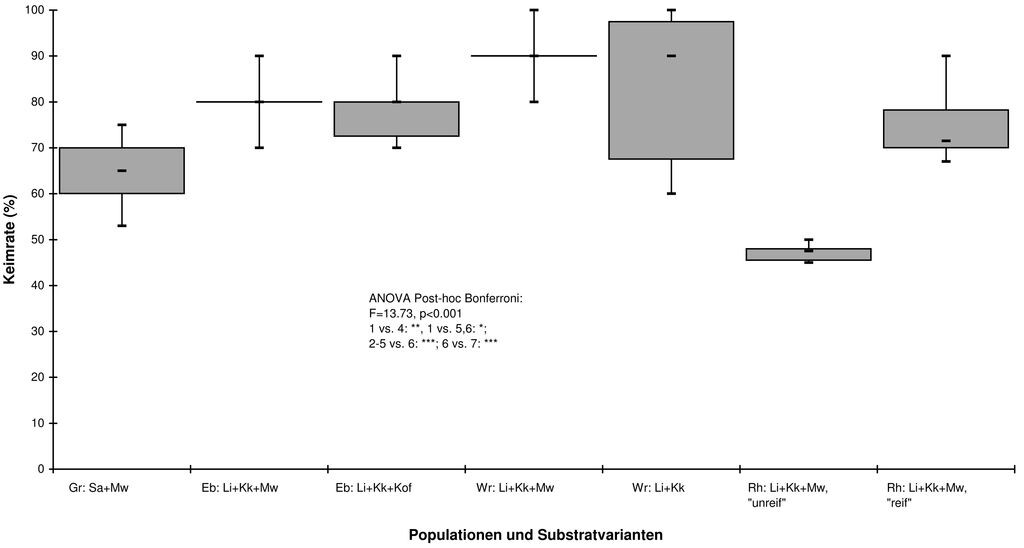
Bei einfacher Freilandüberwinterung zeigen die Ansätze von G. palustris in Topfkultur dagegen insgesamt etwas niedrige Keimraten als im Labor (Abb. 1). Auch gibt es deutliche Unterschiede zwischen dem Substraten. So weist die Variante Sand und Maulwurfserde (= Gartenerde von Maulwurfshaufen entnommen) (Sa+Mw) signifikant niedrigere Keimraten als die meisten anderen auf. Die höchsten Keimraten zeigt die Substratmischung Pflanzerde Typ Lignostrat (Li) und Kalkkrust (= grobkörniger Kalkkies) (Kk) im Verhältnis 1 zu 1, doch sind die Keimraten nicht signifikant höher als bei den anderen Substraten. Die Variante Li+Kk+Mw mit vor der eigentlichen Reife geernteten Samen zeigt die signifikant niedrigsten Keimraten.
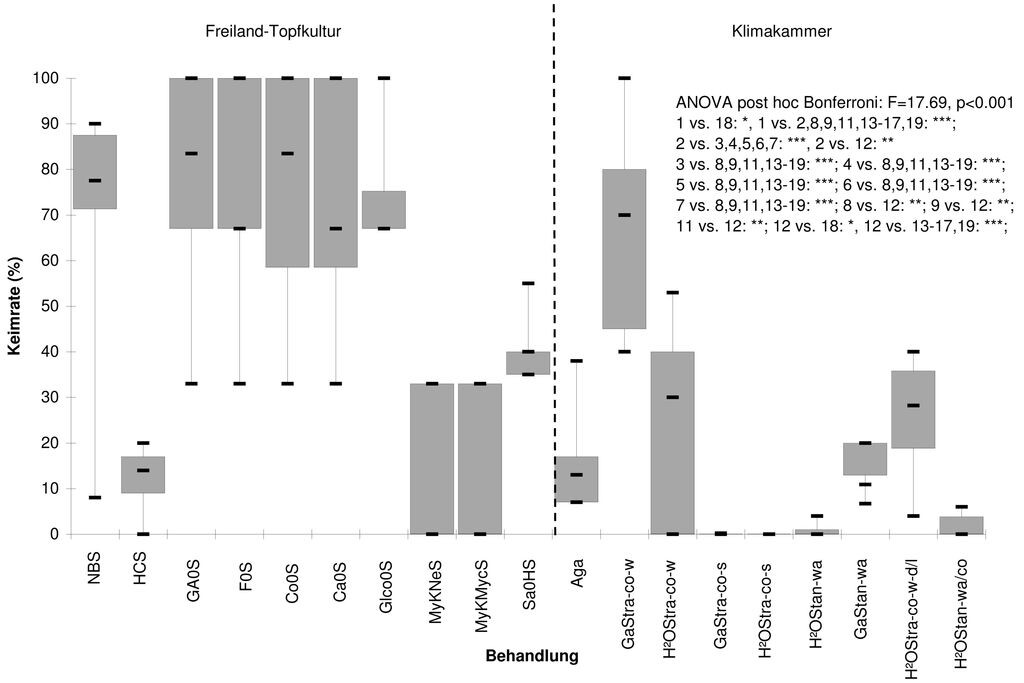
Die Keimung erfolgte bei G. lutescens zwischen dem 25.02. und 10.03. des Folgejahres. Es konnten bei den Freiland-Substratvarianten nur wenige signifikante Unterschiede hinsichtlich der Keimraten festgestellt werden: Die Varianten mit Beimischung von einem Drittel Basalthumus vom Naturstandort, Neudohum-Pflanzenerde oder einer konventionellen, aufgedüngten Gartenerde sowie die Variante mit Beimischung von Mycorrhiza-Substrat wiesen signifikant niedrigere Keimraten als die anderen Varianten auf (Abb. 2). Als ein wesentliches Ergebnis bei den Untersuchungen in der Klimakammer kann zusammenfassend festgehalten werden, dass die Keimraten bei den Verfahren mit Kältestratifikation signifikant am höchsten waren (GaStra-co-w, H2O Stra-co-w, H2O Stra-co-w-d/l) (vgl. Abb. 2). Unter diesen zeigt den Spitzenwert mit bis zu 100 % (Median: 70 %) gekeimten Samen/Petrischale die Variante mit Gibberellinsäure. Die hohen Keimraten bei Kältestratifikation decken sich mit Ergebnissen von Untersuchungen an G. campestris, wo ebenfalls Kältestratifikation die höchsten Keimraten zeitigte (Deno 1996, Milberg 1994).
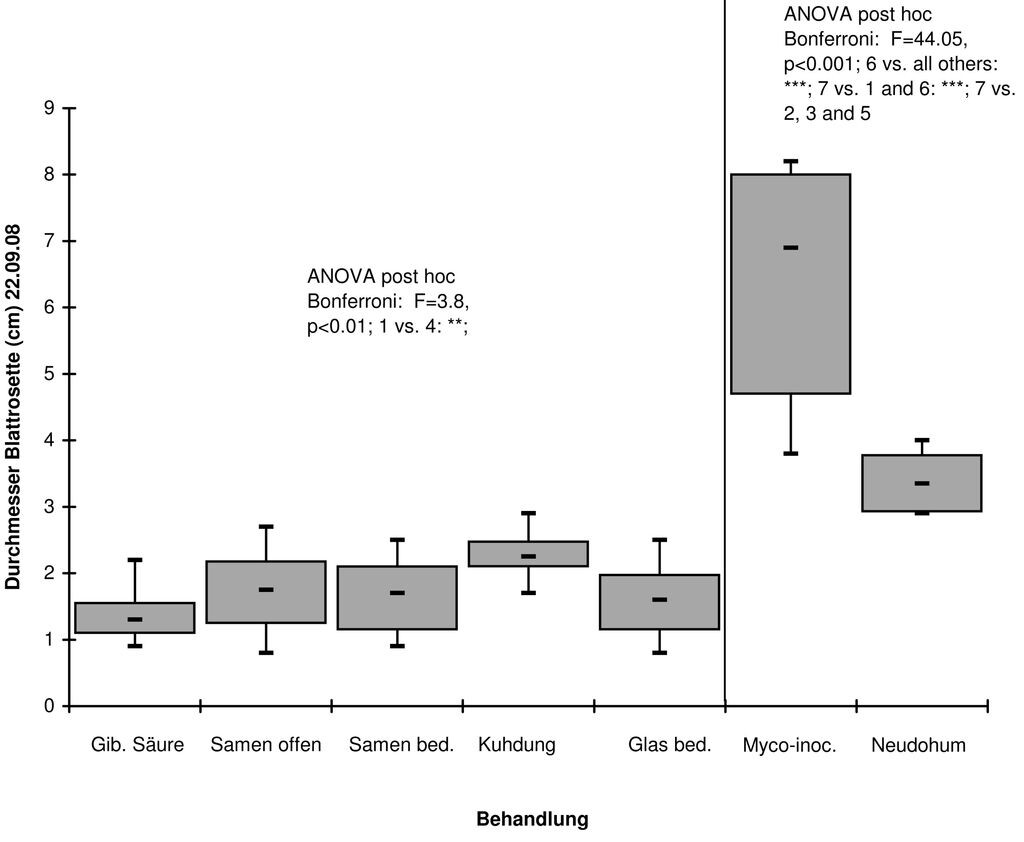
Die Anzahl der Blüten der in der Ex-situ-Kultur herangezogenen Pflanzen schwankte zwischen einer und 32. Je Blüte wurden zwischen 22 und 98 (Mittel: 80) Samenkörner gezählt. Die Auswertungen hinsichtlich der Jungpflanzengröße bei den verschiedenen Substratvarianten in 6 cm tiefen Töpfen zeigen eindeutig, dass die Individuen, deren Substrat eine Zugabe von VA-Mycorrhiza-beimpften Perliten erfahren hatte, signifikant am größten waren (Abb. 3). Am zweitgrößten waren Jungpflanzen, die in eine Mischung 1/3 Sand, 1/3 Lignostrat-Pflanzerde und 1/3 Neudohum-Pflanzerde pikiert wurden. Letztere Pflanzerde gilt ebenfalls als Mycorrhiza-infiziert. Unter den Varianten mit Substraten, die keinerlei Mycorrhiza enthielten, waren die Jungpflanzen, die in mit Kuhdung vermischten Substraten wuchsen, signifikant am größten (vgl. Abb. 3)
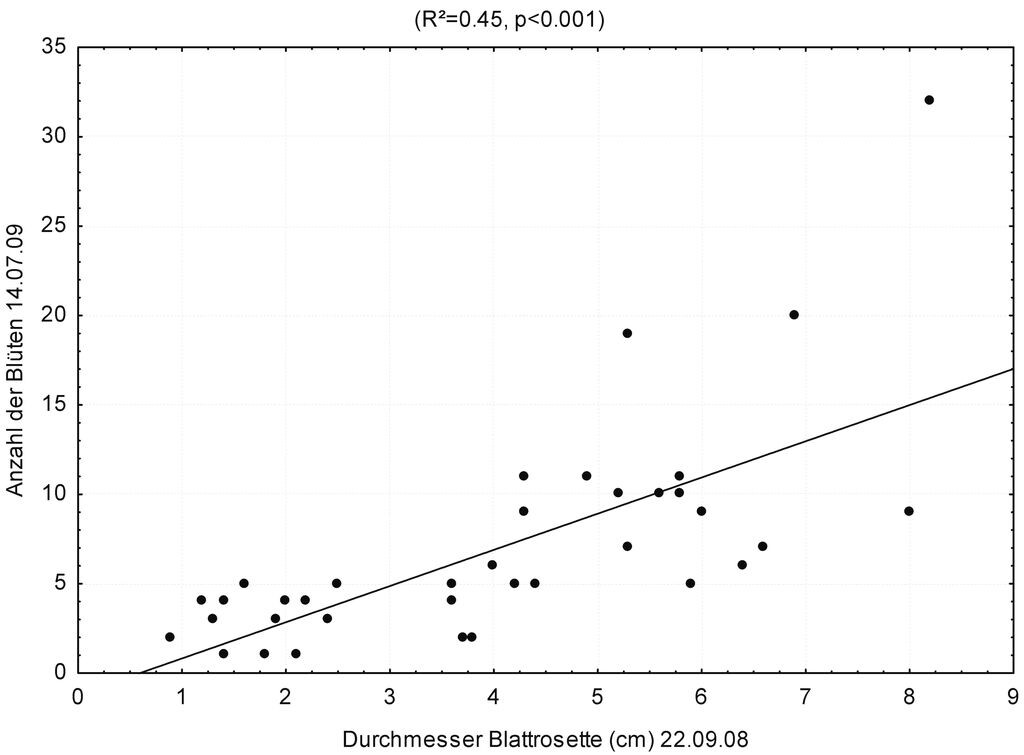
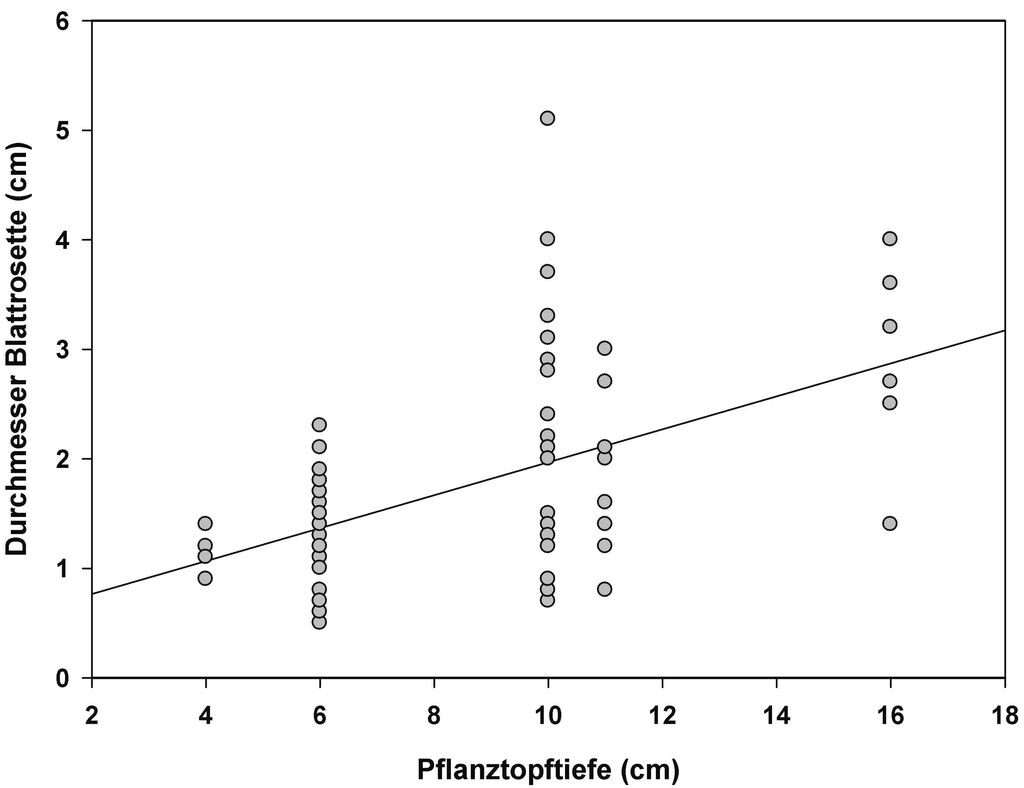
Als alles entscheidend für die gemessenen Fitnessparameter bei G. lutescens erwies sich jedoch die Tiefe der Tontöpfe. So korrelierte die Jungpflanzengröße hochsignifikant positiv mit der Tiefe der Tontöpfe (Abb. 4; R² = 0,11; p < 0,01). Die Jungpflanzengröße des Vorjahres wiederum korreliert bei dieser zweijährigen Art hochsignifikant positiv mit der Anzahl der von den jeweiligen Pflanzen im Folgejahr gebildeten Blüten (Abb. 5).
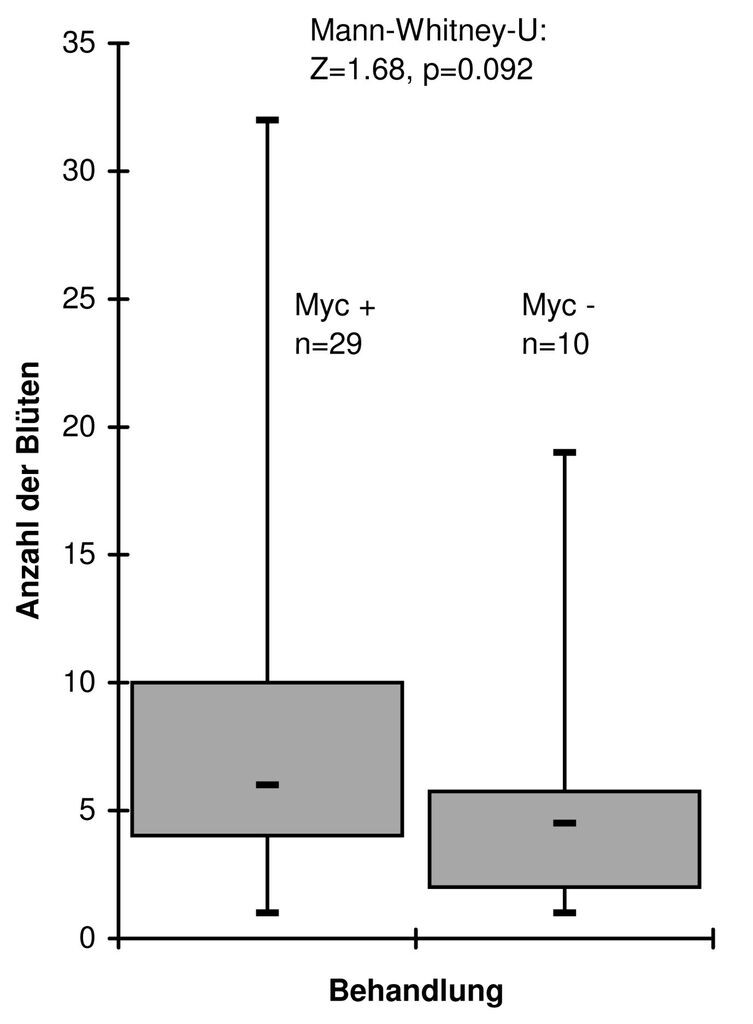
Auch in Mycorrhiza-Substraten sind nicht alle Pflanzen notwendigerweise immer von der VA-Mycorrhiza infiziert, was auch für Gentianella zutrifft (Kempel et al. 2009, mündl. Mitt. Lauerer, Zillig 2009). Um also festzustellen, ob bessere Wuchsleistungen im Falle der Ex-situ-Kulturen von G. lutescens tatsächlich auf Mycorrhiza-Infizierung zurückzuführen sind, wurden die Wurzeln der abgeblühten Individuen auf VAM-Befall untersucht. Hierzu wurden Wurzelproben fixiert, angefärbt und auf Objektträger gebracht. Unter dem Binokular wurden zur statistischen Absicherung des quantitativen Befalls von jeder Probe 60 bis 300 Sichtzählungen der Arbuskeln durchgeführt. Letztere geben am direktesten über die Wirksamkeit des VAM-Befalls Auskunft (vgl. Kempel et al. 2009). Die Ergebnisse dieser Untersuchungen zeigen, dass zwar ein Unterschied hinsichtlich der Blütenzahl/Individuum zwischen der Variante VAM infiziert und nicht VAM-infiziert besteht, dieser aber nicht signifikant ist (Abb. 6). Abb. 6 weist jedoch darauf hin, dass ein signifikanter Unterschied bei größerer n-Zahl der nicht infizierten Individuen nicht ausgeschlossen werden kann. Das hinsichtlich Mycorrhizierung uneindeutige Ergebnis deckt sich jedoch auch mit Beobachtungen an G. bohemica, bei dem die Wurzeln aller im Gelände untersuchten Pflanzen Mycorrhiza-infiziert waren (Dolek 2008), jedoch keine der durchaus üppig blühenden Pflanzen in der Ex-situ-Topfkultur (mdl. Mitt. M. Lauerer). Auch bei G. bohemica zeigt Mykorrhizierung keinen signifikant positiven Einfluss auf die Wuchsleistung (Zillig et al. 2010).
Wenn also die Ressource Topftiefe entscheidend für die Jungpflanzengröße und letztlich für die Blütenanzahl/Pflanze ist, dann muss als wichtiger Ressourcen bestimmender Parameter die Anzahl der Pflanzen je Topf (falls mehre Individuen je Topf gepflanzt waren) bzw. die Anzahl der im Verlauf der Kultur abgestorbenen Pflanzen im jeweiligen Topf in die Analyse miteinbezogen werden. Deshalb wurde eine multiple Regressionsanalyse (MRA, stepforward selection, Ausschlusskriterium 0.05) mit den relevanten determinierenden Variablen Mycorrhiza-Befall (Arbuskeln in %), Topftiefe und der Anzahl der Individuen/Topf sowie den abhängigen Variablen Blütenanzahl und Jungpflanzengröße im Vorjahr durchgeführt. Die Ergebnisse (Tab. 3) zeigen, dass in diesem Modell nur di Topftiefe einen signifikanten positiven Einfluss auf die Blütenanzahl hat. Zieht man in der MRA die Jungpflanzengröße des Vorjahres als erklärende Variable hinzu (die mit der Blütenzahl korreliert, vgl. Abb. 5), so hat nur noch diese einen signifikanten Einfluss auf die Blütenzahl.
Analog zu G. lutescens wurden auch bei G. bohemica Untersuchungen zur Keimungsbiologie und Fitness bei verschieden Behandlungen durchgeführt (vgl. Tab. 2). Diese fielen aufgrund des noch viel geringeren, für die Untersuchungen zur Verfügung stehenden Samenvorrates, weniger umfangreich aus.
Die Pflanzen der Ex-situ-Kulturen besaßen im Mittel 17 Blüten. Im Mittel konnte je Blüte 61 Samen gezählt werden.
Auch für G. bohemica konnte analog zu G. lutescens der signifikante Zusammenhang zwischen Topftiefe und Blütenzahl festgestellt werden (R² = 0,74; p < 0,001). Quantitative, statistisch absicherbare Untersuchungen hinsichtlich des tatsächlichen Mycorrhiza-Befalls konnten aufgrund der geringen Individuenzahl nicht durchgeführt werden (vgl. Zillig et al. 2010).
3.2 In-situ-Management: Erprobung einfacher Maßnahmen zur Verbesserung der generativen Reproduktion von Cypripedium calceolus und Gladiolus palustris
Abb. 7 zeigt, dass diejenigen Cypripedium-Stöcke, bei denen die Streuschicht und Vegetation entfernt wurde (grau, gestrichelte Linie), nach drei Jahren signifikant mehr Blüten aufwiesen als die unbehandelten Stöcke. Dieses Resultat kann damit erklärt werden, dass die Pflanzen in den Flächen ohne darüber liegende Streuschicht schneller und eher im Jahr photosynthetisch aktiv sein können und damit mehr Reserven ansammeln können, die dann für die Blütenbildung zur Verfügung stehen.
Darüber hinaus konnten in zwei der Experimentflächen mit offen liegendem Boden in 2009 eine bzw. vier Jungpflanzen im Einblatt-Stadium festgestellt werden. Leider wurden in den Aussaatflächen des Blockdesigns 2009 noch keine Jungpflanzen gefunden. Dies kann damit zusammenhängen, dass die Blockdesign-Aussaaten erst 2007 eingerichtet wurden und Cypripedium-Protokorme nach Keimung unterirdisch zunächst Nährstoffe sammeln, bevor sie erst nach drei Jahren als Jungpflanze oberirdisch sichtbar werden (Curtis 1943, Kull 1998, Nicolé et al. 2005). Die Kontrollen der Aussaatflächen im Jahr 2010 werden hier weitere Erkenntnisse bringen.
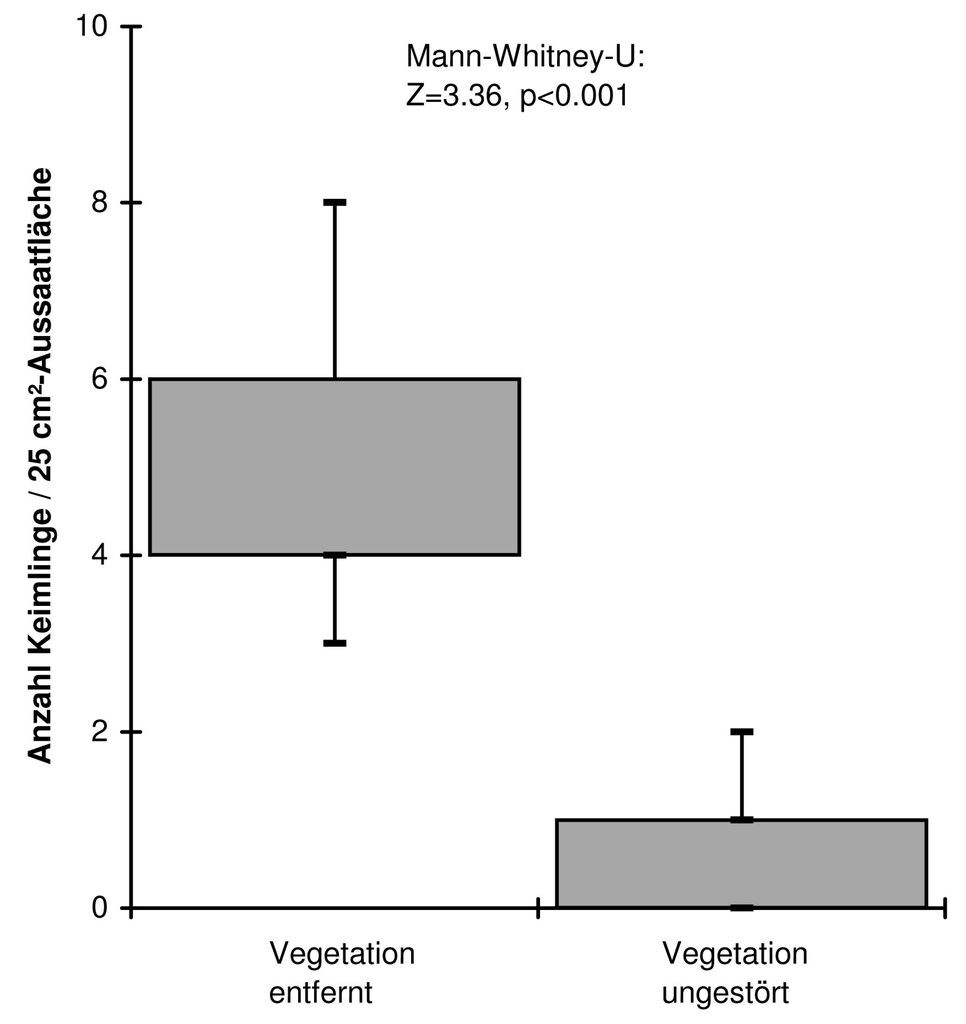
Während der Kontrollen der Aussaatflächen von G. palustris über zwei Jahre hinweg konnten insgesamt 39 Keimlinge in der Aussaatflächenvariante mit entfernter Vegetation, dagegen nur sechsw Keimlinge in der ungestörten Variante festgestellt werden (Abb. 8). Der signifikante Unterschied in diesem Experiment weist auf die Bedeutung hin, die kleinräumige vegetationsfreie Flächen für die Keimung von G. palustris in situ haben können. Es ist jedoch nicht auszuschließen, dass trotz sorgfältiger Suche nach Keimlingen in den 25-cm²-Flächen mit ungestörter Vegetation mehr Keimlinge, die auf den ersten Blick wie Grashalme wirken, übersehen wurden als in den vegetationslosen Flächen. Allerdings ist nicht davon auszugehen, dass es sich nach 15-minütiger Absuche einer 25-cm²-Fläche auch bei vorhandener Vegetation hierbei um größere Anzahlen handelt.
4 In-situ-Maßnahmen und Ex-situ-Kultivierung vor betriebswirtschaftlichem Hintergrund: Aufwand, Kosten, Vermarktung am Beispiel des Frauenschuhs (C. calceolus)
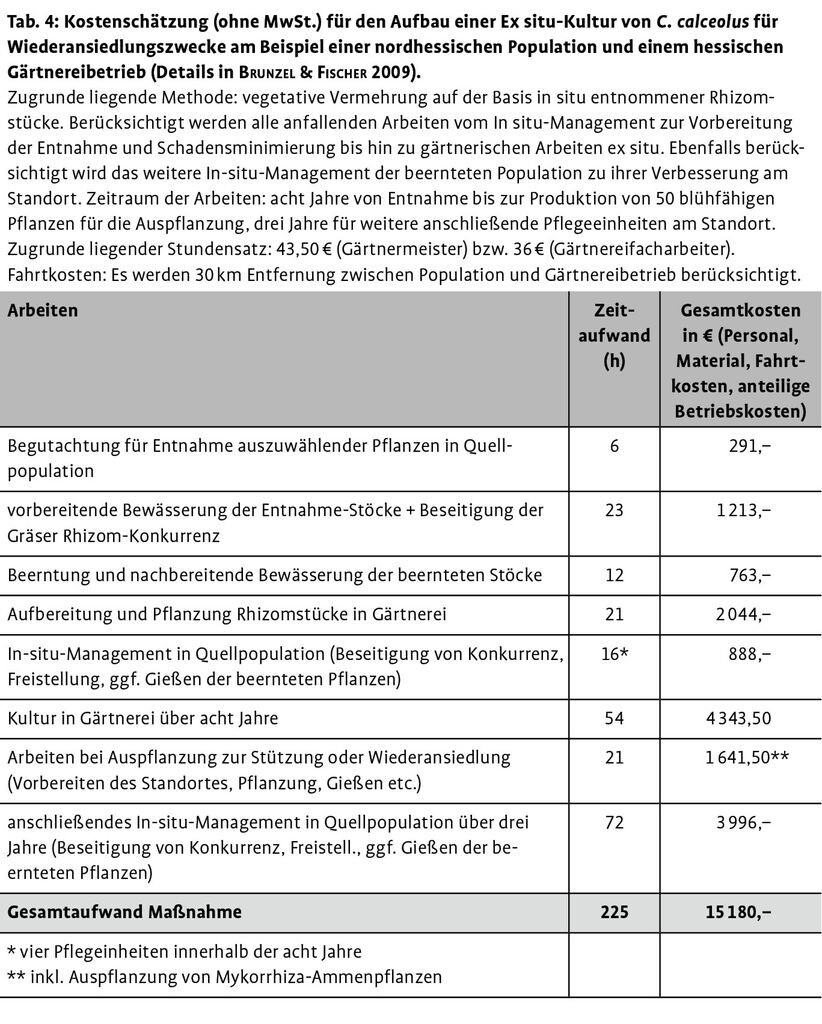
Der Aufbau von Ex-situ-Kulturen und die Durchführung von Wiederansiedlungsprojekten müssen in der Verantwortung von Experten liegen. Zum einen können und sollten mit dieser Aufgabe botanische Gärten betraut werden. Zum anderen erscheint es aber auch möglich, dass z.B. Gärtnereien, die auf die Kultur entsprechenden Arten spezialisiert sind, dies leisten. Gärtnereien, die über das notwendige Fachwissen zur Kultivierung seltener Arten verfügen, bieten diese zumeist aus eigener Nachzucht stammenden Arten auch kommerziell an. So ergab eine durchgeführte Internet-Recherche, dass C. calceolus aus CITES-zertifizierten Nachzuchten zwischen 35 und 45 £ (je nach Größe der blühfähigen Pflanze) und G. palustris zwischen 2,35 und 10,50 CHF (je Zwiebel) kostet. Vor diesem Hintergrund sollte nun geprüft werden, was die In-Kulturnahme einer gefährdeten Cypripedium-Population bei gleichzeitigem In-situ-Management dieser Population für eine spezialisierte Gärtnerei kostet und welche Unkosten nach Abzug potenzieller Einnahmen durch Verkauf von Pflanzen aus den Nachzuchten dieser Population übrig bleiben. Als Beispiel wurde eine nordhessische Gärtnerei und eine Cypripedium-Population in ca. 30 km Entfernung zu dieser Gärtnerei gewählt. Die Kalkulation (Tab. 4, vgl. auch Brunzel & Fischer 2009) wurde von der Gärtnerei selbst durchgeführt und liegt einem tatsächlich begonnenen Projekt zur gärtnerischen Stützung einer hessischen Cypripedium-Population zu Grunde.
Aus dieser Kostenkalkulation ergibt sich, dass bei einer produzierten Gesamtmenge von 50 Pflanzen für eine Wiederansiedlung bzw. Populationsstützung und einer angenommenen, realistischen Projektlaufzeit von acht Jahren Gesamtkosten von 15180 anfallen würden. Dies bedeutet, dass je Pflanze Kosten von 303,60 entstünden. Würden für eine Wiederansiedlung 300 Pflanzen produziert, beliefen sich die Kosten auf ca. 50 bis 60 je Pflanze. Legt man einen für eine Gärtnerei zu erzielenden Marktpreis von mindestens 35 je blühfähiger Pflanze zu Grunde und zieht diese Summe als potenzielle Einnahmequelle (potenziell ca. 200 Pflanzen für Verkauf x 35 ) von den Unkosten ab, so ergibt sich eine Differenz von etwas mehr als 8000 . Hierbei handelt es sich dann um diejenige Summe, mit der ein von einer Gärtnerei durchgeführtes technisches In-situ- und Ex-situ-Management der FFH-Anhang-II-Art C. calceolus von den zuständigen Landesbehörden gegenfinanziert werden müsste. Gleichzeitig ist dies gewissermaßen der Anteil, mit dem der Steuerzahler die last minute-Erhaltung einer durch andere, konservativere Maßnahmen nicht mehr zu haltenden Cypripedium-Population subventionieren müsste.
5 Ausblick
Sowohl für die Arten der FFH-Anhänge als auch für Arten, für die Deutschland aufgrund ihrer Seltenheit eine besondere Verantwortung hat (vgl. Ludwig et al. 2007), ist grundsätzlich anzustreben, diese zunächst in situ und mit geeigneten Maßnahmen zur Verbesserung ihrer Habitate zu erhalten. Da, wo dies jedoch aufgrund der fortgeschrittenen Degradation der Populationen nicht mehr möglich ist, sollte auf das Instrument der Erhaltungskulturen zurückgegriffen werden, die dann Material für Wiederansiedlungen oder Populationstützungen vorhalten können. Gleichzeitig ist infolge schon bestehender gesetzlicher Verpflichtungen wie FFH-Richtlinie und Bundesartenschutzverordnung bei Eingriffen in Natur und Landschaft Kompensation für die Beeinträchtigungen der Populationen entsprechender Arten zu leisten (Lüttmann 2007, Sobotta 2007, Wachter et al. 2004). Beeinträchtigungen werden sich häufig nicht vermeiden lassen und Kompensation ist nicht immer in Form bisher üblicher Praktiken angemessen zu leisten. Ist z.B. das Vorkommen einer FFH-Anhang-Art alternativlos von einer Überbauung bedroht, könnte eine technische Lösung wie die Entnahme und Wiederansiedlung in einem Ersatz-Lebensraum ein Instrument der Kompensation sein. Selbstverständlich sind solche technischen Lösungen als letztes Mittel zu sehen. Sollten sie jedoch zum Einsatz kommen, bestünde dann das Problem, dass das notwendige Wissen oder die nötigen Erhaltungskulturen für derartige Maßnahmen nicht vorhanden sind.
Im Hinblick auf In-situ-Management zur Stützung von Populationen sollten vermehrt Maßnahmen erprobt werden, die auf der Basis der Störung von zu einheitlichen und starren Habitatverhältnisse Nischen für Samenkeimung und Jungpflanzen-Etablierung schaffen. In-situ-Maßnahmen, die die Samenkeimung und Jungpflanzen-Etablierung von C. calceolus, G. palustris und Gentianella-Arten unterstützen, können z.B. kleinflächige Bodenverwundungen, die Entnahme von Oberboden oder die Entfernung der Streuschicht sein (vgl. auch Brunzel & Fischer 2009, Dolek 2008). Maßnahmen, die zur Etablierung von Jungpflanzen führen, tragen dann maßgeblich zur Erhöhung der Überlebenswahrscheinlichkeit insbesondere kleiner Cypripedium-Populationen bei, die ansonsten nur von der Langlebigkeit der Alt-Pflanzen bestimmt wird (Nicolé et al. 2005).
Ob Maßnahmen wie Wieder- oder Neuansiedlungen sowie Populationstützungen auf der Basis von Ex-situ-Kulturen im Rahmen von reinen Artenschutzprojekten oder von Kompensationsleistungen zum Einsatz kommen, ist von Experten und den entsprechenden Behörden zu prüfen und von diesen auch wissenschaftlich zu begleiten. Durchführende Einrichtungen solcher Maßnahmen können botanische Gärten oder entsprechend spezialisierte und zertifizierte Gärtnereien sein. Deren Aufwand für In-situ-Management von Populationen in Kombination mit der Etablierung von Ex-situ-Kulturen sollte entsprechend honoriert werden, wobei nach Beachtung bestimmter Rahmenbedingungen nichts dagegen spricht, die entstehenden Kosten gegen potenzielle Einahmen aus dem Verkauf entsprechend zertifizierter Pflanzen zu rechnen. Dies könnte dann Teil eines generellen Systems sein, bei dem das erfolgreiche Management von Populationen gefährdeter und von Gesetzes wegen zu erhaltender Arten honoriert wird.
Dank
Das Vorhaben Erhaltung und Vermarktung vergessener Zier- und Arzneipflanzen sowie stark gefährdeter Anhang-II-Pflanzenarten der FFH-Richtlinie der Europäischen Union wurde als Modell- und Demonstrationsvorhaben im Bereich der biologischen Vielfalt (Fkz.: 05BM005) durch die Bundesanstalt für Landwirtschaft und Ernährung (BLE), Projektträger Agrarforschung, gefördert.
Literatur
Antonelli, A., Dahlberg, C. J., Calgren, K. H. I., Appelquist, T. (2009): Pollination of the Ladys slipper orchid (Cypripedium calceolus) in Scandinavia taxonomic and conservational aspects. Nordic J. of Botany 27, 266-273.
Barth, U. (2007): Artenhilfskonzept für Cypripedium calceolus (Frauenschuh) in Hessen, Art des Anhangs II der FFH-Richtlinie. Überarbeitete Fassung, Stand März 2008. Im Auftrag von Hessen-Forst, Forsteinrichtung und Naturschutz (FENA), Gießen. Hrsg.: Hessisches Ministerium für Umwelt, ländlichen Raum und Verbraucherschutz.
Bayrisches Landesamt für Umwelt (2006a): Merkblatt Artenschutz 6 Böhmischer Enzian (Gentianella bohemica Skal.). Autoren: M. Dolek, T. Zipp, A. Zehm.
Bayrisches Landesamt für Umwelt (2007 b): Merkblatt Artenschutz 7 Sumpf-Gladiole (Gladiolus palustris GAUDIN). Autor: G. Riegel.
Borner, A. (2006): Preservation of plant genetic resources in the biotechnology era. J. of Biotechnology 12 (1), 1393-1404
Brunzel, S., Fischer, S.F. (2009): Endbericht zum Modell- und Demonstrationsvorhaben Erhaltung und Vermarktung vergessener Zier- und Arzneipflanzen sowie stark gefährdeter Anhang-II-Pflanzenarten der FFH-Richtlinie der Europäischen Union. Marburg.
, Fischer, S.F., Schneider, J., Jetzkowitz, J., Brandl, R. (2009): Neo- and archaeophytes respond more strongly than natives to socio-economic mobility- and disturbance patterns along an urban-rural gradient. J. of Biogeography 36, 835-844.
Cribb, P. (1997): The Genus Cypripedium. Kew Publishing. Royal Botanic Gardens, Kew, 2. Aufl.
Curtis, J.T. (1943): Germination and seedling development in five species of Cypripedium L. American J. of Botany 30, 199-205.
Deno, N.C. (1996): The first Supplement to the second edition of seed germination theory and practice. 150 S. Self-published and self-distributed.
Dolek, M. (2008): Unveröffentlichter Abschlussbericht Artenhilfsprogramm Böhmischer Enzian (Gentianella bohemica) Ergebnisse 2006 bis 2008. 35 S.
Donath, T.W., Eckstein, R.L. (2008): Bedeutung genetischer Faktoren für die Wiederansiedlung seltener Pflanzengemeinschaften. Naturschutz und Landschaftsplanung 40 (1), 21-30.
Engleder, T. (2006): Der Böhmische Kranzenzian / Gentianella bohemica (Gentianaceae) im österreichischen Teil der Böhmischen Masse (Böhmerwald, Mühl- und Waldviertel). Neilreichia 4, 215-220.
EU-Kommission (2007): Guidance document on the strict protection of animal species of community interest provided by the Habitats Directive 92/43/EEC, Final version, February 2007.
Globale Strategie zur Erhaltung der Pflanzen (2007): Deutschsprachige Übersetzung der engl. Originalfassung von 2002, anzufordern bei: Botanical Gardens Conservation International BGCI Deutschland (Botanic Gardens Conservation International (BGCI) 2007: Globale Strategie zur Erhaltung der Pflanzen.)
Louis, H. W. (2007): Perspektiven des Natur- und Artenschutzes. Naturschutz und Landschaftsplanung 39 (8), 228-235.
Ludwig, G., May, R., Otto, C. (2007): Verantwortlichkeit Deutschlands für die weltweite Erhaltung der Farn- und Blütenpflanzen vorläufige Liste. BfN-Skripten 220.
Lüttmann, J. (2007): Artenschutz und Straßenplanung. Naturschutz und Landschaftsplanung 39 (8), 236-241.
Kempel, A., Schmidt, A.K., Brandl, R., Schädler, M. (2009): Support from the underground: Induced plant resistance depends on arbuscular mycorrhizal fungi. Functional Ecology, doi:10.1111/j.1365-2435.
Kull, T. (1998): Fruit-set and recruitment in populations of Cypripedium calceolus L. in Estonia. Botanical J. of the Linnean Society 126, 2738.
(1999): Cypripedium calceolus L. J. Ecol. 87, 913-924.
Milberg, P. (1994): Germination ecology of the endangered grassland biennial Gentianella campestris. Biol. Conserv. 70, 287-290.
Müller, F., Zoephel, B. (2008): Abschlußbericht Erarbeitung eines Konzeptes zur Sicherung und Entwicklung der Population des Karpatenenzians am Geisingberg bei Altenberg Osterzgebirge, April Juni 2008. Technische Universität Dresden, Institut für Botanik, Lehrstuhl für Botanik. Auftraggeber: Regierungspräsidium Dresden, Umweltfachbereich, Dienststelle Radebeul.
Nicolé, F., Brzsosko, E., Till-Bottraud, I. (2005): Population viability analysis of Cypripedium calceolus in a protected area: longevity, stability and persistence. J. Ecol. 93, 716-726.
Rasmussen, H.N. (1995): Terrestrial Orchids from seed to mycotrophic plant. Cambridge University Press, Cambridge, 444 S.
Rauer, G., von den Driesch, M., Ibisch, P.L., Lobin, W., Barthlott, W. (2000): Beitrag der deutschen Botanischen Gärten zur Erhaltung der Biologischen Vielfalt und Genetischer Ressourcen Bestandsaufnahme und Entwicklungskonzept. Bundesamt für Naturschutz, 246 S.
Riegel, G. (2008): Recherchen zur Bestandsituation der Sumpf-Gladiole (Gladiolus palustris) in Bayern. Unveröff. Gutachten im Auftrag des Bayerischen Landesamtes für Umwelt, Augsburg, 10 S.
Sobotta, C. (2007): Artenschutz in der Rechtssprechung des Europäischen Gerichtshofes. Natur und Recht 29, 642-649.
Thum, R., Wätzold, F. (2007): Artenschutz durch handelbare Zertifikate? Grundgedanke des Konzepts und potentielle Einsatzmöglichkeiten im deutschen Rechtssystem. Natur und Recht 29, 299-307.
Uhlig, D., Müller, F. (2001): Zur Bestandsituation ausgewählter vom Aussterben bedrohter und stark gefährdeter Pflanzenarten im Osterzgebirge. Förderverein für die Natur des Osterzgebirges e.V. 1, 1-35.
Wachter, T., Lüttmann, J., Müller-Pfannenstiel, K. (2004): Berücksichtigung von geschützten Arten bei Eingriffen in Natur und Landschaft. Naturschutz und Landschaftsplanung 36 (12), 371-377.
Welk, E. (2002): Arealkundliche Analyse und Bewertung der Schutzrelevanz seltener und gefährdeter Gefäßpflanzen Deutschlands. Schr.-R. Vegetationskde. 37.
Zillig, P. (2009): Vitalität und Wachstum von Gentianella bohemica in Topfkulturen in Abhängigkeit von der Mykorrhizierung. Unveröff. Dipl.-Arb., Univ. Bayreuth.
, Lebermann, K., Morba, D., Arneth, G., Lauerer, M. (2010): Ex-situ-Erhalt des Böhmischen Fransenenzians Erfahrungen bei der Erhaltungskultur im Botanischen Garten Bayreuth. Naturschutz und Landschaftsplanung 42 (3), 78-84.
Zipp, T. (2008): Maßnahmen zur Verbesserung der Bestandssituation von Gentianella bohemica im Bayerischen Wald 2008. Unveröff. Gutachten im Auftrag des Bayerischen Landesamtes für Umwelt (LfU), Augsburg, 21 S.
Züghart, W. (2002) Die Spontanflora nordwestdeutscher Dörfer. Eine floristisch-populationsökologische Untersuchung der dörflichen Spontanflora im Bremer Raum. Diss. Bot. 362, Gebr. Bornträger, Stuttgart.
Anschrift des Verfassers: Dr. Stefan Brunzel, Philipps-Universität Marburg, Fachbereich Biologie, Fachgebiet Spezielle Zoologie und Evolution der Tiere, Karl v. Frisch-Straße 8, D-35032 Marburg, E-Mail brunzel@staff.uni-marburg.de
-
-
-
Abb. 1: Freilandüberwinterungs-Topfkultur von G. palustris: Sa = Sand, MW = Maulwurfshaufen-Erde, Kk = Kalkkrust (grobkörniger Kalkkies), Li = Lignostrat-Pflanzenerde; alles im Mischungsverhältnis 1 : 1 : 1; unreif = zwei bis drei Wochen vor optimaler Ausreifung geerntet. -
Abb. 3: Unterschiede in der Jungpflanzengröße zwischen verschiedenen Substratvarianten bei gleicher Topftiefe (6 cm). Die Varianten 1 bis 6 (von links nach rechts) bestehen jeweils zu 2⁄3 aus nährstoffloser Anzuchterde, 1⁄3 Sand und einer Zusatzbehandlung: Gib.Säure = Gibberellin-Säure, Samen frei = ohne Abdeckung auf Oberfläche: Lichtkeimer, Samen bed. = mit Sand zugedeckt: Dunkelkeimer, Kuhdung = Beimischung von Kuhdung, Glas bed. = mit Glas bzw. Klarsichtfolie abgedeckt, Myco-inoc. = Substrat mit Mycorrhiza-Perliten beimpft. Die Variante „Neudohum“ bestand zu 1⁄3 aus nährstoffloser Anzuchterde, 1⁄3 Sand und 1⁄3 einer „Neudohum“-Pflanzerde. -
Abb. 2: Keimraten in Freiland-Topfkultur und in der Klimakammer.Abkürzungen Substratvarianten Freiland-Topfkultur (von links nach rechts): natürlicher Basaltboden; reiner, nährstoffreicher Gartenboden; in den Varianten 3 bis 7 bedeutet 0 und S eine Mischung aus nährstoffloser Anzuchterde und Sand (Mischungsverhältnisse i.d.R. 1 : 1 : 1). GA = Gibberellin-Säure, F = ohne Abdeckung, Co = Samen mit Sand zugedeckt, Ca = mit Kuhdung gemischt, Glco = mit Glasplatte abgedeckt. Neudohum-Pflanzerde, wahrscheinlich mit VA-Mycorrhiza infiziert, und Sand; Lignostrat-Pflanzerde, mit VA-Mycorrhiza der Firma Mycotown beimpft, und Sand; Gemisch aus Sand, Nuller-Anzuchterde und leicht gedüngter Pflanzenerde. Klimakammerbehandlungen (von links nach rechts): Gibberellin und Stratifikation kalt im Winter, H2O und Stratifikation kalt im Winter, Gibberellin und Stratifikation kalt im Sommer, H2O und Stratifikation kalt im Sommer, H2O Standard warm, Gibberellin Standard warm, H2O und Stratifikation kalt im Winter + hell-dunkel-Wechsel, H2O und Stratifikation kalt-warm-Wechsel. -
Abb. 5: Regression der Jungpflanzengröße im Vorjahr (gemessen von Spitze zu Spitze der längsten Laubblätter) gegen Anzahl der Blüten. Es besteht ein signifikanter positiver Zusammenhang. -
Abb. 4: Signifikanter Zusammenhang zwischen Topftiefe und der Jungpflanzengröße bei G. lutescens (R² = 0,11; p < 0,01). Waren in einem Topf mehrere Individuen gepflanzt, ist die Anzahl der Pflanzen je Topf als Co-Variable in die Analyse eingeflossen (vgl. Tab. 3). -
Abb. 6: Unterschiede in der Anzahl der Blüten (Juli 2009) zwischen Individuen mit tatsächlich mycorrhizierten Wurzeln (Myc +, Grundlage: Zählung der arbusculären Vesikel) und Individuen mit Wurzeln ohne Befall von VA-Mycorrhiza (Myc ). -
-
Abb. 8: Ergebnisse der In situ-Aussaaten im Blockdesign bei G. palustris. Es wurden vier Populationen mit je zwei Sets à acht Plots (25-cm²-Quadrat) je einmal Vegetationsdecke entfernt und ungestört angelegt. In jedem 25-cm²-Quadrat wurden je zehn Samen ausgebracht. In den Plots mit entfernter Vegetationsdecke konnten über zwei Jahre hinweg signifikant mehr Keimlinge festgestellt werden. -
Abb. 7: In-situ-Management von Cypripedium-Stöcken: Blütenzahl bei Stöcken mit (grau, gestrichelte Linie,#NB_SYMBOL[DOT]#= 12 Stöcke) und ohne (weiß, durchgezogene Linie,#NB_SYMBOL[DOT]#= 8 Stöcke) Entnahme der Streuschicht auf einer Fläche von 0,5 bis 1 m² um den jeweiligen Stock. Drei Jahre nach Behandlung weisen die Stöcke mit Entnahme der Streuschicht signifikant mehr Blüten auf. -

![Abb. 7: In-situ-Management von Cypripedium-Stöcken: Blütenzahl bei Stöcken mit (grau, gestrichelte Linie,#NB_SYMBOL[DOT]#= 12 Stöcke) und ohne (weiß, durchgezogene Linie,#NB_SYMBOL[DOT]#= 8 Stöcke) Entnahme der Streuschicht auf einer Fläche von 0,5 bis 1 m² um den jeweiligen Stock. Drei Jahre nach Behandlung weisen die Stöcke mit Entnahme der Streuschicht signifikant mehr Blüten auf.](/vorlagen/webapp/cache/cms/nul-brunzel-abb-71_ge2tgmjwga4a-1024x679.jpg)
Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.