Veränderungen von Tagfaltergemeinschaften
Abstracts
In den Mittelgebirgsregionen Märkisches Sauerland und Westliches Weserbergland in Nordrhein-Westfalen wurde entlang von Transekten ein Tagfalter-Monitoring durchgeführt. Die daraus resultierenden Abundanzdaten wurden Trendanalysen unterzogen. Zudem wurde der Einfluss von Temperatur und Niederschlag auf die Abundanzdynamik untersucht. Die Ergebnisse wurden mit einer Studie für das gesamte Bundesland Nordrhein-Westfalen verglichen, in der die Auswirkungen des Klimawandels auf die Tagfalterfauna abgeschätzt werden. Für die Region Westliches Weserbergland liegen darüber hinaus Rasterfrequenzdaten zur Häufigkeit von Tagfaltern und Zygaeniden seit 1981 vor. Die statistischen Analysen auf Basis der Transektdaten ergaben, dass zwischen der Einschätzung für ganz NRW und den Ergebnissen der vorliegenden Studie hinsichtlich der Trendrichtungen eine hohe Übereinstimmung besteht. Dagegen konnten bei einigen Arten signifikante Trends mit in beiden Untersuchungsregionen unterschiedlichen Vorzeichen festgestellt werden. Bei vielen Arten bestehen zum Teil deutliche Zusammenhänge der Abundanzdynamik mit Temperatur und Niederschlag. Bei einigen Arten scheint die Abundanzdynamik jedoch nicht durch klimatische Einflüsse bestimmt zu werden, so dass hier neben anderen Ursachen Faktoren wie z.B. strukturelle Habitatverschlechterungen eine große Rolle spielen könnten. Die Auswertung der Rasterfrequenz-bezogenen Veränderungen zeigen, dass sich die Entwicklungen der Häufigkeiten in Abhängigkeit der Lebensraumpräferenzen der Arten stark unterscheiden. Arten, die geschlossene Wälder und Gebüsche mittlerer Standorte bevorzugen, nehmen im Verhältnis zu Arten der Magerrasen zu.
Changes of Butterfly Communities Influence of climatic factors on species abundance in the Westliches Weserbergland and in the Märkisches Sauerland
A butterfly monitoring was carried out in two low-mountain regions, Märkisches Sauerland and Westliches Weserbergland, in the federal state of North Rhine-Westphalia (NRW). The study analysed abundance data of the transect counts in order to detect significant trends and to reveal possible impacts of temperature and rainfall on the abundance dynamics of the butterfly species. The results were compared to a state-wide study on the implications of global warming on butterfly species. Additionally, raster data from the year 1981 onwards are available for the Westliches Weserbergland on the frequency of butterflies and burnet moths.
The analysis results of the transect data show a strong correspondance of the trends between the current study and the NRW-assessment. In a few cases, however, the slopes of the trends in abundance data differ between the two study regions. Abundance dynamics of many species show significant correlations to temperature and rainfall. A few species show no correlations of abundance dynamics and climatic variables. This indicates that other factors such as habitat deterioration may have had stronger effects on the abundances than climatic variables. The analysis of changes in grid frequencies reveals significant differences in the proportion of increasing and decreasing species in different habitats. The proportion of species inhabiting forests and woody habitats is significantly increasing compared to species of nutrient poor grasslands.
- Veröffentlicht am
1 Einleitung
1.1 Klimawandel und Umwelt
Der Klimawandel ist heute in weiten Teilen der Fachwelt und der Öffentlichkeit als akute Bedrohung für die Lebensgrundlagen des Menschen akzeptiert. In Europa wurde in den letzten 100 Jahren ein Anstieg der Jahresmitteltemperatur um 0,95 °C beobachtet (EEA 2004). Für Deutschland werden für die nächsten 50 Jahre eine weitere Temperaturzunahme vor allem im Winter, eine starke Zunahme der Winterniederschläge verbunden mit zunehmenden Abflussmengen und Hochwasserereignissen sowie ein Anstieg trocken-heißer Wetterlagen im Sommer bei gleichzeitiger Häufung von Starkniederschlagsereignissen prognostiziert (EEA 2004, Leuschner & Schipka 2004). Für Nordrhein-Westfalen wird bis 2060 ein mittlere Erwärmung von 1,9 °C und eine Zunahme der Jahresniederschlagssumme von 5 % angenommen (MUNLV NRW 2009).
Die vielfältigen Auswirkungen dieser Klimaänderungen machen auch nicht vor der belebten Umwelt halt. In diesem Kontext haben in den vergangenen Jahren zahlreiche Studien belegt, dass erhebliche Veränderungen bei Arten und Lebensgemeinschaften ablaufen werden bzw. bereits zu beobachten sind (Parmesan & Yohe 2003, Walther et al. 2002). Auf internationaler Ebene gibt es bereits eine Vielzahl von Arbeiten, die für Tagfalter klimainduzierte Areal- und Phänologieveränderungen beschreiben (z.B. Hill et al. 2002, Parmesan et al. 1999). Während hierbei in der Anfangsphase spektakuläre Neueinwanderungen bzw. Arealerweiterungen im Blickpunkt des Interesses lagen, mehren sich die Anzeichen, dass Areale zahlreicher Arten in erheblichem Ausmaß schrumpfen, manche Arten sogar ganz verschwinden werden (Settele et al. 2008, Thomas et al. 2006). Schätzungen gehen davon aus, dass weltweit je nach Szenario 15 bis 37 % der Tier- und Pflanzenarten aussterben könnten (Thomas et al. 2004).
1.2 Einschätzung klimainduzierter Veränderungen von Tagfalterzönosen
Es bestehen von europäischer bis zur regionalen Ebene verschiedene methodische Ansätze, klimainduzierte Veränderungen von Tagfaltergemeinschaften zu analysieren. So prognostizieren Settele et al. (2008) die Veränderungen in der Verbreitung europäischer Arten, indem sie ein statistisches Modell der aktuellen Klimanische der jeweiligen Art mit den vorausgesagten Klimaveränderungen verschneiden und daraus die künftige Verbreitung in Europa extrapolieren. Auf nationaler Ebene existieren z.B. für Großbritannien detaillierte Studien über statistische Zusammenhänge von Klimavariablen und der Abundanzdynamik von Tagfalterarten (Roy et al. 2001 u.a.). Für Deutschland fehlen bisher Einschätzungen zu klimainduzierten Veränderungen der Tagfaltergemeinschaften. Für das Bundesland Nordrhein-Westfalen existiert seit kurzem eine auf Experteneinschätzungen basierende Studie, in der die Klimasensitivität der hier vorkommenden Arten sowie klimainduzierte Zu- und Abnahmen auf Basis der beobachteten Veränderungen in den letzten zehn Jahren abgeschätzt werden (Behrens et al. 2009).
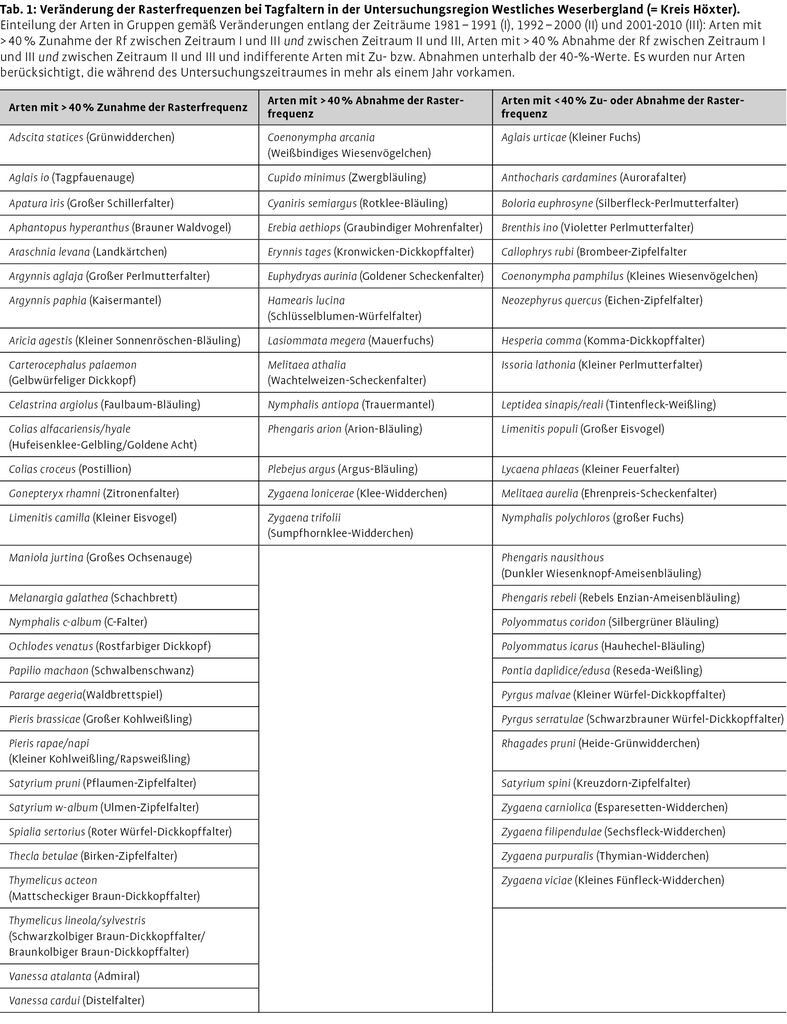
Auf regionaler Ebene finden sich einige Publikationen, in denen Abundanzentwicklungen der Tagfalterarten beschrieben werden, sei es auf der Basis von qualitativen Beobachtungsdaten oder auf der Grundlage von Transektzählungen. So berichten Brunzel et al. (2008) für das Märkische Sauerland (Südwestfalen) von drastischen Abnahmen z.B. des Hochmoor-Perlmutterfalters (Boloria aquilonaris) seit 1989 und starken Zunahmen bei Mauerfuchs (Lasiommata megera), Wald-Brettspiel (Pararge aegeria) sowie Schwefelvögelchen (Lycaena tityrus), ohne hierzu jedoch statistische Analysen durchzuführen (weitere deutsche Namen vgl. Tab. 1). Ähnliche Beobachtungen machen Laussmann et al. (2010) für das benachbarte Bergische Land im Raum Wuppertal, doch ist hier L. megera z.B. in den letzten Jahren stark zurückgegangen. Für Westfalen berichten Fartmann et al. (2002) über eine Ausbreitung von Aricia agestis, für Bayern publizieren Hermann & Bolz (2003) zur Arealausdehnung von Lycaena dispar.
1.3 Statistische Analysen von Tagfalter-Monitoringdaten und Experteneinschätzungen
Statistisch abgesicherte Trendanalysen sind nur möglich, wenn ein längeres, kontinuierliches Monitoring vorliegt, z.B. in Form von Transektzählungen. In Großbritannien konnte die Abundanzdynamik vieler Arten in einem bisher über 30-jährigen Monitoring mit klimatischen Veränderungen in statistischen Zusammenhang gebracht werden (Roy et al. 2001). Längerfristige Monitoring-Programme bei Tagfaltern existieren auch in den Niederlanden (vgl. Bos et al. 2006, van Swaay et al. 2008) und Finnland (vgl. Kuussaari et al. 2007). In Deutschland fehlen solche Langzeit-Datenreihen bisher: Ein Tagfalter-Monitoring in NRW hat 2001 begonnen, ein bundesweites im Jahre 2005. Auf regionaler Ebene besteht beispielsweise das Monitoring im Märkischen Sauerland, das seit dem Jahr 2000 kontinuierlich durchgeführt wird (vgl. Brunzel et al. 2008). Auch für das westliche Weserbergland (Kreis Höxter im östlichen NRW) besteht seit 2001 ein Monitoring der Individuenzahlen auf acht Transekten, zusätzlich existieren Rasterfeld-bezogene Erfassungen seit Anfang der 1980er Jahre. Statistische Analysen beider Datensätze fehlten bisher jedoch.
Statistische Analysen von Monitoring-Daten gerade bei einer gut untersuchten Gruppe wie den Tagfaltern können einen wichtigen Beitrag leisten, um möglichen Auswirkungen klimatischer Veränderungen auf die Biodiversität zu beschreiben. Insbesondere können derartige Auswertungen helfen, so genannte climatic impact indicators zu identifizieren (vgl. Dröschmeister 2009). In diesem Kontext versucht die vorliegende Studie, einen Beitrag zu leisten. Ausgangshypothese ist hierbei die für jede Tagfalterart Nordrhein-Westfalens durch Behrens et al. (2009) getroffene Einschätzung ihrer durch den Klimawandel induzierten Bestandsentwicklungen (positiv, negativ oder unverändert). Vor diesem Hintergrund sollen insbesondere folgende Fragen untersucht werden:
(1) Lassen sich aus den Datensätzen der Untersuchungsregionen im Märkischen Sauerland und im Weserbergland statistisch abgesicherte Trends für einzelne Arten herausarbeiten?
(2) Decken sich diese Trends mit der Gesamteinschätzung für NRW oder mit Trends aus anderen Bezugsräumen?
(3) Lassen sich für die Untersuchungsregionen Zusammenhänge zwischen Entwicklungen bei einzelnen Arten und klimatischen Parametern statistisch absichern?
(4) Unterscheiden sich Arten mit unterschiedlichen Entwicklungen hinsichtlich ihrer Lebensraumpräferenzen?
2 Methoden
2.1 Geländeerhebungen in der Region Märkisches Sauerland
In der Untersuchungsregion Märkisches Sauerland (MS) (= südlicher Märkischer Kreis in Südwestfalen) wurden ab 2000 an 23 Transekten jedes Jahr kontinuierlich Tagfalterabundanzen (= Individuenzahlen) erfasst (vgl. auch Brunzel et al. 2008). Die Transekte decken Lebensräume wie Pionierrasen in Steinbrüchen, magere Weiden, Borstgrasrasen, Feuchtwiesen und saure Seggen-Niedermoore, Laubwälder sowie Heiden und Übergangsmoore ab.
Jedes Transekt wurde an zwei bis vier Terminen pro Jahr aufgesucht, so dass möglichst viele der vorkommenden Arten innerhalb eines Jahres erfasst werden konnten. Die einzelnen Begehungstermine wurden so gewählt, dass Arten und Individuenzahlen an den Transekten jedes Jahr zum phänologisch gleichen Zeitpunkt aufgenommen werden konnten und so über die Jahre vergleichbar sind. Die Transektlänge variierte zwischen 200 m und 2 700 m.
Von den Wanderfaltern V. cardui und V. atalanta sowie von Arten, die zu Beginn des Monitorings sehr häufig waren, liegen erst seit 2005 Daten zu den Individuenzahlen vor, so dass sie bei den Analysen ausgeklammert werden mussten. Da die Untersuchungsintensität die Ergebnisse von Monitoring-Programmen stark beeinflusst und in anderen Studien Datensätze mit wenigen Beobachtungen weggelassen wurden (z.B. Kuussaari et al. 2007), wurden nur diejenigen 22 Arten berücksichtigt, die ausreichend häufig waren, um eine verlässliche Datengrundlage zu bieten. Das bedeutet, dass Arten, die während des Monitorings nie mehr als dreimal pro Jahr beobachtet wurden, von allen weiteren statistischen Analysen ausgeschlossen wurden.
Die Summe der Individuen jeder der 22 Arten wurde auf die Transektlänge und die Anzahl der Zähltermine standardisiert. Dabei wurden nur die maximalen Individuenzahlen jeder Art pro Jahr berücksichtigt. Alle Transektzählungen wurden darüber hinaus dann auf die Länge des kürzesten Transekts standardisiert (200 m). Deshalb verstehen sich alle Abundanzen als maximale Anzahl der Individuen je 200 m Transektlänge und Besuch. Um die Gesamtdynamik über die Jahre zu analysieren, wurden die Abundanzen aller Transekte der Untersuchungsregion aufsummiert.
2.2 Geländeerhebungen in der Region Westliches Weserbergland
In der Untersuchungsregion Westliches Weserbergland (WB) (= Kreis Höxter in Ostwestfalen) wurde seit 2001 ebenfalls ein Tagfalter-Monitoring durchgeführt. Neben den Tagfaltern wurden hier die Rotwidderchen (Gattung Zygaena) einbezogen, so dass 48 Arten in die Analysen einflossen. Die Transekte befinden sich in Kalk-Magerrasen mit anschließenden Gebüschen und Feldgehölzen. Im Unterschied zur Untersuchungsregion Märkisches Sauerland wurde hier jedes Transekt einmal wöchentlich zwischen Ende März/Anfang April und Anfang bis Mitte Oktober begangen. Die Transektlängen schwanken zwischen 200 m und 1400 m. Auch hier wurden die maximalen Individuenzahlen jeder Art auf 200 m Transektlänge und die Anzahl der Transektbegehungen standardisiert. Zur Analyse der Gesamtdynamik über die Jahre wurden die Abundanzen aller acht Transekte wiederum aufsummiert. Die absoluten Abundanzen lassen sich jedoch nicht zwischen den Untersuchungsregionen vergleichen, da es sich nicht um ähnliche Lebensräume handelt.
Zusätzlich bestehen in der Untersuchungsregion Weserbergland Rasterfrequenzdaten für die Zeiträume 1981 bis 1991, 1992 bis 2000 und von 2001 bis 2010 jährlich. Diese geben unter der Voraussetzung einer vergleichbaren Erfassungsintensität für jeden Zeitraum Auskunft über den Anteil der von der jeweiligen Art besetzten Rasterfelder an der Gesamtzahl der untersuchten Rasterfelder. Da die Erfassungsintensität in den Zeiträumen unterschiedlich war, wurde die Rasterfrequenz (%) für weitere Analysen auf die Anzahl der Datensätze pro Zeitschnitt (= Gesamtzahl aller Falterbeobachtungen pro Zeitraum) standardisiert. Die Rasterfelder (auf MTB-Viertel-Quadrantenbasis) sind über die gesamte Untersuchungsregion Westliches Weserbergland (= Kreis Höxter) verteilt. Bei der Analyse der Rasterfrequenzen blieben Arten, die nur in einem Jahr der zu Grunde liegenden Zeiträume beobachtet wurden, unberücksichtigt. Insgesamt konnten so von 71 Arten (inkl. Widderchen) die Veränderungen der Rasterfrequenzen analysiert werden (Tab. 1), wozu jede dieser Arten zusätzlich ihrem bevorzugten Lebensraum (Mehrfachnennungen möglich) als Imago (nach Ebert & Rennwald 1991 und Ebert 1994) zugeordnet wurde.
2.3 Klimatische Parameter als erklärende Variablen
Um Zusammenhänge zwischen der Dynamik der Tagfalterabundanzen und klimatischen Parametern zu untersuchen, wurden monatliche Daten zu Temperatur und Niederschlag der Wetterstation Lüdenscheid (für die Untersuchungsregion MS) und der Wetterstation Warburg (für die Untersuchungsregion WB) von Oktober 1999 bis Dezember 2010 aufbereitet. Hierzu wurden die Daten nach Sommer-Halbjahr (April bis September) und Winter-Halbjahr (Oktober bis März) aufgeteilt und die Temperaturmittel (TS und TW: Temperaturmittel Sommer und Winter) und Mittel der Niederschlagssummen (NiS und NiW: Niederschlagssumme Sommer und Niederschlagssumme Winter) des jeweiligen Zeitraumes gebildet. Hierbei ist zu beachten, dass das Winter-Halbjahr eines Jahres die drei letzten Monate des vorangegangenen Jahres mit einbezieht. Da Abundanzdaten von Arten mit nur einer Generation von der Häufigkeit im vorangegangenen Jahr abhängen, wurden in gleicher Weise die Temperaturmittel und Niederschlagssummen für das vorangegangene Jahr gebildet (TS-1 und TW-1, NiS-1 und NiW-1). Insgesamt flossen so acht Klimaparameter als erklärende Variablen der Abundanzdynamik in die multiplen Regressionsanalysen ein.
2.4 Statistik
In einem ersten Schritt ist zunächst geklärt worden, ob sich hinter der Abundanzdynamik über die Jahre tatsächlich ein klimatisches Signal verbirgt, oder ob die vorgefundenen signifikanten Korrelationen rein zufällige Ergebnisse sind. Hierzu wurden Korrelationsmatrizes für die jeweiligen Untersuchungsregionen angelegt, so dass für das Märkische Sauerland acht Klimavariablen gegen 22 Arten und für das westliche Weserbergland gegen 48 Arten gerechnet wurden. Geht man nun von einem Ergebnis aus, das keine klimatischen Ursachen hat, sondern rein zufallsbedingt ist, so müssten sich für MS auf einem 5%igen Signifikanzniveau rein rechnerisch 8,8 (5 % von 8 Variablen x 22 Arten) und für WB 19,2 statistisch signifikante Korrelationen ergeben. Da sich in der Analyse aber für MS 11 und für WB 24 statistisch signifikante Korrelationen (p ≤ 0,05) ergaben, weist dies auf einen statistisch signifikanten Einfluss klimatischer Parameter auf die Abundanzdynamik der untersuchten Arten hin.
In einem zweiten Schritt wurden für jede Art Regressionen gegen die Zeit gerechnet, um zu ermitteln, ob in der Abundanzdynamik ein signifikant negativer oder positiver Trend über die Jahre besteht. Es werden sowohl Signifikanzen auf dem üblichen 5-%-Niveau (p ≤ 0,05) als auch auf einem 10-%-Niveau angegeben. Da die Abundanzen eines Jahres aber von denjenigen des vorangegangenen Jahres abhängen können, wurde die jährliche Abfolge der Abundanzen auf zeitliche Autokorrelationen getestet. Es konnten in beiden Untersuchungsregionen nur bei drei Arten signifikante (p ≤ 0,05) zeitliche Autokorrelationen der Abundanzen mit den Abundanzen des vorangegangenen Jahres festgestellt werden (vgl. Tab. 2 unter http://www.nul-online-de Service Download).
In einem letzten Schritt wurden multiple Regressionsanalysen der abhängigen Variablen Abundanzdynamik mit den erklärenden acht Klimavariablen durchgeführt. Als Verfahren wurde stepwise forward gewählt. Nur das Modell mit der höchsten Varianzerklärung wurde berücksichtigt. Bei den zeitlich autokorrelierten Abundanzen wurden die Ergebnisse zusätzlich mit einem sequenziellen Bonferroni-Test abgesichert.
Zur Analyse der Veränderung der Rasterfrequenzen (Rf) wurden die Arten gemäß ihrer Häufigkeitsveränderungen in den Zeiträumen 1981-1991 (Zeitraum I), 1992-2000 (II) und 2001-2010 (III) in Gruppen eingeteilt: Arten mit > 40 % Zunahme der Rf zwischen Zeitraum I und III und zwischen Zeitraum II und III, Arten mit > 40 % Abnahme der Rf zwischen Zeitraum I und III und zwischen Zeitraum II und III und indifferente Arten mit Zu- bzw. Abnahmen unterhalb der 40-%-Werte (Tab. 2, Download). Diese Gruppeneinteilung bildet dann die Eingangshypothese für die statistischen Analysen. Die Entwicklung der Rasterfrequenzen in den verschieden Gruppen wurden mittels einer ANOVA mit Messwiederholung und Tukey-HSD als post-hoc-Test analysiert. Unterschiede im Verhältnis der Artengruppen zwischen den Lebensraumpräferenzen wurden dann durch Chi²-Tests ermittelt. Alle statistischen Tests wurden mit Statistica 7.0 und R 2.7.0 durchgeführt.
3 Ergebnisse
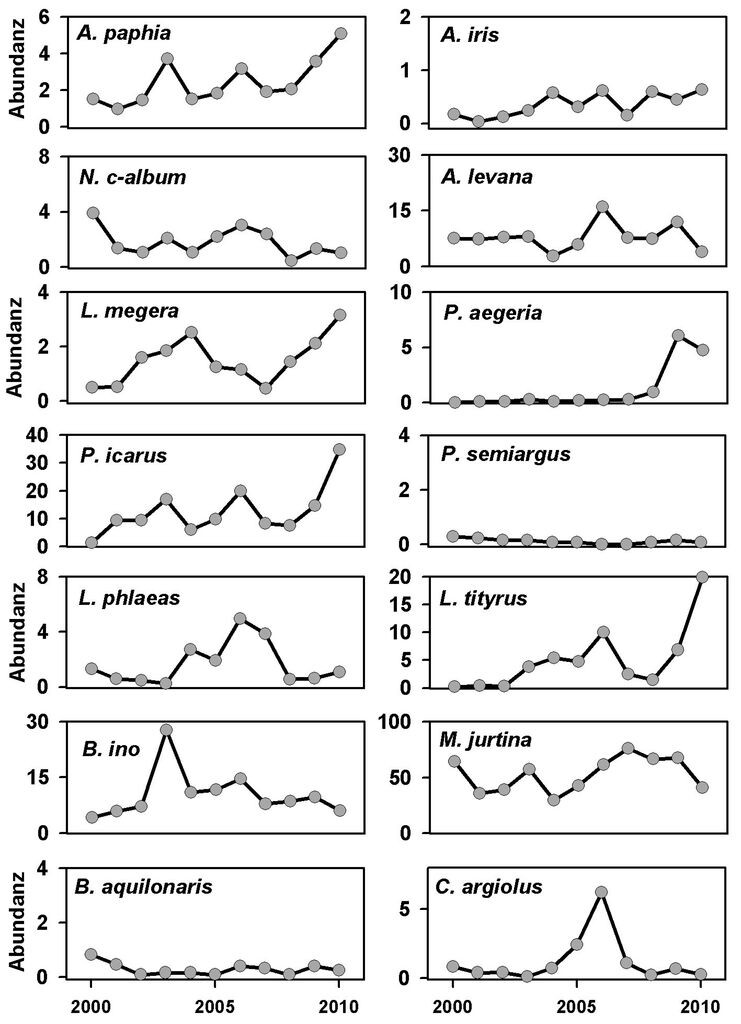
Die untersuchten Arten zeigen beträchtliche Unterschiede hinsichtlich ihrer Abundanzdynamik (Abb. 1a: MS, Abb. 1b: WB). Im Märkischen Sauerland sind auf Basis der Transektdaten seit 2000 zwei Arten auf 5-%-Niveau (p ≤ 0,05) signifikant zurückgegangen (P. semiargus, S. w-album, vgl. Tab. 2, Download), andere Arten sind ebenfalls rückläufig, allerdings nicht signifikant (z.B. B. aquilonaris). Dagegen zeigten sechs Arten signifikante Zunahmen (z.B. A. paphia, A. iris, L. tityrus, S. pruni). Bei zehn Arten (z.B. B. ino, N. c-album, A. levana, C. argiolus, L. phlaeas) ergaben sich aufgrund der beträchtlichen Abundanzschwankungen keine signifikanten Trends (vgl. Abb. 1a und Tab. 2, Download).
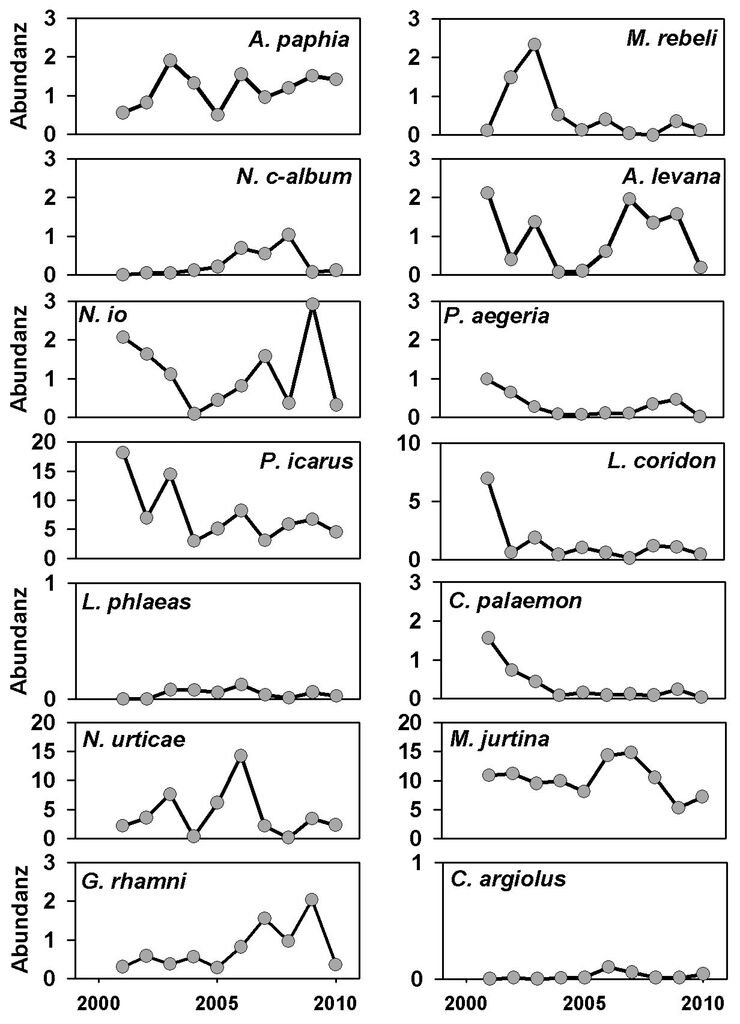
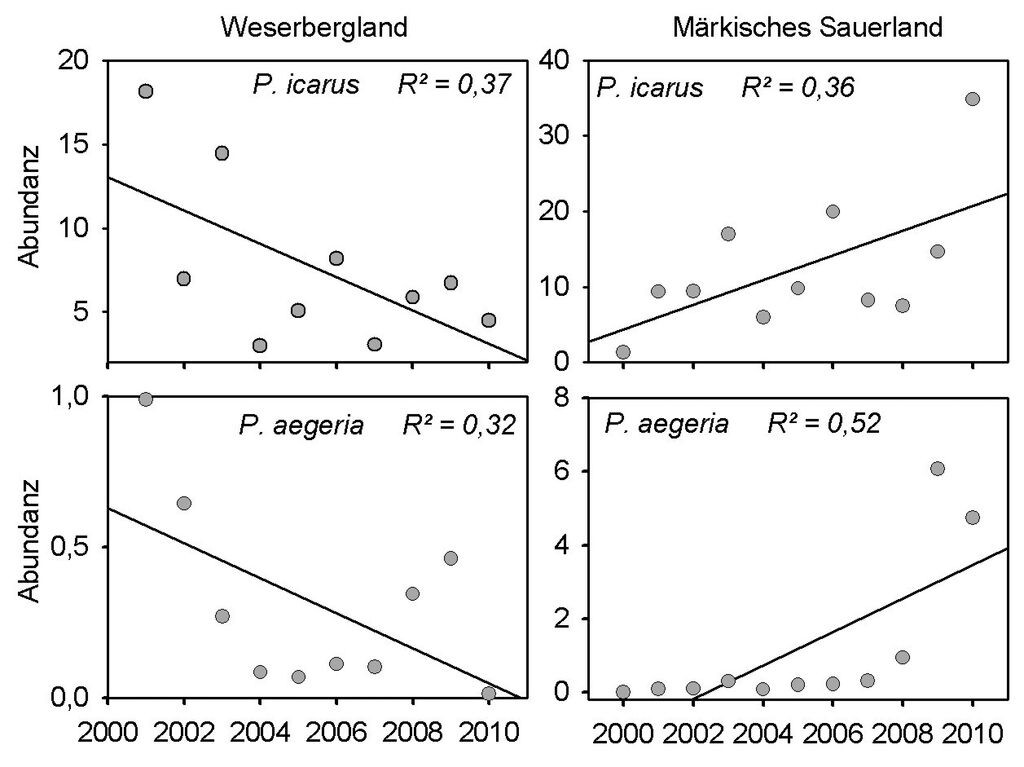
Dagegen bot die Analyse der Transektdaten auf den Kalkmagerrasen in der Region Westliches Weserbergland ein anderes Bild. Hier zeigten acht Arten auf 5-%-Niveau (p ≤ 0,05) signifikante Trends, von diesen waren sechs negativ (C. palaemon, A. cardamines, C. minimus, M. aurelia, C. arcania und Z. purpuralis). Auf 10-%-Niveau wiesen sogar 16 Arten signifikante Trends auf, von denen 14 negativ waren (vgl. Tab. 2, Download). Lediglich L. sinapis und L. camilla nahmen signifikant zu, wobei die jährlichen Abundanzen bei Letzterem jedoch zeitlich autokorreliert sind (vgl. Tab. 2, Download). Auch der Kreuzenzian-Ameisenbläuling (Phengaris rebeli) scheint abzunehmen (vgl. Abb. 1b), doch war der Trend nicht signifikant. Wie bei dieser Art zeigten auch viele andere Arten im WB aufgrund ihrer großen Abundanzschwankungen keine signifikanten Trends. Abb. 2 belegt am Beispiel von P. icarus und P. aegeria, dass einige Arten signifikante Trends aufwiesen, die in den beiden Untersuchungsregionen jedoch in unterschiedliche Richtungen verlaufen.
Nicht bei allen Arten mit signifikanten Trends in den Abundanzveränderungen konnten auch signifikante Korrelationen mit Klimavariablen festgestellt werden. Umgekehrt konnten bei einigen Arten signifikante Korrelationen mit Klimavariablen gefunden werden, obwohl keine signifikanten Zu- oder Abnahmen vorhanden waren. In diesen Fällen werden die Abundanzen zwar von Klimavariablen beeinflusst, der Einfluss manifestiert sich bis jetzt jedoch offenbar nicht in einem Trend. Auch konnten bei einigen Arten keine Übereinstimmungen zwischen den beiden Untersuchungsregionen hinsichtlich der Korrelationen zwischen Abundanzen und Klimavariablen gefunden werden (vgl. Tab. 2, Download: C. crocea, A. levana, L. phlaeas).
Andere Befunde ergeben jedoch eindeutigere Muster: Nicht unerwartet korrelieren die Abundanzen mehrerer Arten wie C. hylae/alfcariensis, N. c-album, A. levana und M. jurtina positiv mit der Sommertemperatur: Je höher diese war, desto mehr Individuen dieser Arten konnten gezählt werden. Überraschend sind jedoch bei relativ vielen Arten negative Korrelationen der Abundanzen mit den Sommertemperaturen (L. coridon, T. betulae, B. euphrosyne, A. cardamines, N. urticae, T. acteon, E. tages). Je höher in diesem Fall also die Temperaturen von April bis September waren, desto seltener wurden diese Arten entlang der Transekte in den Kalk-Magerrasen beobachtet.
Bei der Mehrheit der Arten zeigen die Abundanzen signifikant negative Korrelationen mit den Niederschlagssummen im gleichen Sommer und vorangegangenen Winter, unter ihnen z.B. C. pamphilus, A. agestis, P. semiargus und P. minimus. Je höher also die Niederschläge waren, desto geringere Abundanzen konnten festgestellt werden. Vor allem bei den Dickkopffaltern fiel der Einfluss der Niederschlagsummen auf. Dabei standen sechs negativen Korrelationen zwischen Abundanzen und Niederschlägen lediglich zwei positive gegenüber (vgl. Tab. 2, Download). Wie bei den univoltinen (= nur eine Generation/Jahr) Zygaeniden werden bei den überwiegend univoltinen Dickkopffalterarten die Abundanzen negativ von den Niederschlagsummen des Vorjahres beeinflusst.
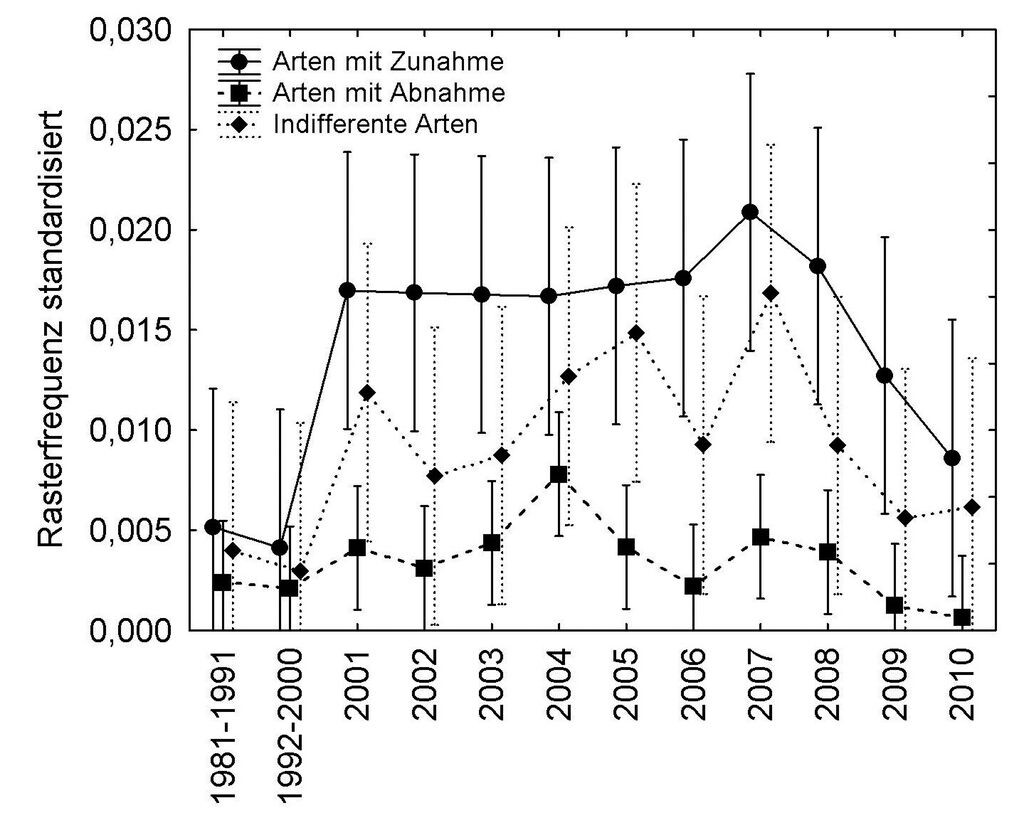
Die Analyse der Veränderungen auf Basis der Rasterfrequenzen ergab, dass lediglich der Trend für die Gruppe der zunehmenden Arten statistisch signifikant war (Abb. 3). Die negative Entwicklung der Artengruppe mit mehr als 40 % Abnahme der Rasterfrequenz (vgl. Tab. 1) war dagegen nicht signifikant.
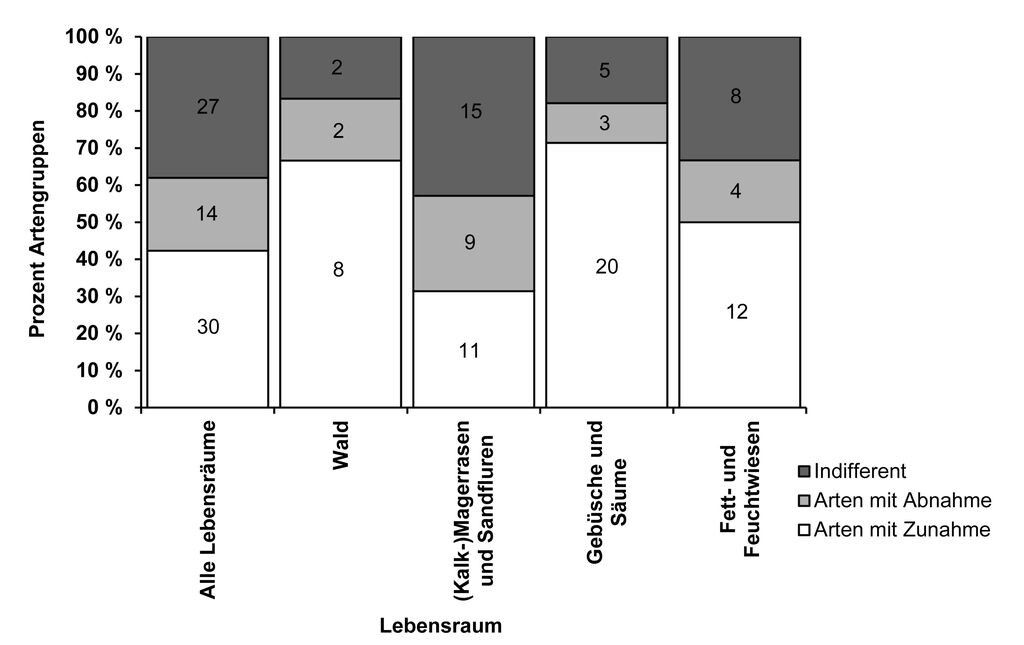
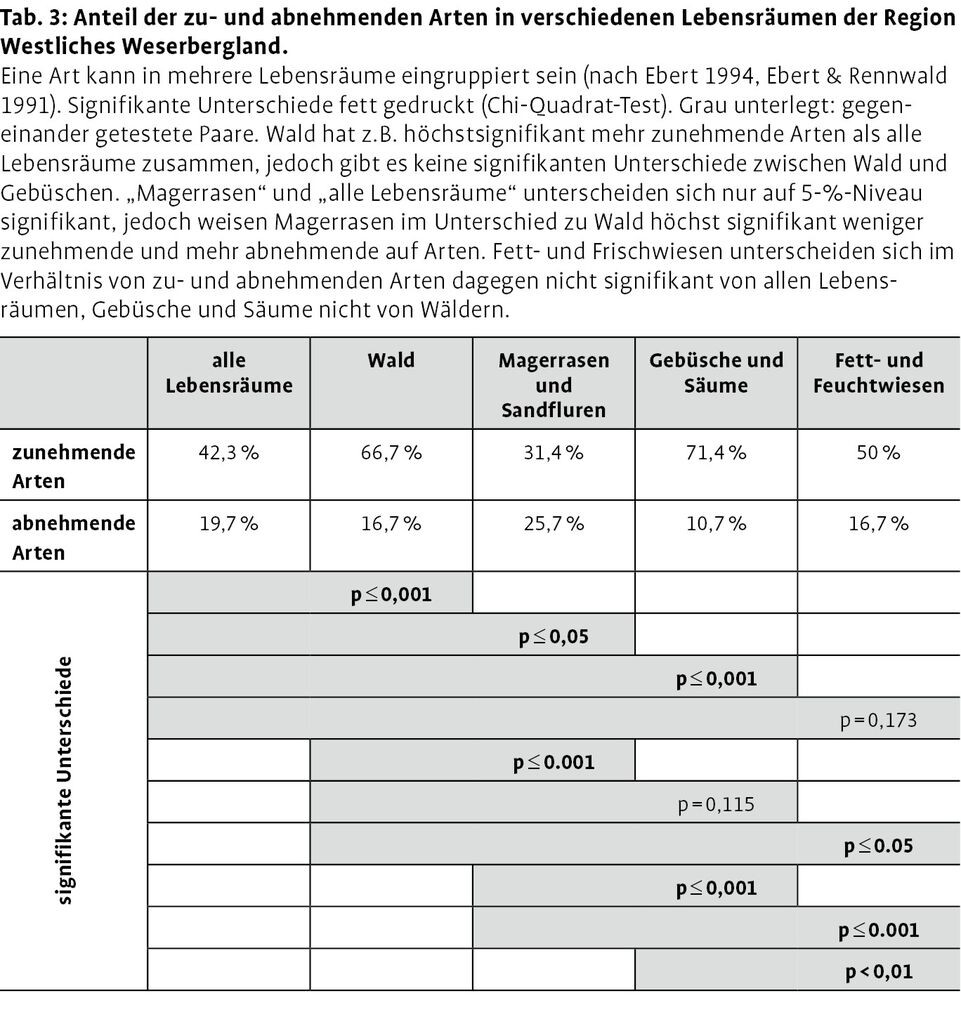
Betrachtet man allerdings die Verteilung der Arten mit Zu- und Abnahme gemäß ihrer Lebensraumpräferenzen, so ergeben sich deutliche Unterschiede (Abb. 4): Arten mit mehr als 40 % Zunahme finden sich im prozentualen Verhältnis signifikant häufiger in den Lebensräumen Wald, Gebüsche und Säume sowie Fett- und Feuchtwiesen als im Lebensraum Magerrasen (Tab. 3). Umgekehrt ist das prozentuale Verhältnis bei den Magerrasen bewohnenden Arten signifikant in Richtung der Gruppe mit mehr als 40 % Abnahme verschoben. Weiterhin wichen alle einzelnen Lebensraumtypen mit Ausnahme der Fett- und Feuchtwiesen signifikant von der prozentualen Verteilung der Artengruppen über alle Lebensräume ab (vgl. Tab. 3).
4 Diskussion
Von 48 Arten, für die Behrens et al. (2009) aufgrund ausreichender Datenlage eine Einschätzung ihrer Klimasensitivität vornehmen konnten (vgl. Tab. 2, Download), wurden 8,3 % eine negative Entwicklung und 41,6 % eine positive Entwicklung zugeschrieben. 51,1 % wurden als indifferent eingestuft. Die vorliegende Studie weist dagegen für die Untersuchungsregionen WB und MS deutlich mehr Arten mit negativem (35,4 %) und weniger Arten (20,8 %) mit positivem Trend aus. Diese Unterschiede zwischen beiden Studien beruhen vermutlich darauf, dass Behrens et al. (2009) explizit nur durch den Klimawandel induzierte Veränderungen bewerten, in der vorliegenden Studie signifikant negative Trends aber auch durch strukturelle Habitatverschlechterungen verursacht werden können.
Andere Gründe für leichte Unterschiede zwischen beiden Studien sind vermutlich in der Kürze des Monitoring-Zeitraumes zu sehen. So reicht wahrscheinlich die Datenreihe noch nicht aus, um beispielsweise den positiven Trend bei V. atalanta auch statistisch zu belegen. Dies gilt wohl auch für den negativen Trend bei B. aquilonaris (vgl. Brunzel et al. 2008). Starke Widersprüche, wie z.B. unterschiedliche Trendvorzeichen, finden sich dagegen zwischen beiden Studien kaum. So zeigen sieben von neun Arten mit positiver Trendeinschätzung bei Behrens et al. (2009) ebenfalls signifikant positive Trends in den Untersuchungsregionen, nur bei E. tages und M. aurelia finden sich unterschiedliche Trendrichtungen (vgl. Tab. 2, Download). Umgekehrt besteht nur bei A. iris eine Abweichung dahingehend, dass dieser Art bei Behrens et al. (2009) ein negativer Trend zugeschrieben wird, die statistischen Analysen ihr für die Untersuchungsregion MS jedoch einen positiven Trend bescheinigen. In der Region WB kam sie für Analysen nicht in ausreichender Zahl vor.
Bei fünf der 17 Arten mit signifikant negativem Trend (N. quercus, C. rubi, M. aurelia, C. arcania und Z. purpuralis) war in den Untersuchungsregionen kein signifikanter Einfluss klimatischer Variablen feststellbar (vgl. Tab. 2, Download). In diesen Fällen dürfte der negative Trend nicht auf klimatische Einflüsse zurückzuführen sein, sondern vermutlich eher auf strukturelle Habitatverschlechterungen (Sukzession o. Ä.), wobei hierfür im Einzelfall weitergehende Untersuchungen nötig wären. In anderen Fällen überlagern sich vermutlich unterschiedliche Ursachen. So zeigt S. w-album entlang der Transekte in der Untersuchungsregion MS einen signifikant negativen Trend, gleichzeitig korrelieren die Abundanzen negativ mit den Niederschlagssummen des vorangegangenen Sommers (vgl. Tab. 2, Download). Dieser Befund bedarf sicherlich einer vorsichtigen Interpretation. Zum einen ist die Antreffwahrscheinlichkeit von Imagines dieser Art stark von zufälligen Ereignissen beeinflusst, da sie zumeist im Wipfelbereich der Bäume fliegen. Zum anderen werden klimatische Einflüsse auf die Abundanz der Art sicherlich von der Abnahme blühfähiger Bergulmenbestände infolge des Ulmensterbens überlagert.
Es ist jedoch auch möglich, dass fehlende Korrelationen der Abundanzen mit klimatischen Variablen rein statistische Ursachen haben, wie z.B. eine zu geringe Datenbasis. So sind fehlende Zusammenhänge zwischen klimatischen Variablen und signifikanten Zunahmen bei L. sinapis/reali und S. pruni sicher darauf zurückzuführen, dass beide Arten erst seit wenigen Jahren überhaupt an den Transekten der beiden Untersuchungsregionen nachgewiesen wurden. Hinsichtlich der signifikanten Zunahme von A. paphia ist diese Deutung jedoch unwahrscheinlich, da die Art seit Beginn des Monitorings in ausreichender Zahl erfasst wurde. Hier könnten strukturelle Habitatverbesserungen, z.B. durch Sturmschäden oder forstliche Maßnahmen, eine gewisse Rolle spielen.
Eine sehr detaillierte Studie zu den statistischen Zusammenhängen von klimatischen Variablen und der Abundanzdynamik bei Tagfaltern haben Roy et al. (2001) publiziert. Auf der Grundlage von Transektzählungen des British Monitoring Scheme (BMS) von 1976 bis 1997 wurde hier der Einfluss von Temperatur und Niederschlag auf Monatsbasis untersucht. Auch in dieser Studie ergibt sich trotz des langen Monitoring-Zeitraumes ein heterogenes Bild der statistischen Zusammenhänge. Aufgrund der unterschiedlichen methodischen Ansätze lassen sich die Ergebnisse nicht mit der vorliegenden Untersuchung vergleichen. Die wenigen deutlichen Muster zeigen jedoch in ähnlichem Ausmaß Übereinstimmungen wie Abweichungen zwischen beiden Studien.
Auf Basis der Transektzählungen sind bei dem überwiegenden Teil der Arten mit statistisch absicherbaren Trends signifikante Zusammenhänge zwischen Klimavariablen und Abundanzdynamik nachweisbar. Vor dem Hintergrund der prognostizierten Auswirkungen des Klimawandels auf die Tagfaltergemeinschaften wird häufig diskutiert, inwieweit sich die Reaktionen von Arten in verschiedenen Lebensräumen unterscheiden und ob bestimmte Lebensräume mögliche Auswirkungen klimatischer Veränderungen eher kompensieren können (vgl. Braschler & Hill 2007, Franco et al. 2006, Oliver et al. 2009, Thomas et al. 2006). In dieser Hinsicht kann die Lebensraumpräferenz der auf Rasterfrequenzbasis zu- und abnehmenden Arten interessante Hinweise liefern. Der überproportional hohe Anteil der zunehmenden Arten in Wald- und Gebüschlebensräumen könnte im Hinblick auf eine Kompensation klimatischer Veränderungen durch einen Habitatwechsel einiger Tagfalterarten gedeutet werden (vgl. auch Pöyry et al. 2009, Thomas et al. 2001). Auch deckt sich der überproportional hohe Anteil stark abnehmender Arten mit einer Lebensraumpräferenz in Magerrasen (vgl. Abb. 4 und Tab. 3) mit der Hypothese, dass Arten trockener Lebensräume stärker unter den prognostizierten klimatischen Veränderungen leiden könnten (vgl. Hill et al. 2002, Davies et al. 2006, Oliver et al. 2009).
Im Bergischen Land um Wuppertal, einem der Untersuchungsregion MS benachbarten Raum, werden ähnlich große Unterschiede in der Entwicklung der Häufigkeiten von Wald- und Offenlandarten beobachtet (Laussmann et al. 2010). Auch in Finnland zeigen Arten aus magerem Grünland und Wald(rand) bewohnende Arten gegensätzliche Trends (Kuussari et al. 2007). In diesen Zusammenhang könnten auch einige zwischen den beiden Untersuchungsregionen unterschiedlich verlaufende Trends eingeordnet werden. So zeigt z.B. P. icarus in den Kalkmagerrasen-Transekten der klimatisch begünstigten Untersuchungsregion WB negative Trends, während die Art in der regenreicheren und sommerkühleren Untersuchungsregion MS einen positiven Trend aufweist.
Die vorliegende Studie zeigt klar, dass in den untersuchten Regionen in NRW in den letzten zehn Jahren erhebliche Veränderungen der Abundanzen von Tagfaltern feststellbar sind, die sich trotz der Kürze des Monitoring-Zeitraums zum Teil schon statistisch absichern lassen. Auch wenn nachgewiesen werden kann, dass hinter der Abundanzdynamik häufig ein klimatisches Signal steckt, wird deutlich, dass die Ergebnisse im Hinblick auf den Einfluss klimatischer Faktoren noch recht heterogen sind und andere Faktoren wie insbesondere strukturelle Habitatveränderungen ebenfalls eine große Rolle spielen.
Dank
Dies Studie wurde im Rahmen des DBU-Projekts Entwicklung von Anpassungsstrategien seitens des Naturschutzes zum Erhalt hochgradig durch den Klimawandel gefährdeter Lebensgemeinschaften (Az.: 25874 33/0) erarbeitet. Das Naturschutzzentrum Märkischer Kreis e.V. hat das Tagfalter-Monitoring im Märkischen Sauerland in früheren Jahren finanziert. Für wertvolle Kommentare zum Manuskript danken wir G. Hermann, Filderstadt. Den Herren H. Biermann, Bad Driburg, und A. Waldhoff, Steinheim, gebührt besonderer Dank für die Überlassung ihrer über viele Jahre erhobenen Daten. Weiter gebührt auch allen Falterfreunden Dank, die ihre Zeit geopfert haben, um die Datenbank für den Kreis Höxter mit aufzubauen.
Literatur
Behrens, M., Fartmann, T., Hölzel, N. (2009): Auswirkungen von Klimaänderungen auf die Biologische Vielfalt: Pilotstudie zu den voraussichtlichen Auswirkungen des Klimawandels auf ausgewählte Tier- und Pflanzenarten in Nordrhein-Westfalen. Teil 1: Fragestellung, Klimaszenario, erster Schritt der Empfindlichkeitsanalyse Kurzprognose. Institut für Landschaftsökologie (ILÖK); Westfälische Wilhelms-Universität Münster. Auftraggeber: Ministerium für Umwelt und Naturschutz, Landwirtschaft und Verbraucherschutz des Landes Nordrhein-Westfalen (MUNLV NRW).
Bos, F.G., Bosveld, M.A., Groenendijk, D.G., van Swaay, C.A.M., Wynhoff, I. (2006): De dagvlinders van Nederland. Verspreiding en bescherming. Nederlandse Fauna 7. Leiden, 381 S.
Braschler, B., Hill, J.K. (2007): Role of larval host plants in the climate-driven range expansion of the butterfly Polygonia c-album. J. Animal Ecology 76, 415-423.
Brunzel, S., Bussmann, M., Obergruber, H. (2008): Deutliche Veränderungen von Tagfalterzönosen als Folge von Ausbreitungsprozessen: Erste Ergebnisse eines Monitorings über 17 Jahre. Natur und Landschaft 83 (6), 280-287.
Davies, Z.G., Wilson, R.J., Coles, S., Thomas, C.D. (2006): Changing habitat associations of a thermally constrained species, the silver-spotted skipper butterfly, in response to climate warming. J. Animal Ecology 75, 247-256.
Dröschmeister, R. (2009): Climatic Impact Indicator ein Indikator der Auswirkungen des Klimawandels auf die biologische Vielfalt. In: Bundesamt für Naturschutz, Hrsg., Biodiversität und Klima Vernetzung der Akteure in Deutschland V Ergebnisse und Dokumentation des 5. Workshops, 32-34.
Ebert, G. (Hrsg., 1994): Die Schmetterlinge Baden-Württembergs, Bd. 3. Nachtfalter I. Ulmer, Stuttgart.
, Rennwald, E. (1991): Die Schmetterlinge Baden-Württembergs, Bd. 1. Tagfalter I. Ulmer, Stuttgart.
EEA European Environment Agency (2004): Impacts of Europes changing climate. An indicator-based assessment. EEA Report 2/04.
Fartmann, T., Dudler, H., Schulze, W. (2002): Zur Ausbreitung des Kleinen Sonnenröschen-Bläulings Aricia agestis ([Denis & Schiffermüller], 1775) in Westfalen (Lep., Lycaenidae) eine erste Übersicht. Mitt. Arb.-gem. Westfäl. Entomol. 18, 41-46.
Franco, A.A., Hill, J.K., Kitschke, C., Collingham, Y.C., Roy, D.B., Fox, R., Huntley, B. (2006): Impacts of climate warming and habitat loss on extinctions at species low-latitude range boundaries. Global Change Biology 12, 1545-1553.
Hermann, G., Bolz, R. (2003): Erster Nachweis des Großen Feuerfalters Lycaena dispar (Haworth, 1803) in Bayern mit Anmerkungen zu seiner Arealentwicklung in Süddeutschland (Insecta: Lepidoptera: Lycaenidae). Beitr. Bayer. Entomofaunistik 5, 17-23.
Leuschner, C., Schipka, M. (2004): Vorstudie Klimawandel und Naturschutz in Deutschland. BfN-Skripten 115, 35 S.
Hill, J.K., Thomas, C.D., Fox, R., Telfer, M.G., Willis, S.G., Asher, J., Huntley, B. (2002): Responses of butterflies to twentieth century climate warming: implications for future ranges. Proceedings of the Royal Society of London Series B Biological Sciences 269, 2163-2171.
Kuussaari, M., Heliölä, J, Pöyry, J., Saarinen, K. (2007): Contrasting trends of butterfly species preferring semi-natural grasslands, field margins and forest edges in northern Europe. J. Insect Conserv. 11, 351-366.
Laussmann, T., Radtke, A., Wiemert, T., Dahl, A. (2010): 150 Jahre Schmetterlingsbeobachtung in Wuppertal Auswirkungen von Klima- und Landschaftsveränderungen (Lepiodoptera). Entomolog. Zeitschr. 120, 269-277.
MUNLV, Ministerium für Umwelt und Naturschutz, Landwirtschaft und Verbraucherschutz des Landes Nordrhein-Westfalen (2009): Anpassung an den Klimawandel. Eine Strategie für Nordrhein-Westfalen. Düsseldorf, 166 S. ( http://www.umwelt.nrw.de/umwelt/klimawandel/index.php, 04.05.2009).
Oliver, T., Hill, J.K., Thomas, C.D., Brereton, T., Roy, D.B. (2009): Changes in habitat specificity of species at their climatic range boundaries. Ecology Letters 12, 1091-1102.
Parmesan, C., Rhyrholm, N., Stefanescu, C., Hill, J.K., Thomas, C.D, Descimons, H., Huntley, B., Kaila, B., Kullberg, J., Tammaru, T., Tennent, W.J., Thomas, J.A., Warren, M. (1999): Polewards shifts in geographical ranges of butterfly species associated with regional warming. Nature 399, 579-583.
, Yohe, G. (2003): A globally coherent fingerprint of climate change impacts across natural systems. Nature 421, 37-42.
Pöyry, J., Luoto, M., Heikkinen, R., Kuussaari, M., Saarinen, K. (2009): Species traits explain recent range shifts of Finnish butterflies. Global Change Biology 15, 732-743.
Roy, D.B., Rothery, P., Moss, D., Pollard, E., Thomas, J.A. (2001): Butterfly numbers and weather: predicting historical trends in abundance and the future effects of climate change. J. Animal Ecology 70, 201-217.
Settele, J., Kudrna, O., Harpke, A., Kühn, I., Swaay, C.V., Verovnik, R., Warren, M., Wiemers, M., Hanspach, J., Hickler, T., Kühn E., Halder, I.v., Veling, K., Vliegenthart, A., Wynhoff, I., Schweiger, O. (2008): Climatic risk atlas of European butterflies. Pensoft (Sofia Moscow), 710 pp.
Thomas, C.D., Bodsworth, E.J., Wilson, R.J., Simmons, A.D., Davies, Z.G., Musche, M., Conradt, L. (2001): Ecological and evolutionary processes at expanding range margins. Nature 411, 577-581.
Thomas, C.D., Cameron, A., Green, R.E., Bakkenes, M., Beaumonz, L.J., Collingham, Y.C., Erasmus, B.F., De Siqeira, M.F., Graininger, A., Hannah, L., Hughes, L., Huntley, B., Van Jaarsfeld, A.S., Midgley, G.F., Miles, L., Ortega-Huerta, M.A., Peterson, A.T., Philipps, O.L., Williams, S.E. (2004): Extinction risk from climate change. Nature 427, 145-148.
, Franco, A.M., Hill, J.K. (2006): Range retractions and extinction in the face of climate warming. Trends in Ecology and Evolution 21, 415-416.
van Swaay, C.A.M., Van Strien, A.J. (2008): The European Butterfly Indicator for Grassland Species 1990 2007. Report VS2008.022. De Vlinderstichting, Wageningen.
Walther, G.R., Post, E., Convey, P., Menzel, A., Parmesan, C., Beebee, T.J., Fromentin, J.M., Hoegh-Guldberg, O., Bairlein, F. (2002): Ecological responses to recent climate change. Nature 416, 389-395.
Anschriften der Verfasser: Dr. Stefan Brunzel, Institut für Biodiversitätsmanagement, Gießener Straße 9, D-35085 Ebsdorfergrund, E-Mail: [email protected] ; Alexandra Aparicio, Lindenstraße 2, D-35287 Amöneburg; Mathias Lohr, Hochschule Ostwestfalen-Lippe, Fachgebiet Landschaftsökologie und Naturschutz, An der Wilhelmshöhe 44, D-37671 Höxter, E-Mail [email protected] ; Dr. Burkhard Beinlich, Landschaftsstation im Kreis Höxter e.V., Zur Specke 4, D-34344 Borgentreich, E-Mail [email protected].
-
Abb. 1a: Abundanzschwankungen in der Untersuchungsregion Märkisches Sauerland. Individuenzahlen entlang von 23 Transekten, standardisiert auf 200 m. -
Abb. 1b: Abundanzschwankungen in der Untersuchungsregion Westliches Weserbergland. Individuenzahlen entlang von acht Transekten, standardisiert auf 200 m. -
Abb. 2: Unterschiedliche Vorzeichen bei signifikanten Trends (mit Regressionskoeffizient) in den beiden Untersuchungsregionen. Dargestellt sind die maximalen jährlichen Individuenzahlen, standardisiert auf 200 m. -
Abb. 3. Entwicklung der einzelnen Gruppen (ANOVA mit Messwiederholungen und post-hoc Tukey-HSD-Test). Signifikanter Unterschied zwischen Gruppen: Wilks‘ Lambda = 0,7173, p ≤ 0,05. Fehlerbalken zeigen 95-%-Konfidenzintervalle (negative Werte abgeschnitten). Als einzelne Gruppe zeigen aber nur die Arten mit Zunahme einen signifikanten Trend, (R = 0,38, p ≤ 0,01). Zwischen den Jahren konnte ein signifikanter Unterschied nur von 1992 2000 zu 2007 bei den Arten mit Zunahme festgestellt werden (Post-hoc Tukey-HSD-Test: p ≤ 0,05). -
Abb. 4: Anteil zu- und abnehmender sowie indifferenter Arten in Lebensraumtypen der Untersuchungsregion Westliches Weserbergland. Arten können in mehrere Lebensräume eingruppiert sein. Zunehmende Arten weisen eine Zunahme > 40 % in der Rasterfrequenz von Zeitraum I (1981-1991) zu III (2001 2010) und von Zeitraum II (1992 2000) zu III auf, Arten mit > 40 % Abnahme in den gleichen Zeiträumen und indifferente Arten mit weniger als 40 % Veränderung der Rasterfrequenz (vgl. Tab. 1). Absolute Artenzahlen in Säulen, signifikante Unterschiede zwischen Lebensräumen in Tab. 3. -
Veränderungen von Tagfaltergemeinschaften -
Veränderungen von Tagfaltergemeinschaften -
Veränderungen von Tagfaltergemeinschaften

Bedienhilfen
Barrierefreiheit
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.