Heuschrecken und Klimawandel
Abstracts
Während in Rheinland-Pfalz in den letzten 100 Jahren die mittlere Jahrestemperatur um 0,8°C angestiegen ist, wird gleichzeitig eine Zunahme der Anzahl der Fang- und Heuschreckenarten festgestellt. Allein seit 1987 erhöhte sich die Anzahl bodenständiger, hauptsächlich aus dem Süden kommender Arten um sieben auf nunmehr 59.
Im selben Zeitraum wird auch eine stetige Ausbreitung bereits seit längerem etablierter Arten festgestellt. Von einem Teil dieser in Ausbreitung begriffenen Arten verläuft die nördliche Arealgrenze durch Rheinland-Pfalz oder nicht allzu weit nördlich davon entfernt (südliche Arten). Doch breiten sich auch einige Arten aus, die bereits seit langem im nördlichen Mittel- und Osteuropa vorkommen (nördliche Arten). Da südliche Arten gleichzeitig überwiegend thermophiler als nördliche sind, profitieren sie offenbar stärker von einer Klimaerwärmung.
Von den meisten seit dem Jahr 1987 hinzugekommenen Neubürgern ist bewiesen oder wird vermutet, dass sie von Süden kommend eingeschleppt wurden. Aber meist erst die verbesserten klimatischen Bedingungen ermöglichten es ihnen, sich zu etablieren.
Grasshoppers, Mantids and Climate Change The propagation of southern species in Rhineland-Palatinate
Within the last 100 years the mean annual temperature in Rhineland-Palatinate has risen by 0.8°C. Simultaneously, the number of grasshoppers and mantids which reproduce locally has increased to 59 species, seven of which were first observed after 1986. In addition, the range of several long established species has increased. The northern limit to the range in Europe of some of these expanding species was (or still is) roughly the northern border of Rhineland-Palatinate; these species are called southern species, in contrast to northern species which are also distributed farther northwards in central and eastern Europe. Among the species with expanding ranges, the proportion of southern species is higher than that of northern species. Being more thermophilic than northern species, southern species are apparently the first to profit from climate warming. For most of the species observed for the first time after 1986 it is proven or suspected that they were introduced from the south through human activity, some would never have reached Rhineland-Palatinate autochorously. However, most were probably able to establish themselves there only because of improved climatic conditions.
- Veröffentlicht am

1 Einleitung
Die bislang festgestellten Auswirkungen des rezenten Klimawandels auf Insekten betreffen die Morphologie, die Phänologie, den Voltinismus, die Abundanzen und das Areal von Insekten (Robinet & Roques 2010). Einige Arten gehören zu den so genannten Klimawandelverlierern; bei ihnen ist eine Verkleinerung ihres Areals oder eine Abnahme der Populationsgröße zu beobachten (Wilson & MacLean 2011). Da die überwiegende Anzahl der Insektenarten thermophil ist, zählen sie aber häufig zu den Klimawandelgewinnern, die entweder ihr Areal nach Norden (auf der Nordhemisphäre) ausdehnen, im Gebirge in die Höhe steigen oder deren Abundanzen zunehmen (Ott 2010, Poniatowski & Fartmann 2011, Wissmann et al. 2009).
Viele Arbeiten über die räumlichen Veränderungen der Insektenfauna sind prognostisch, d.h. sie versuchen vorherzusagen, wie die Verbreitung einer Art zu einem bestimmten Zeitpunkt in der Zukunft aussehen wird (beispielsweise Buse & Griebeler 2011) oder welche Arten durch eine großklimatische Veränderung gefährdet werden könnten (Detzel et al. 2005). Diese Prognosen sind jedoch auf eine verlässliche Datengrundlage angewiesen und bedürfen der Validierung anhand der tatsächlich in der Natur beobachteten Veränderungen; dabei lässt sich die Ausbreitung von Arten leichter feststellen als deren Rückgang (Rabitsch et al. 2010).
In Rheinland-Pfalz wurde in den letzten 100 Jahren ein Anstieg der Jahresmitteltemperatur um durchschnittlich 0,8°C festgestellt (Ministerium für Umwelt, Forsten und Verbraucherschutz Rheinland-Pfalz 2007), was einer Verschiebung der Isothermen um etwa 120km nordwärts entspricht (Leuschner & Schipka 2004). Diese Erwärmung wird mit Veränderungen in der Insektenfauna in Verbindung gebracht. In Rheinland-Pfalz wird eine Ausbreitung südlicher, thermophiler Libellenarten festgestellt (Ott 1996). Bei den Heuschrecken war in Rheinland-Pfalz etwa zwischen 1970 und 1990 eine nordwärts gerichtete Expansion bei der Gemeinen Sichelschrecke (Phaneroptera falcata) feststellbar und ab etwa 1985 bis 1995 auch beim Weinhähnchen (Oecanthus pellucens) (Kleukers 1997, Pfeifer et al. 2011).
Nachfolgend wird überprüft, ob seit 1995 eine Ausbreitung weiterer Fang- und Heuschreckenarten in Rheinland-Pfalz feststellbar ist.
2 Material und Methode
2.1 Datengrundlage
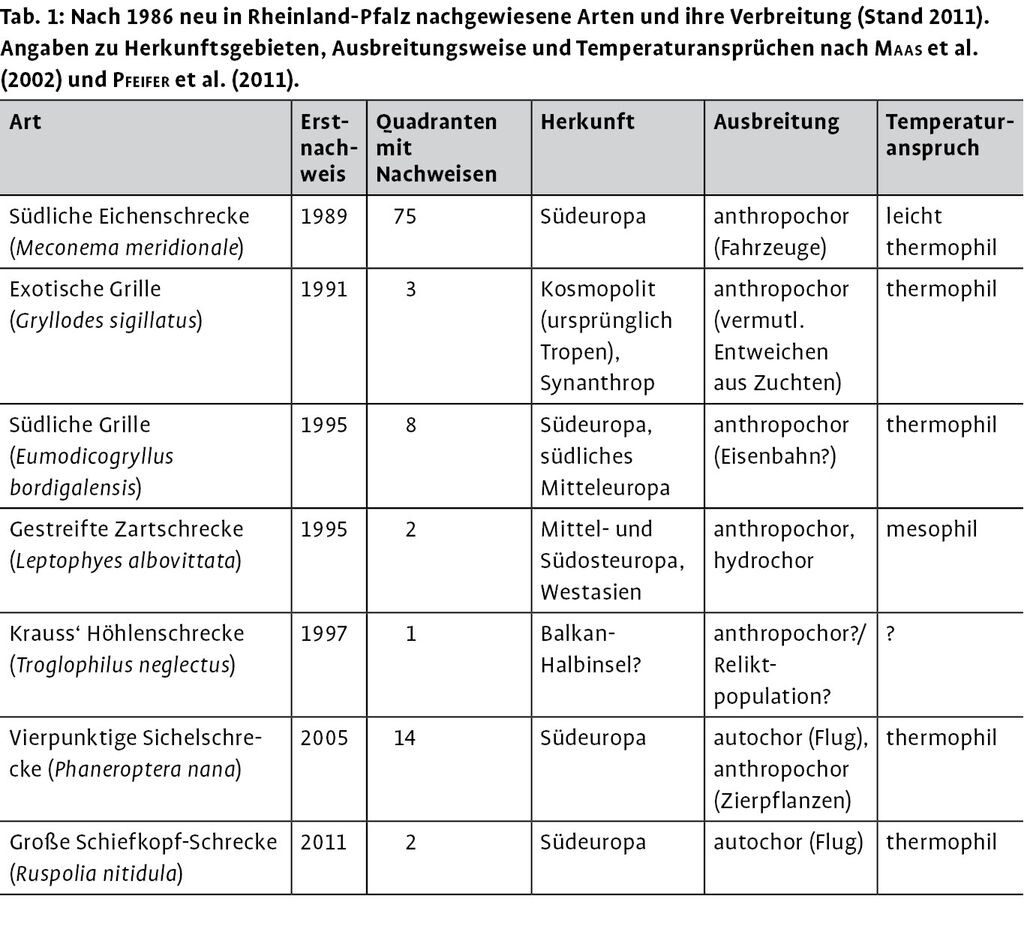
Die systematische Erfassung der Fang- und Heuschreckenfauna begann in Rheinland-Pfalz ab 1987 und wurde von Pfeifer et al. (2011) in einem Verbreitungsatlas dokumentiert. Sie führen in ihrem Atlas 58 aktuell (mit Nachweisen nach 1986) vorkommende bodenständige und fünf ausgestorbene Arten auf. Pfeifer et al. (2011) stuften die erstmals 2005 sowohl in Baden-Württemberg als auch in Rheinland-Pfalz festgestellte Chinesische Gottesanbeterin (Tenodera sinensis) bei den wildlebenden Arten ein und nannten daher 59 bodenständige, seit 1987 nachgewiesene Arten. Offenbar ist das wilde Vorkommen dieser Fangschreckenart aber wieder erloschen, denn sie wurde letztmals 2006 in Rheinland-Pfalz und letztmals 2008 in Baden-Württemberg nachgewiesen (Ehrmann & Reinhardt 2011). T. sinensis wird daher hier nicht mehr zu den bodenständigen Arten gezählt. 2011 kam nach Herausgabe des Werkes mit der Großen Schiefkopf-Schrecke (Ruspolia nitidula) eine weitere Art hinzu (Schirmel & Niehuis 2011, Röller 2011), die vermutlich ebenfalls bodenständig ist. Die für den Verbreitungsatlas herangezogenen Daten, ergänzt um wenige nachträglich bekannt gewordene Funde, wurden in der nachfolgenden Auswertung herangezogen.
Die Fundorte wurden annähernd quadratischen Rasterfeldern mit einer Größe von 3 geografischer Breite und 5 geografischer Länge (TK- oder Messtischblatt-Quadranten), was etwa einer Fläche von 33 km2 je Rasterfeld entspricht, zugeordnet (Pfeifer et al. 2011). Unter einer Meldung wird die Angabe eines Artnamen, des Datums des Nachweises und des Fundortes verstanden. Die Anzahl der vorgefundenen Individuen einer Art bleibt unberücksichtigt.
2.2 Auswertung: Feststellung der Arealerweiterer
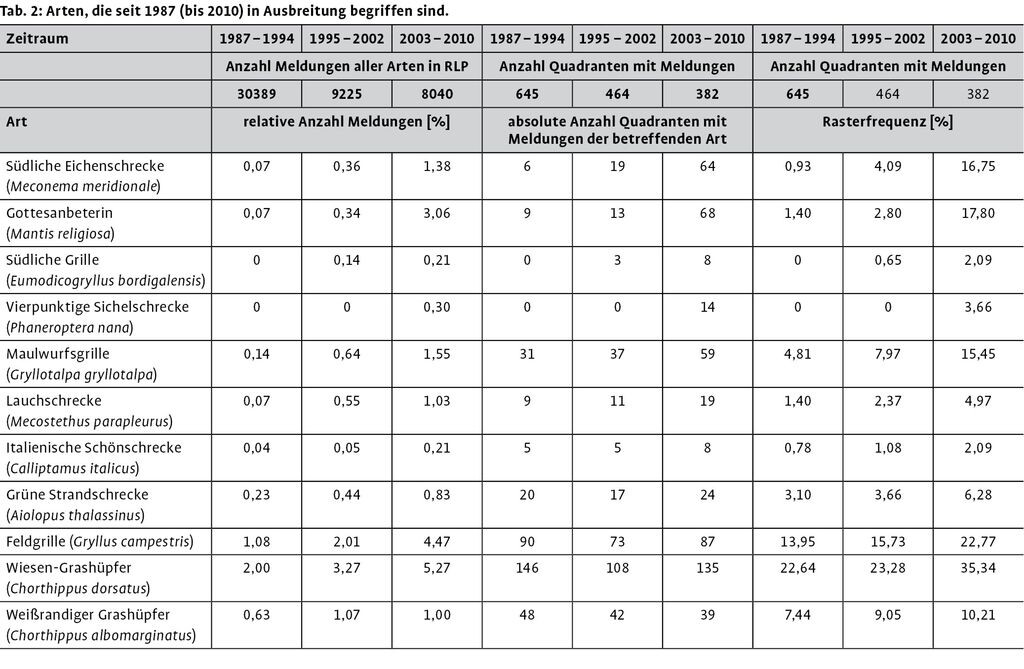
Der gesamte Datenpool mit Meldungen ab 1987 wurde in drei Zeitperioden à acht Jahre aufgeteilt (s. Tab. 2). Da die Anzahl an Meldungen als auch der Quadranten mit Meldungen in den Perioden verschieden ist, ist ein direkter Vergleich nicht zielführend. Die Daten wurden daher relativiert.
Die relative Häufigkeit der Meldungen einer Art ist die Anzahl der Meldungen einer Art dividiert durch die Anzahl der Meldungen aller Arten. Falls kein Anstieg der relativen Häufigkeit einer Art zwischen zwei Zeitschnitten beobachtet wird, bedeutet dieses nicht zwangsläufig, dass die Art seltener geworden ist. Möglich ist auch, dass die Zunahme dieser Art nur unter dem Durchschnitt der anderen Arten liegt. Ein Anstieg der relativen Häufigkeit an den Meldungen deutet zumindest auf eine überdurchschnittliche Ausbreitung hin.
Die Rasterfrequenz ist die Anzahl der Quadranten mit Nachweisen der betreffenden Art dividiert durch die Anzahl aller Quadranten mit Nachweisen mindestens einer Art. Der Rasterfrequenz wird hier die größte Bedeutung beigemessen, dennoch muss bedacht werden, dass in der Regel keine gezielte Suche nach einer Art erfolgte und somit ein Quadrant ohne Nachweis einer Art keine qualitative Aussage darüber ist, dass die Art dort tatsächlich nicht vorkommt.
Von den in Rheinland-Pfalz vorkommenden Arten wurden die ermittelt, deren Rasterfrequenz von der Periode 19871994 zu der nachfolgenden Periode und von dieser Periode ebenfalls zur jüngsten Periode zugenommen hat (Tab. 2).
2.3 Auswertung: Einordnung in nördliche bzw. südliche Arten
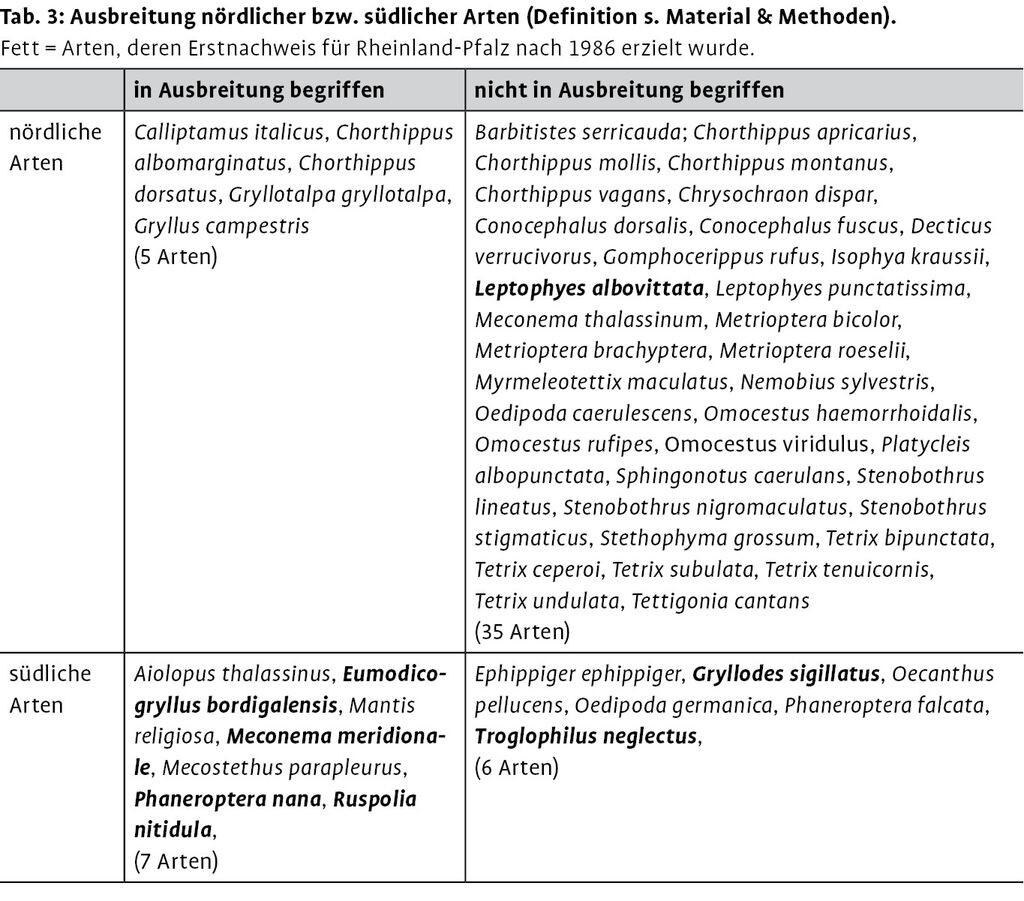
Die aktuell in Rheinland-Pfalz vorkommenden Arten wurden in zwei Verbreitungstypen unterteilt (Tab. 3). Mit südlichen Arten sind hier Taxa gemeint, deren nördliche Arealgrenze in Europa vor 1987 höchstens unwesentlich über 52° nördlicher Breite reichte. Die nördliche Arealgrenze der betreffenden Art verlief somit nicht allzu weit nördlich der Nordgrenze von Rheinland-Pfalz die etwa auf 51° liegt oder sogar durch Rheinland-Pfalz hindurch, insofern die Art nicht erst später vom Süden eingewandert ist. Nördliche Arten kamen demzufolge 1987 bereits auch nördlich des 52sten Breitengrades vor. Die Unterscheidung anhand des Vorkommens südlich oder nördlich des genannten Breitengrades gilt selbstverständlich nur für Rheinland-Pfalz; in weiter nördlich gelegenen Bundesländern würde man eine andere Grenze ziehen. Die hinter der Einteilung stehende Idee ist, dass südlich verbreitete Arten meist wärmebedürftiger als nördlich verbreitete sind und sich infolge einer Klimaerwärmung die Umgebungsbedingungen vor allem für thermophile Arten verbessern.
Das in Häusern lebende Heimchen (Acheta domesticus) wurde nicht berücksichtigt, da es ein vom Freiland-Klima weitgehend unabhängiges Leben führt. Ebenso wurden Arten, die in Rheinland-Pfalz sehr häufig sind (>75 % aller Quadranten mit Nachweisen), nicht einbezogen. Diese Arten haben bereits alle oder nahezu alle potenziellen Habitate besetzt und oft nahezu ihre maximale Ausbreitung erreicht. Bei ihnen handelt es sich ausnahmslos um nördliche Arten. Selbst wenn ein verändertes Klima zu einer Verbesserung der Habitatqualität führen würde, wäre dieser Effekt daher schwerlich durch eine Ausbreitung in weitere Quadranten feststellbar, da schon nahezu alle Quadranten besetzt sind.
Die Hypothese, dass der Anteil an in Ausbreitung befindlichen Arten unter den südlichen signifikant (Irrtumswahrscheinlichkeit <5 %) höher ist, wurde mit dem Exakten Test nach Fisher getestet (Agresti 1992).
3 Ergebnisse
Von den 59 aktuell vorkommenden Arten wurden sieben erstmals nach 1986 nachgewiesen (Tab. 1). Krauss Höhlenschrecke (Troglophilus neglectus) wurde möglicherweise lange übersehen (Pfeifer et al. 2011). Die übrigen sechs Arten sind mit sehr hoher Wahrscheinlichkeit neu eingewandert. Fünf dieser Arten gelten als thermophil und haben eine südeuropäische oder tropische Herkunft. Bei der Vierpunktigen Sichelschrecke (Phaneroptera nana) ist der Erstnachweis für Rheinland-Pfalz erst vor wenigen Jahren erfolgt, doch hat ihr Bestand bereits merklich zugenommen. Bei der erst 2011 an zwei Standorten entdeckten Großen Schiefkopf-Schrecke (Ruspolia nitidula) liegen zwar aus Rheinland-Pfalz erst seit Neustem Hinweise auf eine Arealexpansion vor, aber Beobachtungen aus anderen europäischen Regionen (Hugel 2006, Holua et al. 2007a, Kritin et al. 2007) lassen darauf schließen, dass diese Art sich auch in Rheinland-Pfalz vermehrt ausbreiten wird.
Von den 52 bereits vor 1987 entdeckten und aktuell in Rheinland-Pfalz wildlebenden Arten wurden acht weitere Arten ermittelt, deren Rasterfrequenz in den zwei Folgeperioden nacheinander angestiegen sind. Zusammen mit den erst nach 1987 festgestellten Arten werden somit zwölf Arten als in kontinuierlicher Ausbreitung begriffen angesehen (Tab. 3). Bei acht Arten erfolgte trotz abnehmender Kartierungsintensität (sinkende Anzahl von Meldungen aller Arten in den drei Perioden) sogar eine absolute Zunahme der Anzahl von Quadranten mit Nachweisen. Bei zehn Arten nahm außerdem die relative Häufigkeit der Meldungen zu.
Von 13 Spezies, die als südliche Arten eingestuft wurden, sind sieben in Ausbreitung begriffen, bei den nördlichen Arten sind dieses nur fünf bei einer Gesamtanzahl von 40 nördlichen Arten (Tab. 3). Der Anteil an in Ausbreitung befindlicher Arten ist unter den südlichen Arten signifikant höher.
Lässt man die Arten unberücksichtigt, die nach 1986 vermutlich eingeschleppt wurden Südliche Eichenschrecke (Meconema meridionale), Südliche Grille (Eumodicogryllus bordigalensis), Exotische Grille (Gryllodes sigillatus), Gestreifte Zartschrecke (Leptophyes albovittata) oder deren Herkunft bisher ungeklärt ist Krauss Höhlenschrecke (Troglophilus neglectus) , ist der Anteil südlicher in Ausbreitung befindlicher Arten immer noch signifikant höher.
Wenn man allerdings alle erst nach 1986 neu für Rheinland-Pfalz nachgewiesene Arten (Tab. 1) unberücksichtigt lässt, ist ein signifikanter Unterschied nicht mehr feststellbar, denn die Irrtumswahrscheinlichkeit läge bei etwa 9 %.
4 Diskussion
4.1 Überblick
Wie die Studie zeigt, hat die Anzahl der in Rheinland-Pfalz bodenständigen Heu- und Fangschreckenarten seit 1987 durch Einwanderung und Einschleppung von sieben, vorwiegend südlichen (thermophilen) Spezies deutlich zugenommen. Außerdem wurde bei einem signifikant höheren Anteil südlicher als nördlicher Arten eine Ausbreitung festgestellt.
Interessanterweise wurde bei der Gemeinen Sichelschrecke (Phaneroptera falcata) und dem Weinhähnchen (Oecanthus pellucens) beides südliche Arten keine aufeinanderfolgende Zunahme in den Perioden ab 1995 nachgewiesen, obwohl diese Arten sich in den Jahren zuvor ausbreiteten und eine Ausbreitung in Nordrhein-Westfalen, Niedersachsen und den Niederlanden aktuell beobachtet wird (Behrens et al. 2009a, b; Handke et al. 2011, Pfeifer et al. 2011). Offenbar haben diese beiden Arten in Rheinland-Pfalz bereits den größten Teil der potenziellen Habitate, insbesondere entlang der großen Flusstäler, besetzt. Eine weitere Ausbreitung erfolgt nun in die Höhenlagen, die anscheinend in geringerem Tempo vonstatten geht.
In Rheinland-Pfalz beherbergen wie auch andernorts die wärmsten Gebiete gleichzeitig die höchste Anzahl an Fang- und Heuschreckenarten (Pfeifer et al. 2011). Der Schluss, dass durch die Klimaerwärmung bisher vergleichsweise kühle Lebensräume für thermophile Arten erschlossen werden, liegt daher nahe und steht in Übereinkunft mit den festgestellten Ausbreitungen. Doch kommen auch andere Ursachen in Betracht, die eine klimagesteuerte Zunahme der Artenanzahl und der Ausbreitung südlicher Arten infrage stellen?
4.2 Folge einer andauernden postglazialen Ausbreitung?
Höhere, kältere Breitengrade sind in der Regel artenärmer als niedrigere, wärmere. Die Ursachen für diesen Zusammenhang sind komplex und im Detail oft nicht geklärt (Currie et al. 2004). Dieses Nord-Süd-Gefälle besteht aber zweifellos und ist an Heuschrecken auf kleinräumiger Skala in Deutschland (Maas et al. 2002) und sogar innerhalb von Rheinland-Pfalz (Pfeifer et al. 2011) festzustellen. Neben klimatischen Ursachen für ein Nord-Süd-Gefälle kommt auch infrage, dass die postglaziale Wiederbesiedlung noch nicht abgeschlossen ist, was Svenning & Skov (2007) für Baumarten diskutieren, die ein geringes Migrationspotential (<100 m/a) haben.
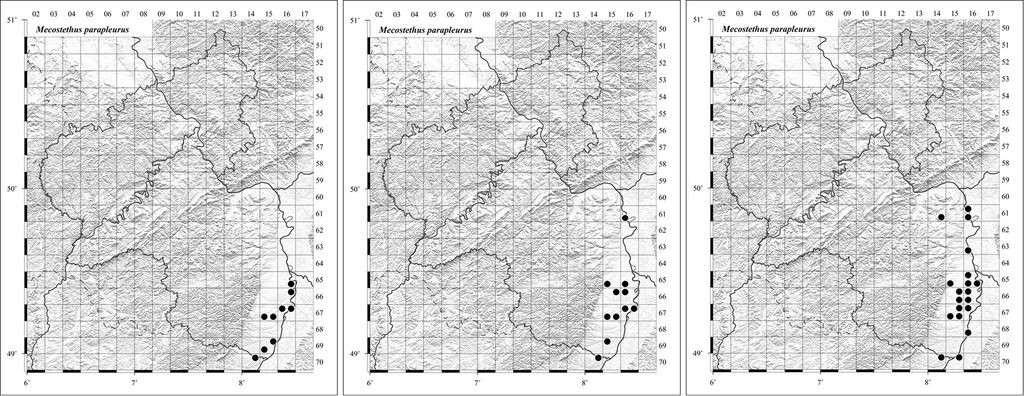
Bei sich schneller ausbreitenden Arten ist in Mitteleuropa weitgehend davon auszugehen, dass die Arten bereits vor längerer Zeit ihre nördliche klimatische Grenze erreicht haben. Bei den in Rheinland-Pfalz in nördlicher Ausbreitung begriffenen Heuschreckenarten ist die postglaziale Wiederbesiedlung bei den flugfähigen südlichen Arten Grüne Strandschrecke (Aiolopus thalassinus), Lauchschrecke (Mecostethus parapleurus) (Abb. 1, 2), Gemeine Sichelschrecke (Phaneroptera falcata) , die ein hohes Migrationspotential haben, abgeschlossen bzw. sie reagieren auf klimatische Änderungen direkt mit der Verschiebung vor allem der nördlichen Arealgrenze. Diese Arten haben in Rheinland-Pfalz ihren Verbreitungsschwerpunkt im klimatisch begünstigen Oberrheintiefland.
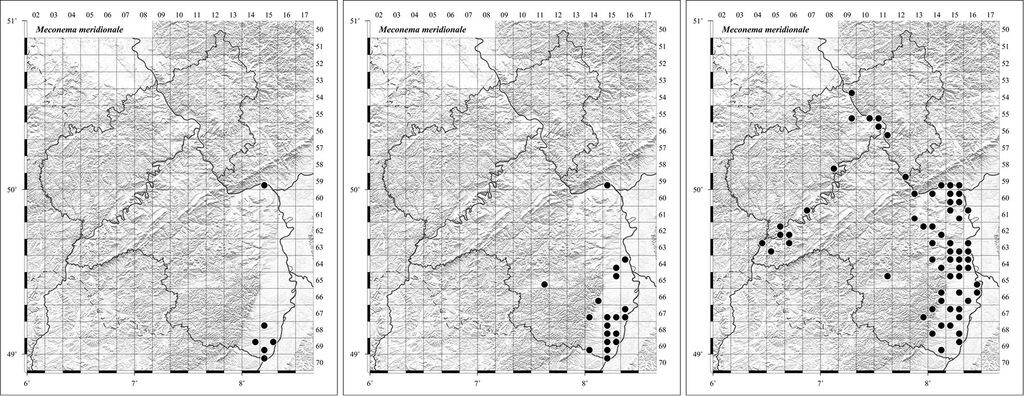
Als potenziell in Rheinland-Pfalz in postglazialer Arealerweiterung befindlich kommen vermutlich nur Arten in Frage, die flugunfähig sind und auch niemals gelegentlich flugfähige Formen auszubilden vermögen; bei allen anderen Arten dürfte das Migrationspotenzial zu hoch sein. Unter den festgestellten Arealerweiterern ist die Südliche Eichenschrecke (Meconema meridionale) flugunfähig. Sie stellt an die Temperatur keine ausgesprochen hohen Ansprüche, weshalb die Art gut mit den klimatischen Verhältnissen in Rheinland-Pfalz zu Recht kommt. Warum also war diese erstmals 1989 in Rheinland-Pfalz nachgewiesene Art nicht schon längst bodenständig?
Um aus eigener Kraft von ihrem adriatomediterranen Glazialrefugium bis heute nach Rheinland-Pfalz zu gelangen, hätte sie seit Ende des Pleistozäns ca. 50 m/a zurücklegen müssen. Bei einem Migrationspotential von >100m/a würde sie bereits über Rheinland-Pfalz hinausgewandert sein. Die heute beobachtete Ausbreitungsgeschwindigkeit der Art liegt im Bereich von Dutzenden km/a und ist anthropogen bedingt (s.u.). Ein Migrationspotenzial von <100 m/a, welches für ein durch andauernde postglaziale Arealexpansion getriebene Ausbreitung in Rheinland-Pfalz in Frage kommt, dürfte in dem hier in Betracht gezogenen Zeitraum eine kaum merklich Zunahme verursachen. Vielmehr ist die hier festgestellte Zunahme der flugunfähigen Arten (Leptophyes albovittata und Meconema meridionale) eindeutig eine Folge anthropogener Verschleppung.
4.3 Folge der Einschleppung und Verschleppung?
Unter allen nach 1987 arealerweiternden Fang- und Heuschrecken in Rheinland-Pfalz ist die Südliche Eichenschrecke (Meconema meridionale) die erfolgreichste Art (Abb. 3, 4). Seit den 1990er-Jahren breitet sich die Art enorm aus und hat bereits England erreicht. Dabei ist M. meridionale flugunfähig und wäre aus eigener Kraft nicht zu solch einer Expansion in der Lage gewesen (s.o.). Erwiesenermaßen lässt sie sich von Autos und Zügen mitnehmen und vermag auf diese Weise weite Strecken zurückzulegen. Deutliche Hinweise auf die Verschleppung liefert auch das plötzliche Auftreten der Art besonders in Städten fernab ihrer zuvor bekannten Verbreitungsgrenzen, während sie aus ländlicher Umgebung meist erst später und weniger häufig gemeldet wird. Die Arealexpansion ist daher zweifellos eine Folge des zunehmenden Verkehrsaufkommens (Pfeifer et al. 2011). Vermutlich begünstigte eine Verbesserung der Klimabedingungen den Boden für das invasive Auftreten der Art (s.a. Walther et al. 2009).
Die Südliche Grille (Eumodicogryllus bordigalensis) wurde erstmals 1995 für das Bundesland in dem im Süden von Rheinland-Pfalz liegenden Bahnhof Wörth nachgewiesen. Die Art wurde mittlerweile an Fundorten angetroffen, an denen sie nachweislich zuvor nicht vorkam. Somit ist eine Ausbreitung bewiesen, auch wenn sie vergleichsweise zur vorherig genannten Art gering ist. Aktuell ist auch andernorts eine Ausbreitung feststellbar, beispielsweise in Österreich (Zuna-Kratky et al. 2009), der Schweiz, dem nördlichen Frankreich und der Tschechischen Republik (Pfeifer et al. 2011). Alle Funde in Rheinland-Pfalz wurden ausschließlich im Gleisschotter erbracht, was auf eine Vorliebe heißer Standorte hindeutet. Die Fundumstände in Basel, Weil am Rhein und in der Tschechischen Republik (Holua et al. 2007 b) sind identisch mit denen in Rheinland-Pfalz.
Ursprünglich ist die Art in Südeuropa und dem südlichen Mitteleuropa zuhause (Harz 1969). Maas et al. (2002) und Pfeifer et al. (2011) sehen E. bordigalensis für Deutschland daher als ein Neozoon an, das eventuell mit Zügen eingeschleppt wurde. Da insgesamt eine Arealerweiterung nach Norden stattzufinden scheint (Pfeifer et al. 2011), vermuten die Autoren, dass diese durch die Klimaerwärmung wie bei der Südlichen Eichenschrecke zumindest begünstigt wird.
Die Exotische Grille (Gryllodes sigillatus) wurde vielfach nach Mitteleuropa eingeschleppt, konnte sich zuvor aber nie im Freiland halten (Harz 1969). Da ihr dieses nun im klimatisch begünstigen Oberrheintiefland in Rheinland-Pfalz gelang, spricht dafür, dass sich das Klima für die Art verbessert hat. Die Art ist nach wie vor sehr selten, eine weitere Ausbreitung praktisch nicht feststellbar.
Die Vierpunktige Sichelschrecke (Phaneroptera nana) wurde 2005 erstmalig in Rheinland-Pfalz nachgewiesen. Bei der Art geht Harz (1969) von einer ursprünglich zirkummediterranen Verbreitung aus. Derzeit wird auf breiter Front eine Ausbreitung nach Norden beobachtet, was auf eine klimagetriebene Arealexpansion hindeutet. Anzunehmen ist, dass P. nana mit Pflanzenmaterial anthropogen verfrachtet wird (Pfeifer et al. 2011). In welchem Ausmaß das zur derzeitigen Arealexpansion beiträgt, ist unbekannt. Anders aber als M. meridionale ist P. nana flugfähig und es liegen keine Hinweise vor, dass sie sich in ihren Fähigkeiten zur aktiven Migration von ihrer Schwesterart Phaneroptera falcata unterscheidet. P. falcata ist in ihren Temperaturansprüchen lediglich weniger anspruchsvoll, ebenfalls in nördlicher Ausbreitung begriffen und bereits weiter nach Norden vorgestoßen (Pfeifer et al. 2011).
Die aus Osteuropa kommende Gestreifte Zartschrecke (Leptophyes albovittata) dehnt ihr Areal derzeit westwärts aus (Pfeifer et al. 2011) und kommt in Norddeutschland bereits an der Elbe vor. Die anthropochor und hydrochor verfrachtete Art ist außerdem nicht ausgesprochen thermophil. L. albovittata scheint daher nach Meconema meridionale ein weiteres Beispiel für eine anthropogen unterstützt arealerweiternde Art zu sein, bei der es in Rheinland-Pfalz keiner Beihilfe durch ein verändertes Klima bedurfte, d.h. die Art hätte sich vermutlich schon viel früher und ohne Klimaerwärmung etabliert, wenn sie früher eingeschleppt worden wäre. In Rheinland-Pfalz ist die als kryptisch zu bezeichnende L. albovittata bisher lediglich von zwei Fundorten bekannt. Von einer nennenswerten Ausbreitung kann daher bis heute nicht gesprochen werden.
Die Gottesanbeterin (Mantis religiosa) gehört zu den Arten aus den beiden untersuchten Insektenordnungen, von der Vorkommen auf dem Territorium von Rheinland-Pfalz bereits seit dem Beginn des 19. Jahrhunderts bekannt sind. Die bis in die 1920er-Jahre bekannten Fundorte liegen ausnahmslos in den wärmsten Regionen von Rheinland-Pfalz und die Art wurde immer als Rarität angesehen (Pfeifer et al. 2011). Seit etwa 1990 breitet sich M. religiosa von Süden her kommend auf breiter Front nordwärts aus und besiedelt dabei nicht nur das wärmebegünstigte nördliche Oberrheintiefland, sondern dringt in die höher gelegenen südlichen, offeneren Lagen des Haardtgebirges (Pfälzerwald) ein.
Ein Großteil der Zunahme an Meldungen aus Rheinland-Pfalz ist das Ergebnis von Aufrufen in der Tagespresse, ohne die der in Tab. 2 dargestellte rasante Anstieg an besetzten Quadranten nach 2002 nicht in dieser Höhe ausgefallen wäre. Doch sind diese Meldeaufrufe bei Weitem nicht allein dafür verantwortlich, denn unabhängig davon stellen auch ortskundige Entomologen eine starke Abundanzzunahme fest. Insgesamt ist in Europa eine Ausbreitung in nördlicher Richtung feststellbar (Berg et al. 2011).
Bei M. religiosa spielt anthropogene Verfrachtung ebenfalls eine Rolle. Sie erfolgt ungewollt oder oftmals auch absichtlich als Oothek, Larve und Imago. Funde fernab der Arealgrenzen, z.B. in Berlin, Brandenburg oder Braunschweig, sind nur durch anthropogene Verfrachtung erklärbar (Berg et al. 2011, Ehrmann & Reinhardt 2011, Pfeifer et al. 2011). Merkmal ihrer anthropogen erfolgten Ausbreitung ist wie auch bei Meconema meridionale zu beobachten , dass zerstreut Satellitenpopulationen entstehen, meist in Städten und vielfach weit entfernt von den nächsten Populationen. Die derzeit erfolgende Wiedereroberung des Südens von Rheinland-Pfalz auf breiter Front scheint daher nicht allein diese Ursache zu haben, sondern dürfte überwiegend das Produkt einer aktiven Arealausdehnung dieser flugfähigen Mantide sein.
Zusammenfassend gesagt, spielt anthropochore Verbreitung eine wichtige Rolle besonders bei neu auftretenden Arten und ist konkret oft nicht eindeutig von klimatisch begünstigter Ausbreitung zu unterscheiden. Dass insbesondere südliche, von einer Klimaerwärmung profitierende Arten anthropochor sind, dürfte oftmals daran liegen, dass die meisten autochoren Arten nur relativ langsam wandern und deshalb noch nicht die (mit der aktuellen Klimaerwärmung verschobene) klimatisch mögliche nördliche Arealgrenze erreichten. Dass aber auch autochore Arten derzeit weiter nach Norden vordringen, zeigt das Beispiel der gut flugfähigen Großen Schiefkopf-Schrecke sowie südlicher Libellenarten (Ott 2010).
4.4 Fluktuation oder andere nicht klimatische Faktoren?
Wie bei der Gottesanbeterin ist es bei der Maulwurfsgrille (Gryllotalpa gryllotalpa) aufgrund von Presseaufrufen im Jahr 2006 zu einer starken Zunahme von Meldungen gekommen (Pfeifer 2007). Die Anzahl an Meldungen von G. gryllotalpa ab 2003 erreichte aber nur etwa die Hälfte der Anzahl von Meldungen von M. religiosa, obwohl G. gryllotalpa in den Perioden zuvor häufiger gemeldet wurde.
Dieser Vergleich unterstreicht nochmals die Feststellung, dass die zu beobachtende Arealerweiterung von M. religiosa kein methodisches Artefakt ist. Bei der Maulwurfsgrille ist aber ebenso wie bei der Gottesanbeterin schon in der Periode vor dem Meldeaufruf trotz sinkender Anzahl von Meldungen insgesamt (Tab. 2) eine absolute Zunahme der Anzahl Quadranten feststellbar; ein Indiz dafür, dass sich die Art derzeit in Rheinland-Pfalz ausbreitet. Die Art kommt vor allem in den Wärmegebieten von Rheinland-Pfalz vor; durch eine Klimaerwärmung sollte daher das potenzielle Areal für die Art in die höheren und nördlichen Regionen des Bundeslandes ausgedehnt werden. In Nordrhein-Westfalen prognostiziert man ebenfalls eine Ausdehnung der Art (Behrens et al. 2009 a, b). In Niedersachsen (Grein 2010) hingegen wird von einem Rückgang der Art ausgegangen, was mit zunehmender Intensität des Ackerbaus begründet wird. Äcker beherbergen aber in Rheinland-Pfalz schon lange keine Populationen der Art mehr; die Meldungen der Art in Rheinland-Pfalz konzentrieren sich vielmehr auf Gärten (Pfeifer et al. 2011). Möglicherweise ist daher die neuerdings zu beobachtende Zunahme in Rheinland-Pfalz teilweise auf einen verringerten Umfang der Bekämpfung dieses Gartenschädlings zurückzuführen; die Maulwurfsgrille kehrt in ihre zuvor weggenommenen Habitate zurück.
Größen und Ausbreitung von Populationen unterliegen einer Dynamik, die nicht klimatisch gesteuert sein müssen. Das berühmteste Beispiel hierfür sind Lemminge in Skandinavien. Bei Heuschrecken sind ebenfalls Fluktuationen der Populationsgrößen zu beobachten, die zumindest teilweise mit Wettereinflüssen erklärt werden (beispielsweise Köhler et al. 1999). Merkmal der nicht durch Klimaänderung in eine Richtung getriebenen Fluktuationen ist, dass sie über längere Zeiträume gesehen keinen Trend aufweisen, sondern um einen Mittelwert pendeln. Ott (2010) bezeichnet dieses deshalb als Oszillation. Die hier ausgewerteten Daten gehen noch nicht so weit zurück, als dass dieser Mittelwert bei lediglich oszillierenden Arten schon bekannt wäre. Daher besteht immer noch die Möglichkeit, dass bei einer einzelnen Art ein Trend lediglich vorgetäuscht wird. Über Fluktuationen sind aber nicht die Unterschiede zwischen der Gruppe der südlichen und der nördlichen Arten zu erklären.
5 Schlussfolgerung
Dass die Verbreitung auch einiger nördlichen Arten in Rheinland-Pfalz zunimmt, stellt keinen Widerspruch zu der Hypothese einer durch die Klimaerwärmung begünstigen Ausbreitung von Fang- und Heuschreckenarten dar, denn auch für viele Vertreter dieser Gruppe werden die Standortbedingungen günstiger. Ausschlaggebend für die Annahme einer durch die Klimaerwärmung getriebenen Ausbreitung ist, dass südliche, thermophile Arten proportional stärker profitieren. Ein hoher Anteil der derzeit neu auftretenden südlichen Arten ist anthropochor. Diese Tatsache dürfte darauf zurückzuführen sein, dass rein autochore Arten nicht mit den hohen Migrationsdistanzen anthropochorer Arten mithalten können. Klimatisch bedingte Ausbreitung südlicher Arten wird durch Verschleppung unterstützt, Verschleppung alleine hingegen scheint hingegen nicht als alleinige Ursache für die nordwärts gerichtete Expansion der Mehrzahl südlicher Arten in Frage zu kommen.
Wie Steck et al. (2007) bereits bei Heuschrecken in der Schweiz feststellten, gehört die Mehrzahl der Arten der Ordnung nebst der Gottesanbeterin offenbar auch in Rheinland-Pfalz zu den Gewinnern des Klimawandels.
Dank
Die hier vorliegende Auswertung basiert auf Daten von weit über 200 ehrenamtlichen Meldern, die in einem Verbreitungsatlas zusammengefasst wurden (Pfeifer et al. 2011). Ohne diese breite Hilfe bestünde die Grundlage für die hier getroffenen Aussagen nicht. Besonderen Dank gilt den Mitherausgebern und Mitautoren an diesem Atlas: Manfred Niehuis, Carsten Renker, Reinhard Ehrmann, Andreas Kiefer, Frank Schlotmann, Tom Schulte, Ludwig Simon, Ansgar van Elst und besonders Wolfgang Fluck, der die Verbreitungskarten erstellte. Ebenfalls besonders dankt der Autor Thomas Fartmann von Institut für Landschaftsökologie an der Universität Münster; von ihm kam die Anregung zu diesem Artikel. Robin Gordon (Berlin) übernahm freundlicherweise die Korrektur der englischsprachigen Zusammenfassung.
Literatur
Agresti, A. (1992): A survey of exact interference for contingency tables. Statistical science 7 (1), 131-153.
Behrens, M., Fartmann, T., Hölzel, N. (2009a): Auswirkungen von Klimaveränderungen auf die biologische Vielfalt: Pilotstudie zu den Auswirkungen des Klimawandels auf ausgewählte Tier- und Pflanzenarten in Nordrhein-Westfalen. Teil 1: Fragestellung, Klimaszenario, erster Schritt in der Empfindlichkeitsanalyse Kurzprognose. Münster, 288 S.
, Fartmann, T., Hölzel, N. (2009b): Auswirkungen von Klimaveränderungen auf die biologische Vielfalt: Pilotstudie zu den Auswirkungen des Klimawandels auf ausgewählte Tier- und Pflanzenarten in Nordrhein-Westfalen. Teil 2: zweiter Schritt der Empfindlichkeitsanalyse Wirkprognose. Münster, 364 S.
Berg, M.K., Schwarz, C.J., Mehl, J.E. (2011): Die Gottesanbeterin. Westarp, Hohenwarsleben, 521 S.
Buse, J., Griebeler, E.M. (2011): Incorporating classified dispersal assumptions in predictive distribution models A case study with grasshoppers and bush-crickets. Ecological modelling 222 (13), 2130-2141.
Currie, D.J, Mittelbach, G.G, Cornell, H.V., Field, R., Guégan, J.-F., Hawkins, B.A., Kaufman, D.M., Kerr, J.T., Oberdorff, T, OBrien, E., Turner, J.R.G. (2004): Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness. Ecology letters 7, 1121-1134.
Detzel, P., Maas, S., Staudt, A. (2005): Heuschrecken (Saltatoria). 331-350 + Anh. In: Günther, A., Nigmann, U., Achtziger, R., Gruttke, H., Analyse der Gefährdungsursachen planungsrelevanter Tiergruppen in Deutschland, Naturschutz und Biologische Vielfalt 21.
Ehrmann, R., Reinhardt, R. (2011): Fauna der Fangschrecken (Mantodea) Sachsens. Mitt. Sächs. Entomologen, Suppl. 9, 82-96.
Grein, G. (2010): Fauna der Heuschrecken (Ensifera & Caelifera) in Niedersachsen. Naturschutz und Landschaftspflege in Niedersachsen 46, 1-183.
Handke, K., Horstkotte, J., Kleinkuhle, J. (2011): Neue Funde der Sichelschrecke Phaneroptera falcata (Poda, 1761) in Nordwestdeutschland. Articulata 26 (2), 163-167.
Harz, K. (1969): Die Orthopteren Europas I. Dr. W. Junk N.V., The Hague, 749S.
Holua, J., Kocárek, P., Marhoul, P. (2007a): First sightings of Ruspolia nitidula (Orthoptera: Tettigoniidae) and Mecostethus parapleurus (Orthoptera: Acrididae) after fifty years in the Czech Republic. Articulata 22 (1), 47-51.
, Kocárek, P., Vlk, R., Marhoul, P. (2007b): Southern Cricket Eumodicogryllus bordigalensis (Orthoptera: Gryllidae) in the Czech Republic: new records and notes on the biology and stridulation. Polish journal of ecology 76 (1), 47-55.
Hugel, S. (2006): Présence de la Sauterelle Ruspolia nitidula (Scopoli, 1786) en Alsace (Orthoptera, Ensifera). Bulletin de la Societé entomologique de Mulhouse 62 (3), 48-49.
Kleukers, R.M.J.C. (1997): Northward expansion of Orthoptera in Western Europe. Proceedings of the International Colloquium of the European Invertebrate Survey, Saarbrücken, 6.-7. July 1995, 29-36.
Köhler, G., Perner, J., Schumacher, J. (1999): Grasshopper population dynamics and meteorological parameters lesson from a case study. Ecography 22, 205-212.
Kritin, A., Kanuch, P., Sárossy, M. (2007): Did the northern range of distribution of two tropical Orthopterans (Insecta) change recently? Polish journal of ecology 55 (2), 297-304.
Lähne, W. (2008): Klimawandel in der Pfalz Entwicklungen und Trends Regionale Zukunftsszenarien. Pollichia-Kurier 24 (2), 4-9.
Leuschner, C., Schipka, F. (2004): Vorstudie Klimawandel und Naturschutz in Deutschland. BfN-Skripten 115, 1-35.
Maas, S., Detzel, P., Staudt, A. (2002): Gefährdungsanalyse der Heuschrecken Deutschlands. Bundesamt für Naturschutz, Bonn-Bad Godesberg, 401 S.
Ministerium für Umwelt, Forsten und Verbraucherschutz Rheinland-Pfalz (2007): Klimabericht Rheinland-Pfalz 2007. Mainz, 184S.
Ott, J. (1996): Zeigt die Ausbreitung der Feuerlibelle in Deutschland eine Klimaveränderung an? Mediterrane Libellen als Indikatoren für Änderungen in Biozönosen. Naturschutz und Landschaftsplanung 28 (2), 53-61.
Ott, J. (2010): Dragonflies and climatic change recent trends in Germany and Europe. BioRisk 5, 253-286.
Pfeifer, M.A. (2007): Schätzung des Kartierungsstandes und Umfangs der Verbreitung der Maulwurfsgrille (Gryllotalpa gryllotalpa) in der Pfalz. Fauna und Flora in Rheinland-Pfalz 11 (1), 55-70.
, Niehuis, M., Renker, C. (2011): Die Fang- und Heuschrecken in Rheinland-Pfalz. Landau, 679S.
Poniatowski, D., Fartmann, T. (2011): Weather-driven changes in population density determine wing dimorphism in two bush-cricket species. Agriculture, Ecosystems and Environment 145, 59.
Rabitsch, W., Winter, M., Kühn, E., Kühn, I., Götzl, M., Essl, F., Gruttke, H. (2010): Auswirkungen des rezenten Klimawandels auf die Fauna in Deutschland. Naturschutz und Biologische Vielfalt 98, 265S.
Robinet, C., Roques, A. (2010): Direct impact of recent climate change on insect populations. Integrative Zoology 5 (2), 132-142.
Röller, O. (2011): Ein Nachweis der Großen Schiefkopfschrecke (Ruspolia nitidula) am Ortsrand von Haßloch. Pollichia-Kurier 27 (4), 36.
Schirmel, J., Niehuis, M. (2011): Nachweis der Großen Schiefkopfschrecke Ruspolia nitidula (Scopoli, 1786) in Rheinland-Pfalz. Fauna und Flora in Rheinland-Pfalz 12 (1), 311-314.
Steck, C.E., Bürgi, M., Bolliger, J., Kienast, F., Lehmann, A., Goseth, Y. (2007): Conservation of grasshopper diversity in a changing environment. Biological conservation 138, 360-370.
Svenning, J.-C., Skov, E. (2007): Could the tree diversity pattern in Europe be generated by postglacial dispersal limitation? Ecology letters 10, 453-460.
Walther, G.-R., Roques, A., Hulme, P.E., Sykes, M. T., Pyek, P., Kühn, I., Zobel, M., Bacher, S., Botta-Dukát, Z., Bugmann, H., Czúcz, B., Dauber, J., Hickler, T., Jaroik, V., Kenis, M., Klotz, S., Minchin, D., Morra, M., Nentwig, W., Ott, J., Panov, V. E., Reineking, B., Robinet, C., Semenchenko, V., Solarz, W., Thuiller, W., Vilá, M., Vohland, K., Settele, J. (2009): Alien species in a warmer world: risks and opportunities. Trends in ecology and evolution 24 (12), 686-693.
Wilson, R.J., Maclean, I.M.D. (2011): Recent evidence for the climate change threat to Lepidoptera and other insects. Journal of insect conservation 15, 259-268.
Wissmann, J., Schielzeth, H., Fartmann, T. (2009): Landscape-scale expansion of Roesels bush-cricket Metrioptera roeselii at the north-western range limit in central Europe (Orthoptera: Tettigoniidae). Entomologia Generalis 31 (4), 317-326.
Zuna-Kratky, T., Karner-Ranner, E., Lederer, E., Braun, B., Berg, H.M., Denner, M., Bieringer, G., Ranner, A., Zechner, L. (2009): Verbreitungsatlas der Heuschrecken und Fangschrecken Ostösterreichs. Verlag des Naturhistorischen Museum Wien, Wien, 303S.
Anschrift des Verfassers: Manfred Alban Pfeifer, Arbeitskreis Heuschrecken in der Gesellschaft für Naturschutz und Ornithologie Rheinland-Pfalz, Bahnhofsplatz 5, 67240 Bobenheim-Roxheim. Email: heuschrecken.rlp@googlemail.com .
-
Abb. 1: Die flugfähige Lauchschrecke (Mecostethus parapleurus). Foto: Oliver Niehuis -
Abb. 2: Nachweise der Lauchschrecke (Mecostethus parapleurus) in Rheinland-Pfalz, (a) 1987 1994, (b) 1995 2002, (c) nach 2002. Beispiel für eine Art, die langsam, aber selbstständig ihr Areal nach Norden erweitert. -
Abb. 3: Die flugunfähige Südliche Eichenschrecke (Meconema meridionale). Foto: Oliver Röller -
Abb. 4: Nachweise der Südlichen Eichenschrecke (Meconema meridionale) in Rheinland-Pfalz, (a) 1987 1994, (b) 1995 2002, (c) nach 2002. Beispiel für eine eingeschleppte Art, die ihr Areal massiv und fast ausschließlich anthropochor erweitert. -
Heuschrecken und Klimawandel -
Heuschrecken und Klimawandel -
Heuschrecken und Klimawandel


Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.