Ausbreitung und Management von Götterbaum und Robinie in Niederwäldern
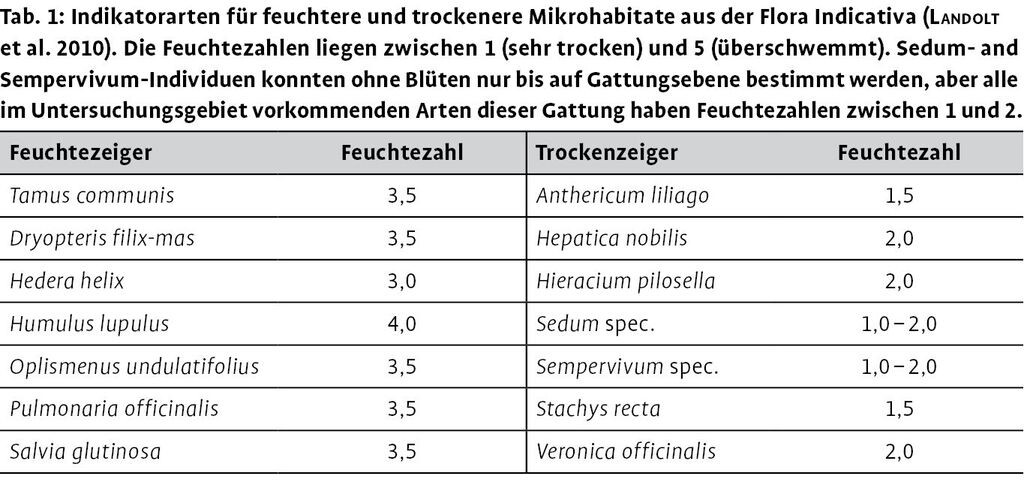
Abstracts
Die Robinie (Robinia pseudoacacia L.) und der Götterbaum (Ailanthus altissima (Mill.) Swingle) sind Neophyten, die europaweit anthropogene und naturnahe Habitate besiedeln. Ihre Invasion in Waldökosysteme ist bislang kaum untersucht worden. In Südtirol wanderten beide Arten in aktiv bewirtschaftete Niederwälder ein. Um die Entwicklung der Invasion nachzuvollziehen, wurde das Vorkommen beider Arten in einem Niederwald untersucht. Dazu wurde die Baumartenzusammensetzung der Waldbestände einschließlich der Verjüngung erfasst und mit dem Bestandesalter (Zeit seit letztem Kahlhieb) und den Standortfaktoren verglichen.
Die Ergebnisse zeigen, dass die Robinie vermutlich schon länger als der Götterbaum im Untersuchungsgebiet vorhanden ist. Letzterer hat sich jedoch in den vergangenen 30 Jahren deutlich schneller über die Fläche ausgebreitet. Beide Arten besiedeln typischerweise neu entstandene Hiebflächen, wo sie sich durch ihr rasches Wachstum dauerhaft etablieren können. Weil beide Neophyten stark von der klassischen Niederwaldbewirtschaftung profitieren, ist eine Anpassung der Bewirtschaftungspraxis notwendig, um ein Fortschreiten der Invasion zu vermeiden.
Dispersal and Management of Heaven and Black Locust in Coppiced Forests Results of a case study in Southern Tyrol
Throughout Europe, both anthropogenic and near-natural habitats are invaded by black locust (Robinia pseudoacacia L.) and tree of heaven (Ailanthus altissima (Mill.) Swingle). However, their invasion into forests has been rarely studied, particularly the influence of coppice management, which is characterised through regular cut-backs. To retrace the invasion the study investigated the abundance in the canopy as well as the natural regeneration of these species in a coppice in South Tyrol which showed ideal conditions due to its patchy mosaic of different ages (time since last coppicing). Tree regeneration of the invasive and native species was compared among these forest age classes and correlated to environmental parameters. Apparently, R. pseudoacacia has established earlier than A. altissima but A. altissima has spread with higher speed during the last 30 years. Synchronously to the coppice cycle both species colonise fresh clearings where they establish and remain. Since current coppice management favors the spread of both invasive species an adaptation of the management system is necessary to avoid further invasion.
- Veröffentlicht am

1 Einleitung
Niederwaldbewirtschaftung ist eine traditionelle Form der Waldnutzung, bei der alle 20 bis 30 Jahre kleinflächige Holzeinschläge durchgeführt werden. In Mitteleuropa wurden Niederwälder weitgehend von Hochwäldern abgelöst, aber besonders in Süd- und Südosteuropa werden Wälder noch auf diese Art bewirtschaftet. In Südtirol (Norditalien) wird in den submediterranen Laubmischwäldern der kollinen Höhenstufe mit Flaumeiche (Quercus pubescens), Hopfenbuche (Ostrya carpinifolia), Edelkastanie (Castanea sativa) und anderen Gehölzen (vgl. Peer 1995) auch heute noch Niederwaldwirtschaft praktiziert. Sie dient hier vor allem dem Schutz vor Steinschlägen, aber auch der Brennholzproduktion (Autonome Provinz Bozen-Südtirol 2007).
Im Vergleich zur Hochwaldwirtschaft stellt die Niederwaldbewirtschaftung eine relativ intensive Form der Waldnutzung mit starken Eingriffen in kurzen Zeitabständen dar. Forstwirtschaftliche Eingriffe sind ökologisch betrachtet Störungsereignisse und können die Invasion der Waldbestände durch nicht-einheimische Arten begünstigen (Böhmer et al. 2001). Biologische Invasionen werden als eine der wichtigsten Ursachen für den weltweiten Biodiversitätsverlust angesehen und haben z.T. erhebliche Veränderungen der Ökosystemstruktur und -funktionen zur Folge (Higgins et al. 1999). Deshalb ist das Zurückdrängen invasiver Arten ein wichtiges Teilziel der europäischen Biodiversitätsstrategie (Europäische Kommission 2011). Um dieses umzusetzen, sind ökologische Grundlagenuntersuchungen notwendig, die das Invasionspotenzial der örtlich relevanten Neophyten und deren Konkurrenzkraft im Vergleich zu den einheimischen Arten beleuchten.
Mittlerweile sind die beiden nicht-einheimischen Baumarten Götterbaum (Ailanthus altissima aus Ost-Asien) und Robinie (Robinia pseudoacacia aus den Appalachen, Nord-Amerika) europaweit in zahlreiche Ökosysteme vorgedrungen und stellen somit invasive Arten dar. In Waldgebieten sind insbesondere lichte Wälder, Kahlschlagflächen und Waldränder betroffen (Kohler 1963; Kowarik 2010). Vor diesem Hintergrund geht der Beitrag der Frage nach, ob die Niederwaldbewirtschaftung mit ihren relativ häufigen Kahlhieben die Einwanderung dieser nicht-einheimischen Baumarten fördert. Während die Ökologie dieser beiden Baumarten in urban-industriellen Ökosystemen gut dokumentiert ist (Basnou 2009, Kowarik & Säumel 2007), gibt es bisher vergleichsweise wenige Untersuchungen in Waldökosystemen und zu den Wechselwirkungen mit anderen Baumarten. Insbesondere die Auswirkungen der Waldbewirtschaftung auf die Ausbreitung und Etablierung der beiden invasiven Arten wurde bislang kaum beleuchtet (Kowarik 2010). Unklar ist auch, wie sich das Invasionspotenzial der beiden Baumarten in Niederwäldern verhält (Liess & Drescher 2008).
Diese Studie beleuchtet deshalb folgende Fragen:
1. Sind die beiden nicht-einheimischen Baumarten in jüngeren Waldbeständen stärker vertreten als in älteren? Zeigen Altbäume und Verjüngung dabei dasselbe Muster?
2. Welche Standortfaktoren begünstigen die Verjüngung der beiden nicht-einheimischen Arten?
3. Verjüngen sich Robinie und Götterbaum besser in den Niederwäldern als einheimische Baumarten? Wachsen sie zudem schneller?
2 Methoden
2.1 Untersuchungsgebiet
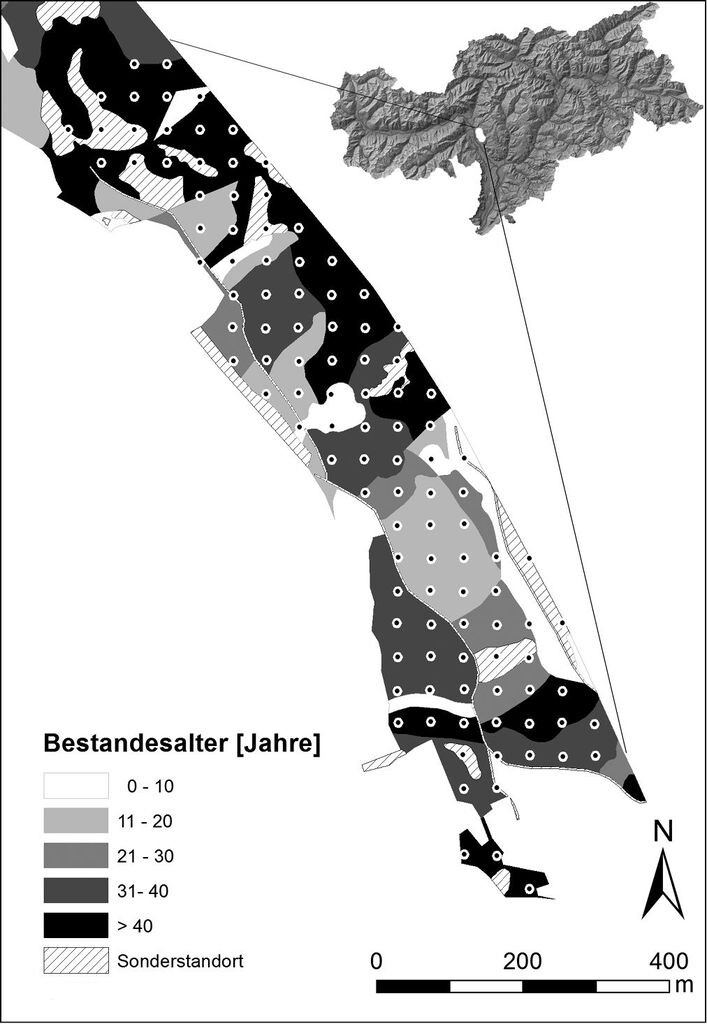
Die Untersuchungen wurden im oberen Etschtal in Südtirol (Norditalien), südlich der Stadt Meran (46°34'50''N, 11°12'46''E), durchgeführt (Abb. 1). Der untersuchte Niederwald befindet sich in einer Höhenlage zwischen 260 und 430 m ü.d.M. Das Klima ist submediterran mit mittleren Jahresniederschlägen von ca. 700 mm. Die mittlere Jahrestemperatur beträgt 11,5 °C mit absoluten sommerlichen Höchstwerten von 40 °C. Das Ausgangsgestein bildet vulkanischer Quarzporphyr, über dem sich hauptsächlich basenarme Syroseme und Ranker gebildet haben (Autonome Provinz Bozen-Südtirol 2007).
Im untersuchten Waldgebiet herrscht natürlicherweise Hopfenbuchen-Mannaeschenwald vor (Fraxino orni-Ostryetum quercetosum pubescentis). In sehr flachgründigen und sonnenexponierten Bereichen wachsen thermophile Flaumeichenwälder (Antherico liliaginis-Quercetum pubescentis; Peer 1995). Neben Robinie (Robinia pseudoacacia) und Götterbaum (Ailanthus altissima) kommen im Untersuchungsgebiet die einheimischen Baumarten Feldahorn (Acer campestre), Zürgelbaum (Celtis australis), Vogelkirsche (Prunus avium), Weichselkirsche (Prunus mahaleb), Traubeneiche (Quercus petraea), Speierling (Sorbus domestica), Elsbeere (Sorbus torminalis), Winterlinde (Tilia cordata), Bergulme (Ulmus glabra) und Feldulme (Ulmus minor) vor.
Die Bewirtschaftung des Niederwaldes erfolgt mit einer festgelegten Umtriebszeit von 30 Jahren (Autonome Provinz Bozen-Südtirol 2007), jedoch blieben viele Bestände in der Vergangenheit ungenutzt und sind heute im Sinne des Niederwaldes überaltert. Die Einschläge erfolgen in schachbrettartiger Verteilung über die Fläche. Vereinzelt wurden Überhälter auf den Schlagflächen belassen.
2.2 Untersuchungsdesign
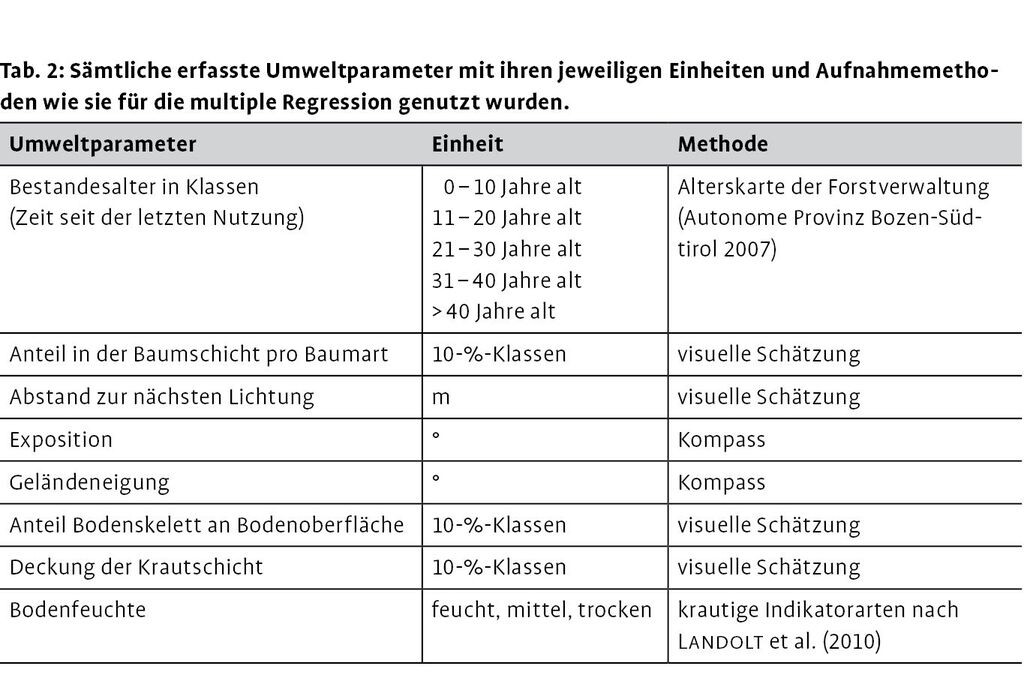
Wegen der kleinräumigen Verteilung von Beständen verschiedenen Alters und des ausgeprägten Mikroreliefs im Gargazoner Gemeindewald wurde ein regelmäßiges Punktegitter über das 25,5 ha große Untersuchungsgebiet gelegt. Bei einem Abstand von 45 m ergaben sich 126 mögliche Aufnahmepunkte, wovon 13 nicht zugänglich waren oder auf einem Weg lagen. Auf den verbleibenden 113 Punkten wurde die Verjüngung sämtlicher Baumarten in einem Radius von 1,5 m (Probekreisfläche ca. 7 m²) gezählt. Die krautigen Zeigerarten für die Einschätzung der Bodenfeuchte wurden im Radius von 2,0 m aufgenommen. Beide Radien wurden mit der Hangneigung korreliert. Die Aufnahmen fanden im Mai und Juni 2011 statt (Radtke et al. 2013).
Die Verjüngung wurde ab einer Höhe von 10 cm und bis zu einem Brusthöhendurchmesser (BHD) von 7 cm getrennt nach Höhenklassen (11-33, 34-66, 67-99, 100-199, > 199 cm) aufgenommen. Zwischen generativen und vegetativen Sprossen fand keine Unterscheidung statt, weil das Ausgraben aus dem skelettreichen Untergrund zu zeitaufwendig gewesen wäre.
Die Bodenfeuchte wurde über 14 festgelegte krautige Indikatorarten abgeleitet, von denen jeweils sieben Arten Zeigerwerte für trockene oder feuchtere Bedingungen aufwiesen (Tab. 1). Aufnahmepunkte wurden als feucht oder trocken klassifiziert, wenn ausschließlich Arten der jeweiligen Gruppe vorkamen. Waren auf einem Aufnahmepunkt Arten beider Gruppen vorhanden, wurde der Punkt als mittel im Feuchte-Trockenheitsgradienten bewertet.
Das Bestandesalter, d.h. die Zeit seit dem letzten Kahlhieb, eingeteilt in Zehn-Jahresstufen (0-10, 11-20, 21-30, 31-40, > 40 Jahre), ließ sich aus der Altersklassenkarte der Forstverwaltung entnehmen (Abb. 1). Die Deckung der Baumschicht wurde visuell und nach Arten getrennt in einem Radius von 20 m um den Aufnahmepunkt in 10-%-Klassen geschätzt. Zusätzlich wurden Einzelindividuen von Robinie und Götterbaum mit einem BHD > 7 cm als potenzielle Mutterbäume erfasst. Als Einflussfaktor für die Lichtverfügbarkeit wurde die Entfernung eines Aufnahmepunkts zur nächsten Lichtung visuell geschätzt. Auf Freiflächen liegende Aufnahmepunkte wurden mit 0 m Abstand registriert. War keine Lichtung in Sichtweite, wurde ein Maximalwert von 50 m angegeben. Der Anteil des Bodenskeletts (sichtbare Steine) an der Bodenoberfläche und der Deckungsgrad der Krautschicht wurden in 10-%-Klassen geschätzt.
2.3 Statistische Auswertung
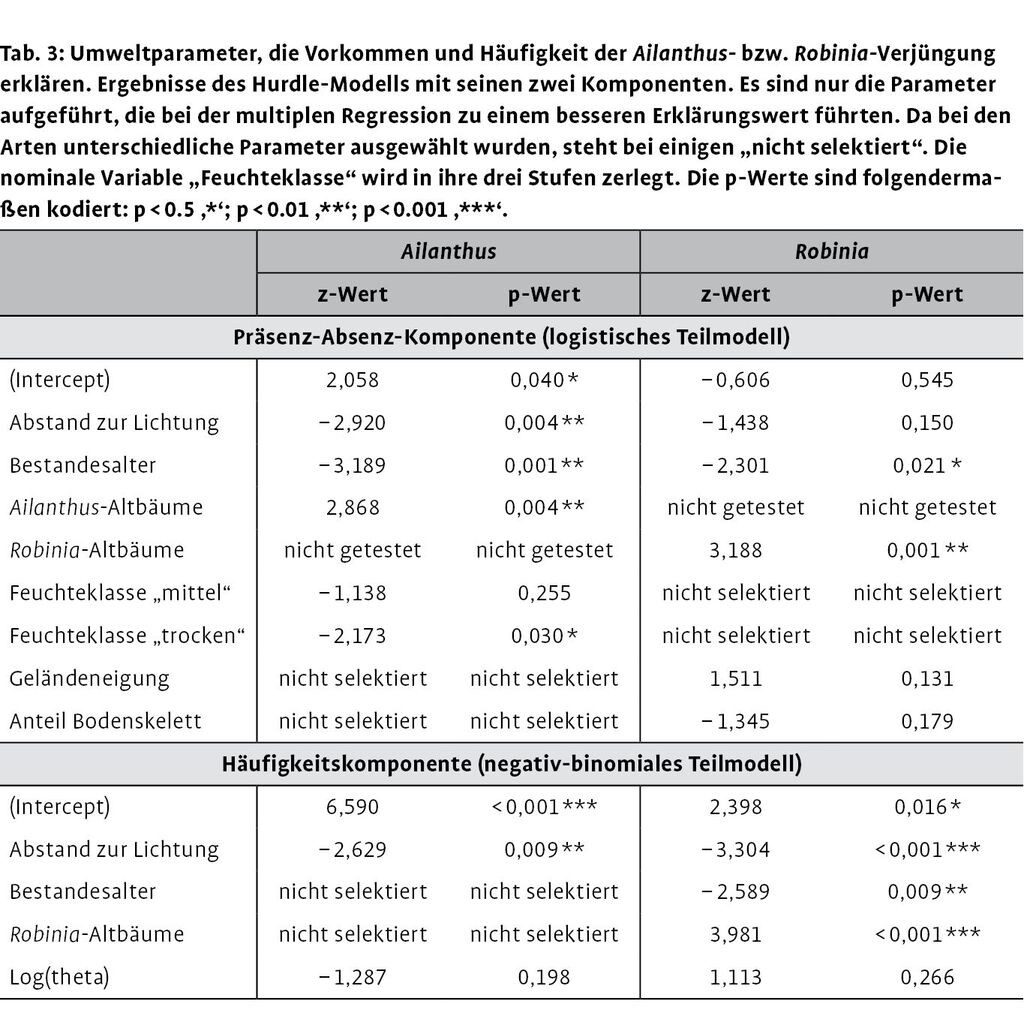
Alle statistischen Analysen wurden mit der freien Software R durchgeführt (R Development Core Team 2011). Verjüngungsaufnahmen enthalten typischerweise viele Nullen, z.B. dann, wenn Arten auf einem Aufnahmepunkt fehlen (Zhang et al. 2012). Um diese zero-inflated count distribution (eine mit Nullen aufgeblasene Zählverteilung) statistisch zu berücksichtigen, haben wir ein Hurdle-Modell aus dem pscl package (Zeileis et al. 2008) angewandt. Dieses Modell besteht aus zwei Komponenten mit einem logistischen Teil für die Präsenz-Absenz-Daten und einem negativ-binomialen Teil für die Zähldaten (Häufigkeiten). Die negative Binomialverteilung ist für Baumverjüngungsdatensätze geeignet (Zhang et al. 2012). Um zu untersuchen, welche der acht aufgenommenen Parameter das Vorkommen von Robinien- und Götterbaumverjüngung bzw. deren Häufigkeiten am besten erklären, wurde eine multiple Regression durchgeführt (Tab. 2). Da für das Hurdle-Modell noch keine automatische Variablenauswahl (step Funktion) verfügbar ist, wurden für jede Art je zwei generalized linear models gerechnet: ein logistisches für die Präsenz-Absenz-Daten und ein negativ-binomiales für die Häufigkeitsdaten. So konnten die eingebauten Funktionen step und stepAIC genutzt werden. Die Modelle für die Häufigkeitsdaten wurden mit den im logistischen Modell ausgewählten Variablen gestartet, da acht Variablen für die relativ geringe Anzahl an Aufnahmepunkten mit Verjüngung zu viel waren (Ailanthus= 26; Robinia= 20). Die auf diese Weise ausgewählten Variablen wurden dann für die Berechnung des Hurdle-Modells verwendet (Tab. 3).
Das Verhältnis nicht-einheimischer zu einheimischer Verjüngung in den verschiedenen Altersklassen wurde mittels eines nicht-parametrischen Kruskal-Wallis-Tests verglichen.
3 Ergebnisse
3.1 Einfluss des Bestandesalters
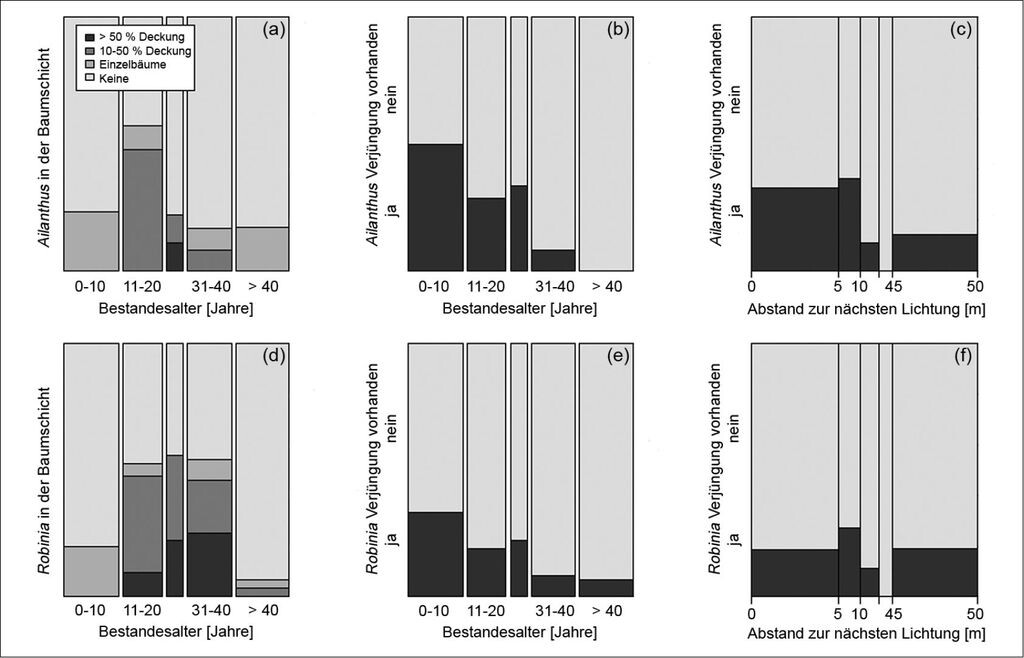
Im Gargazoner Gemeindewald war die Etablierung der Robinie deutlich weiter fortgeschritten als jene des Götterbaums. Ihr Anteil an der Baumschicht betrug 14 % und der des Götterbaums nur 3 %. Die Robinie war vor allem im südlichen Teil des Untersuchungsgebiets und in Beständen mit einem Alter zwischen 31 und 40 Jahren bestandsbildend (Abb. 2d). Der Götterbaum kam auf der gesamten Fläche vor, allerdings häufig nur als Einzelbaum und in jüngeren Beständen (Abb. 2a). Götterbaumverjüngung wurde auf 26 von 113 Aufnahmepunkten und Robinienverjüngung auf 20 Punkten gefunden. 9 % aller Jungpflanzen waren Götterbäume und nur 2 % Robinien. Die fortschreitende Invasion in Zeit und Raum wird durch den treppenförmigen Anstieg in der Verjüngungsabbildung deutlich, d.h. in jüngeren Beständen wurde häufiger Neophytenverjüngung gefunden als in älteren (Abb. 2b, e). Dieses Muster war beim Götterbaum deutlicher (Stufen sind steiler) als bei der Robinie (Stufen sind flacher).
3.2 Einfluss der Standortfaktoren
Das Vorhandensein von Götterbaumverjüngung wurde am besten durch die folgenden Parameter erklärt: Bestandesalter, Entfernung zur nächsten Lichtung, Vorhandensein von Mutterbäumen des Götterbaums und Bodenfeuchte (Tab. 3). Die Häufigkeit der Götterbaumjungpflanzen und die Vorkommenswahrscheinlichkeit stiegen mit abnehmendem Abstand zu einer Lichtung (Abb. 2c). Götterbaumverjüngung kam hauptsächlich in unter 30 Jahre alten Beständen vor (Abb. 2b). Mutterbäume in der Umgebung erhöhten die Vorkommenswahrscheinlichkeit für Götterbaumverjüngung deutlich. Bezüglich der Feuchteklassen war Götterbaumverjüngung häufiger auf feuchten und mittleren Standorten als auf trockenen Standorten zu finden (Tab. 3; feucht: 16 von 55 Punkten mit Götterbaumverjüngung, mittel: 1 von 10 und trocken: 6 von 25).
Vorkommen und Häufigkeit der Robinienverjüngung nahmen mit dem Bestandesalter ab und stiegen beim Vorhandensein von Mutterbäumen (Tab. 3, Abb. 2e). Das Vorkommen der Robinienverjüngung korrelierte zwar nicht mit dem Abstand zur Lichtung (Abb. 2f), aber ihre Häufigkeit nahm mit der Nähe zur Lichtung zu (Tab. 3). Jungpflanzen der Robinie kamen häufiger auf feuchten und mittleren Flächen vor als auf trockenen (feucht: 12 von 55 mit Robinienverjüngung, mittel: 3 von 10 und trocken: 3 von 25).
Zusammenfassend kann festgehalten werden, dass sowohl das Vorkommen als auch die Häufigkeit der Neophytenverjüngung positiv mit dem Vorhandensein von Mutterbäumen (der jeweiligen Art) und negativ mit dem Bestandesalter korrelieren.
3.3 Verhältnis von einheimischer zu nicht-einheimischer Verjüngung
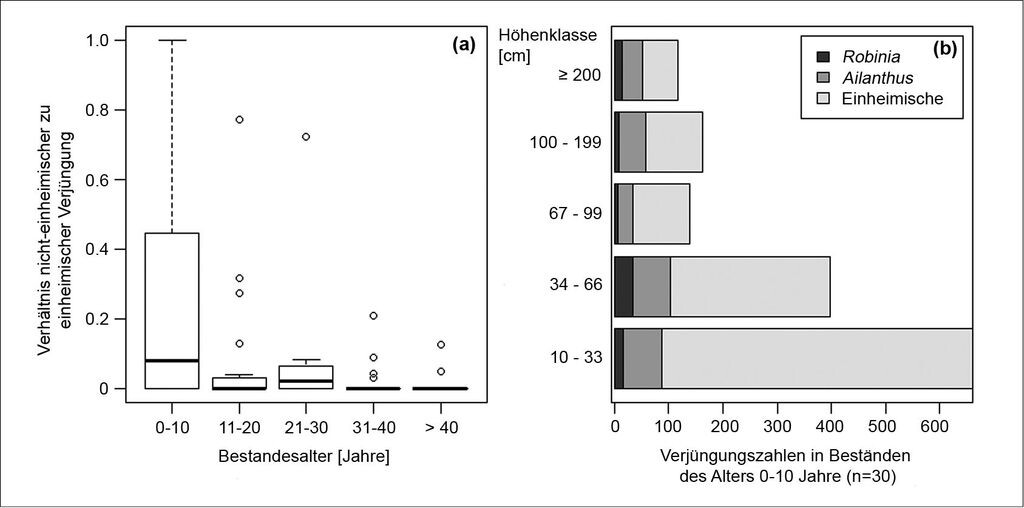
Der Anteil von Neophyten war in den jüngsten Beständen am höchsten (Abb. 3a). Eine genauere Betrachtung dieser Flächen (n = 30) zeigte, dass die Anteile der einheimischen Arten von der kleinsten zur größten Höhenklasse hin abnahmen (Abb. 3b).
4 Diskussion
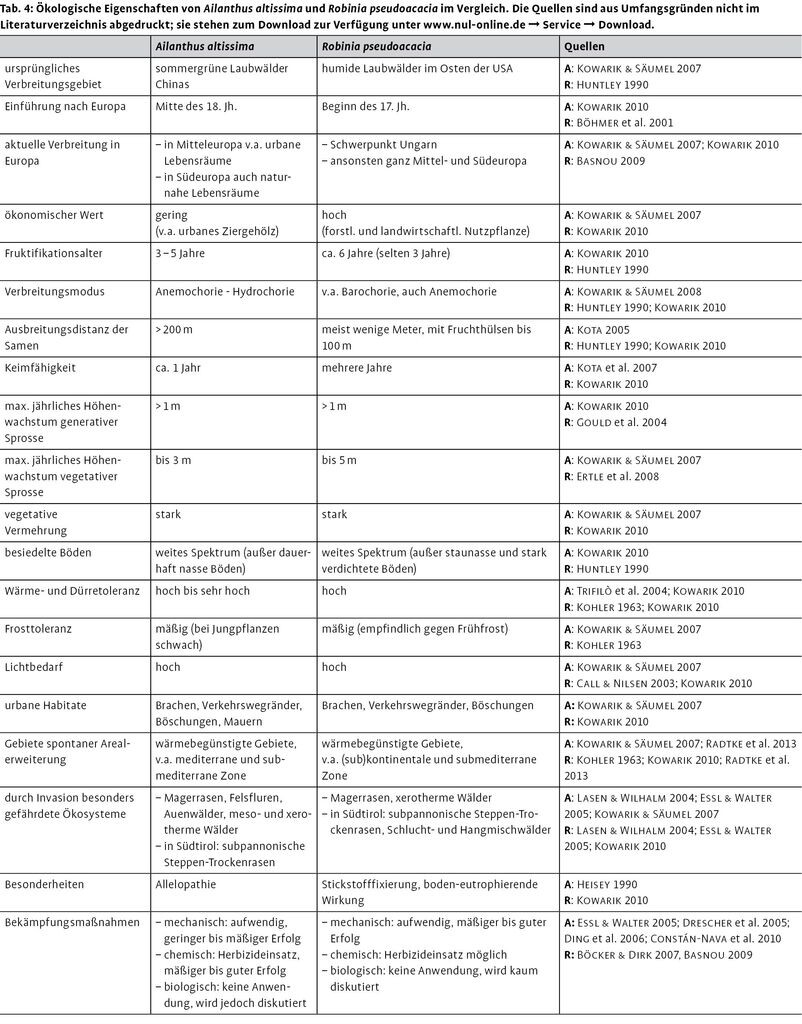
Zur besseren Interpretation der Ergebnisse stellt Tab. 4 die ökologischen Eigenschaften von A. altissima und R. pseudoacacia einander gegenüber.
Beide Arten sind typische Pionierbaumarten, d.h. ihre Diasporen verbreiten sich weit und ihre Sämlinge wachsen schnell. Im untersuchten Niederwald ist mit einem stetigen Sameneintrag zu rechnen, weil er an Siedlungsflächen mit zahlreichen Altbäumen angrenzt. Die Samen finden auf frisch geschlagenen Kahlflächen mit hohem Lichtangebot und geringer Konkurrenz einheimischer Arten sehr gute Keimungs- und Wachstumsbedingungen. So können sich die Jungpflanzen schnell im Kronenraum etablieren. Wegen der relativ kurzen Einschlagzyklen im Niederwald ist die Dauer der sukzessiven Entwicklung für die einheimischen Arten zu kurz, um relevante Konkurrenzwirkungen auf die lichtbedürftigen Neophyten auszuüben (z.B. Ausdunkeln) und diese auf lange Sicht zu verdrängen, wie es bei längeren Umtriebszeiten beobachtet wurde (Call & Nilsen 2003).
Daher wäre es sinnvoll, vorhandene Niederwälder langfristig in Hochwälder ohne Kahlhiebe zu überführen. Dieses stellt eine effektive Methode dar, um den Einfluss unerwünschter Arten zu regulieren (Ammer et al. 2011). Allerdings könnte dieses die Schutzfunktion gegen Steinschlag und die Vielfalt der Baumarten verringern. Eine weitere Möglichkeit wäre, die Umtriebszeiten so zu verlängern, dass einheimische Arten die konkurrierenden Neophyten zumindest teilweise ausdunkeln können. Alternativ könnte man die Bewirtschaftung stärker in Richtung Mittelwaldwirtschaft lenken, indem mehr Überhälter auf den Schlagflächen belassen werden, um den bodennahen Lichteinfall und damit den Konkurrenzvorteil von Götterbaum und Robinie einzugrenzen.
Die Robinie ist im Gargazoner Gemeindewald schon deutlich stärker etabliert als der Götterbaum. Sie ist in der Baumschicht generell häufiger und kommt auch in bis zu 40 Jahre alten Beständen vor. Die meisten Götterbäume befinden sich hingegen in bis zu 20 Jahre alten Beständen. Die Einwanderung des Götterbaums scheint sich also nach dem gleichen Muster wie die der Robinie zu wiederholen nur 20 Jahre später. Nach der Besiedlung konkurrenzfreier, sonniger Kahlflächen wächst der Götterbaum in die Baumschicht ein. Mit der zunehmenden Anzahl von Mutterbäumen steigen die Samenproduktion und die Konzentration allelopathischer Substanzen im Boden (Heisey 1990), so dass sich der Götterbaum zukünftig noch stärker ausbreiten könnte als die Robinie.
Zusammenfassend können die Niederwaldhiebe also als Störungsereignisse gesehen werden, die für Robinie und Götterbaum Habitate mit einer sehr günstigen Ressourcenversorgung und geringer intraspezifischer Konkurrenz schaffen. Auf ähnliche Weise wanderten beide Arten nach Waldbränden in Wälder der Südschweiz ein (Maringer et al. 2012). Störungsereignisse gelten generell als invasionsfördernd (Catford et al. 2012).
Die Ergebnisse unterstreichen die hohe Lichtbedürftigkeit der Keimlinge und Jungpflanzen des Götterbaums (Kowarik 1995). Im Gegensatz zur Götterbaumverjüngung war aber das Vorhandensein der Robinienverjüngung nicht mit der Entfernung zur Lichtung korreliert. Wahrscheinlich waren viele der aufgenommenen Robinienjungpflanzen vegetative Sprosse, die im Vergleich zu aus Samen entstandenen Pflanzen deutlich schattenverträglicher sind (Kowarik 1996). Die Robinie pflanzt sich in Waldökosystemen überwiegend vegetativ fort (Böhmer et al. 2001). Das dürfte auch im Untersuchungsgebiet der Fall sein, da Robinienverjüngung nur auf sieben Punkten ohne Mutterbäume gefunden wurde. Der Götterbaum reagiert bei einer Auflichtung der Bestände mit einer ähnlichen Strategie: Die im Vergleich zu Sämlingen schattentoleranteren, vegetativen Nachkommen etablieren sich ausgehend von der Rametenbank rasch im Kronenraum (Kowarik 1995).
Generell gelten Götterbaum und Robinie als trockenresistent und kommen auf verschiedensten Trockenstandorten wie z.B. in den typischerweise trockenen Flaumeichenwäldern vor (Kohler, 1963, Liess & Drescher 2008, Walter et al. 2005). Gerade die Jungpflanzen des Götterbaums können wegen ihres ausgeprägten Wurzelsystems auch unter äußerst trockenen Bedingungen überleben (Trifilò et al. 2004). Im Untersuchungsgebiet wurden beide Arten sowohl auf trockenen als auch auf feuchten Standorten gefunden, jedoch zeigte sich nur beim Götterbaum, dass er signifikant seltener auf trockenen Flächen vorkam. Dennoch konnte beobachtet werden, dass er sich teils auch auf Geröllhalden, also Primärstandorten, wo sich aus oberen Hanglagen gelöste Felsen abgelagert haben, etablieren konnte. Somit ist damit zu rechnen, dass beide Arten unter günstigen Lichtbedingungen imstande sind, sich über das gesamte Untersuchungsgebiet auszubreiten. Zur Überprüfung, ob es für beide Arten eine physiologische Trockengrenze in den thermophilen Eichenwäldern gibt oder inwieweit ihr Wachstumspotenzial unter den dortigen Bedingungen eingeschränkt ist, müssten weitere, großflächigere Untersuchungen in den trockenen Gebieten durchgeführt werden.
Mit zunehmendem Bestandesalter verschob sich das Verhältnis von Neophyten zu einheimischen Arten in der Verjüngung zugunsten der einheimischen Arten. Betrachtet man nur die weniger als zehn Jahre alten Bestände, so besaßen die Neophyten in den oberen Höhenklassen größere Anteile als in den unteren Höhenklassen. Ähnlich wie heimische Pioniere scheinen sie also eine höhere Überlebensrate zu haben und/oder ein rascheres Jugendwachstum. Beides ist wahrscheinlich, da sowohl die vegetativen als auch die generativen Sprosse beider Arten ein großes Wuchspotenzial aufweisen (Tab. 4) und die Robinie darüber hinaus durch ihre Dornen vor Verbiss geschützt ist.
Zur Bekämpfung wurden in Gargazon auf einigen Flächen die ca. 1 m hohen Götterbäume abgeschlagen. Das regte jedoch die Bildung vegetativer Sprosse an, denn beide Arten verfügen über ein enormes Stockausschlagsvermögen (Kowarik 1996, Kowarik & Säumel 2007). Andere mechanische Regulierungsmaßnahmen wie das Ringeln der Stämme könnten bei beiden Arten zielführend sein (vgl. Tab. 4), sind jedoch enorm kosten- und zeitaufwendig (Annighöfer et al. 2012).
Dank
Wir bedanken uns für die Hilfe bei der statistischen Auswertung bei A. Terwei und Prof. A. Zeileis sowie bei Prof. G. Tonon und der Abteilung Forstwirtschaft der Provinz Bozen bei der Planung und Durchführung dieser Studie. Unser Dank gilt auch der Stemmler-Stiftung im Stifterverband für die Deutsche Wissenschaft und der Stiftung der Freien Universität Bozen für die Finanzierung.
Literatur
Ammer, C., Balandier, P., Bentsen, N., Coll, L., Löf, M. (2011): Forest vegetation management under debate: an introduction. Eur. J. Forest Res. 130, 1-5.
Annighöfer, P.J., Schall, P., Kawaletz, H., Mölder, I., Terwei, A., Zerbe, S., Ammer, C. (2012): Vegetative growth response of black cherry (Prunus serotina, Ehrh.) to different mechanical control methods in a biosphere reserve. Canadian J. For. Res. 42 (12), 2037-2051.
Autonome Provinz Bozen-Südtirol (2007). Gemeinde Gargazon, Behandlungsplan der Wald- und Weidegüter 2008 2017. 11 S.
Basnou, C. (2009): Robinia pseudoacacia L., black locust (Fabaceae, Magnoliophyta). Handbook of Alien Species in Europe. Springer, Dordrecht, Niederlande, p. 357.
Böhmer, H.-J., Heger, T., Trepl, L. (2001): Fallstudien zu gebietsfremden Arten in Deutschland. Texte des Umweltbundesamtes 13, Berlin.
Call, L.J., Nilsen, E.T. (2003): Analysis of spatial patterns and spatial association between the invasive Tree-of-Heaven (Ailanthus altissima) and the native Black Locust (Robinia pseudoacacia). The American Midland Naturalist 150 (1), 1-14.
Catford, J.A., Daehler, C.C., Murphy, H.T., Sheppard, A.W., Hardesty, B.D., Westcott, D.A., Rejmánek, M., Bellingham, P.J., Pergl, J., Horvitz, C.C., Hulme, P.E. (2012): The intermediate disturbance hypothesis and plant invasions: Implications for species richness and management. Perspectives in Plant Ecology, Evolution and Systematics 14 (3), 231-241.
Europäische Kommission (2011): Lebensversicherung und Naturkapital: Eine Biodiversitätsstrategie der EU für das Jahr 2020. Mitteilung der Kommission an das Europäische Parlament, den Rat, den Europäischen Wirtschafts- und Sozialausschuss und den Ausschuss der Regionen. Brüssel.
Heisey, R.M. (1990): Evidence for allelopathy by tree-of-heaven (Ailanthus altissima). J. Chem. Ecol. 16 (6), 2039-2055.
Higgins, S., Richardson, D., Cowling, R., Trinder-Smith, T. (1999): Predicting the landscape-scale distribution of alien plants and their threat to plant diversity. Conserv. Biol. 13, 303-313.
Kohler, A. (1963): Zum pflanzengeografischen Verhalten der Robinie in Deutschland. Beitr. Naturk. Forsch. SW-Deutschland 12 (1), 3-18.
Kowarik, I. (1995): Clonal growth in Ailanthus altissima on a natural site in West Virginia. Journal of Vegetation Science 6 (6), 853-856.
(1996). Funktionen klonalen Wachstums von Bäumen bei der Brachflächen-Sukzession unter besonderer Beachtung von Robinia pseudoacacia. Verh. Ges. Ökol. 26, 173-181.
(2010): Biologische Invasionen. Ulmer, Stuttgart, 2. Aufl.
, Säumel, I. (2007): Biological flora of Central Europe: Ailanthus altissima (Mill.) Swingle. Perspectives in Plant Ecology, Evolution and Systematics 8 (4), 207-237.
Landolt, E., Bäumler, B., Erhardt, A., Hegg, O., Klötzli, F., Lämmler, W., Nobis, M., Rudmann-Maurer, K., Schweingruber, F.H., Theurillat, J.-P., Urmi, E., Vust, M., Wohlgemuth, T. (2010): Flora indicativa. Ökologische Zeigerwerte und biologische Kennzeichen zur Flora der Schweiz und der Alpen. Haupt, Bern.
Lasen, C., Wilhalm, T. (2004): Natura 2000-Lebensräume in Südtirol. Auton. Prov. Bozen-Südtirol, Abt. Natur und Landschaft, Bozen.
Liess, N., Drescher, A. (2008). Ailanthus altissima spreading in the Danube National Park. In: Rabitsch, W., Essl, F., Klingenstein, F., Hrsg., Biological Invasions from Ecology to Conservation, Neobiota 7, 84-95.
Maringer, J., Wohlgemuth, T., Neff, C., Pezzatti, G.B., Conedera, M. (2012): Post-fire spread of alien plant species in a mixed broad-leaved forest of the Insubric region. Flora 207 (1), 19-29.
Peer, T. (1995): Die natürliche Pflanzendecke Südtirols. Begleittext und Karte der natürlichen Vegetation 1:200.000. Frangart, Südtirol.
R Development Core Team (2011): R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Wien, Österreich.
Trifilò, P., Raimondo, F., Nardini, A., Lo Gullo, M.A., Salleo, S. (2004): Drought resistance of Ailanthus altissima: root hydraulics and water relations. Tree Physiology 24 (1), 107-114.
Walter, J., Essl, F., Englisch, T., Kien, M. (2005): Neophytes in Austria: Habitat preferences and ecological effects. In: Nentwig, W., Bacher, S., Cock, M., Dietz, H.J., Gigon, A., Wittenberg, R., eds., Biological Invasions from Ecology to Control, Neobiota 6, 13-25.
Zeileis, A., Kleiber, C., Jackman, S. (2008): Regression Models for Count Data in R. Journal of Statistical Software 27 (8), 1-25.
Zhang, X., Lei, Y., Cai, D., Liu, F. (2012): Predicting tree recruitment with negative binomial mixture models. Forest Ecology and Management 270, 209-215.
Anschriften der Verfasser(innen): Forstreferendar Stefan Ambraß, Hessen-Forst, Forstamt Wetzlar, Hörnsheimer Eck 11a, D-35578 Wetzlar, E-Mail stefan.ambrass@googlemail.com; Anna Radtke und Prof. Dr. Stefan Zerbe, Fakultät für Naturwissenschaften und Technik, Freie Universität Bozen, Universitätsplatz 5, I-39100 Bozen, E-Mail anna.radtke@natec.unibz.it bzw. stefan.zerbe@unibz.it; Veronika Fontana, Institut für Ökologie, Leopold-Franzens Universität, Sternwartestraße 15, A-6020 Innsbruck, E-Mail Veronika.Fontana@student.uibk.ac.at; Prof. Dr. Christian Ammer, Abteilung Waldbau und Waldökologie der gemäßigten Zonen, Georg-August-Universität, Büsgenweg 1, D-37077 Göttingen, E-Mail Christian.Ammer@forst.uni-goettingen.de.
-
Ausbreitung und Management vonGötterbaum und Robinie in Niederwäldern -
Ausbreitung und Management vonGötterbaum und Robinie in Niederwäldern -
Ausbreitung und Management vonGötterbaum und Robinie in Niederwäldern -
Ausbreitung und Management vonGötterbaum und Robinie in Niederwäldern -
Abb. 1: Netz der Aufnahmepunkte im Untersuchungsgebiet mit dem Bestandesalter in Klassen laut vorliegender Forstkarte (Autonome Provinz Bozen-Südtirol 2007). Oben rechts Lage des Untersuchungsgebietes innerhalb von Südtirol (grau), Italien. -
Abb. 2: Häufigkeit von Ailanthus (a-c) und Robinia (d-f) in der Baumschicht (a, d) und in der Verjüngung (b, e) über das Bestandesalter (a, b, d, e) und im Verhältnis zum Abstand zur nächsten Lichtung (c, f). Je breiter eine Klasse auf der x-Achse, desto mehr Aufnahmepunkte sind darin enthalten. -
Abb. 3: (a) Verhältnis nicht-einheimischer zu einheimischer Verjüngung in den verschiedenen Altersklassen als Boxplot-Darstellung (Median mit dem 25- bzw. 75-%-Quantil, außerhalb liegende Werte sind als Kreise dargestellt). (b) Kumulative Verjüngungszahlen aller 010 Jahre alten Aufnahmepunkte nach Höhenklassen und Art. -
Ausbreitung und Management vonGötterbaum und Robinie in Niederwäldern

Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.