Wiederherstellung von Ökosystemleistungen der Flusstalmoore in Mecklenburg-Vorpommern
Abstracts
In Mecklenburg-Vorpommern wurden Flächen des Moorschutzprogramms (6600 ha) landschaftsökologisch untersucht, um deren Ökosystemleistungen qualitativ und quantitativ zu erfassen. Hierbei lag der Schwerpunkt auf der Vegetationsentwicklung nach einer Wiedervernässung, der oberirdischen Biomasseproduktion und der Bindung von Nährstoffen durch die Sumpf- und Wasserpflanzen und einer Bilanzierung u.a. des Torfbildungspotenzials auf Landschaftsebene. Es zeigte sich, dass insbesondere dem Schilf (Phragmites australis) eine hohe Bedeutung bezüglich der Biomasseproduktion und der Nährstoffspeicherung in wiedervernässten Mooren zukommt.
Im Hinblick auf die Ökosystemleistungen wiedervernässter Moore kann zusammenfassend festgestellt werden, dass das Moorschutzprogramm nach über zehn Jahren Wiedervernässungspraxis eine positive Bilanz ziehen kann, da sich (1) auf den wiedervernässten Moorflächen ein vielfältiges Vegetationsmosaik aus Sumpf- und Wasserpflanzen entwickeln konnte, welches zahlreichen Tierarten einen Lebensraum bietet, (2) erhebliche Stoffmengen (C, N, P) gespeichert werden, die bei entsprechender Abschöpfung (Mahd) oder langfristiger Festlegung (Torfbildung) positiv zu einer Regulation der Nährstoffbilanz in einer durch Eutrophierung gekennzeichneten Landschaft beitragen, und (3) sich bereits jetzt ein erhebliches Potenzial der Torfbildung abzeichnet, eines der wesentlichen Ziele eines nachhaltigen Moorschutzes.
Lowland Fens in Mecklenburg-Western Pomerania Restoration of Ecosystem Services
In the German federal state of Mecklenburg-Western Pomerania lowland fens having been rewetted within the Mire Conservation Program were investigated in order to qualitatively and quantitatively assess ecosystem services. A particular focus was laid on vegetation development after rewetting, above-ground biomass production, and nutrient standing stock by helophytes and water plants and an assessment of peat forming potential on the landscape level. In particular, reed (Phragmites australis) plays a major role regarding biomass production and nutrient storage.
In conclusion, after 10 years of rewetting the Mire Conservation Program in Mecklenburg-Western Pomerania has to be considered a success with regard to the restoration of ecosystem services: (1) A diverse mosaic of different vegetation types has developed providing a valuable habitat for wetland species, (2) considerable amounts of nutrients (C, N, P) are retained which positively contribute to the landscape nutrient balance in an eutrophicated landscape by natural peat accumulation or artificial removal through mowing, and (3) a considerable potential of peat accumulation as one of the main objectives of sustainable mire conservation.
- Veröffentlicht am

1 Einleitung
Einst großflächig verbreitete Moorflächen in Mitteleuropa sind seit dem Beginn des 20. Jahrhunderts zunehmend einer Intensivierung der Landnutzung gewichen. Insbesondere der landwirtschaftliche Nutzungsdruck seit den 1950er Jahren hat mit Entwässerung und einer intensiven Grünlandnutzung zu einer starken Degradation der Moore geführt. Diese Entwicklung führte auch in Mecklenburg-Vorpommern, mit über 300000 ha dem moorreichsten Bundesland, zu einem Verlust der Kohlenstoff-, Nährstoff- und Wasserspeicherfunktion der Moore sowie zu der Zerstörung des Lebensraumes für seltene Tier- und Pflanzenarten (Succow 2001a).
Aufgrund der hohen Bedeutung der Moore für die nordostdeutschen Landschaften und dem mit der Moordegradation einhergehenden Verlust wichtiger Ökosystemleistungen (Timmermann et al. 2009) hat man auf diese Entwicklung mit einem umfassenden Moorschutzkonzept reagiert, welches nicht nur beispielhaft für das norddeutsche Tiefland, sondern auch für ähnliche Landschaften in Mitteleuropa ist. Mit diesem Schutzkonzept gab Mecklenburg-Vorpommern als erstes Bundesland der Renaturierung von Niedermooren einen konzeptionellen und finanziellen Rahmen (Moorschutzprogramm; Kowatsch 2007). Seit nunmehr über zehn Jahren wurden ca. 14000 ha stark degradierter Moore, hauptsächlich tiefgründige Flusstalmoore, wiedervernässt mit dem Ziel einer Nährstoffretention und Wiederbelebung der Torfbildung und Kohlenstofffestlegung (Lenschow & Thiel 2000). Nach der Anhebung des Wasserspiegels entstanden, je nach Ausmaß der Moorsackung und des Moorschwundes sowie der geländemorphologischen Situation, unterschiedlich hoch überstaute Bereiche. In Abhängigkeit von der mittleren Höhe und Dynamik der Wasserstände entwickelten sich neue Ökosysteme mit Flachgewässern und einem Mosaik aus Röhricht- und Großseggenbeständen sowie Grünlandbrachen. Die derzeitige Vegetation ist jedoch noch am Beginn der Sekundärsukzession und demzufolge einem ständigen Wandel unterworfen. Bisher liegen weitgehend nur Einzelerhebungen für den Vegetationswandel wiedervernässter Moore für Mecklenburg-Vorpommern vor (u.a. Timmermann et al. 2006).
Um die Entwicklung dieser neu entstandenen Ökosysteme und ihre Leistungen zu bewerten, ihre Wechselwirkungen mit angrenzenden Ökosystemen abzuschätzen und allgemeine Aussagen für weitere Wiedervernässungsprojekte ableiten zu können, ist eine flächendeckende Untersuchung der Vegetation und deren Funktion für die Nährstoffretention unerlässlich.
Im Rahmen einer vom Landesamt für Umwelt, Naturschutz und Geologie in Mecklenburg-Vorpommern (LUNG MV) in Auftrag gegebenen Studie wurden wiedervernässte Niedermoore in Nordostdeutschland deshalb hinsichtlich folgender Fragestellungen untersucht:
Welche Vegetation entwickelt sich nach der Wiedervernässung und sind regelhafte Trends der Sukzession abzuleiten?
Welche Rolle spielen die dominanten Pflanzenarten hinsichtlich der oberirdischen Nährstoffbindung in der Vegetationsperiode?
Sind flächenhafte Aussagen zur saisonalen Nährstoffbindung und zum Torfbildungspotenzial mittels Fernerkundung auf Landschaftsebene machbar?
Insgesamt liegen dieser Synthese Untersuchungen von 27 Poldern mit einer Gesamtfläche von ca. 6600 ha zugrunde. Die interdisziplinären Forschungsarbeiten wurden in Kooperation mit dem Leibniz-Institut für Gewässerökologie und Binnenfischerei in Berlin (IGB) durchgeführt (Gelbrecht et al. 2008).
2 Untersuchungsgebiet
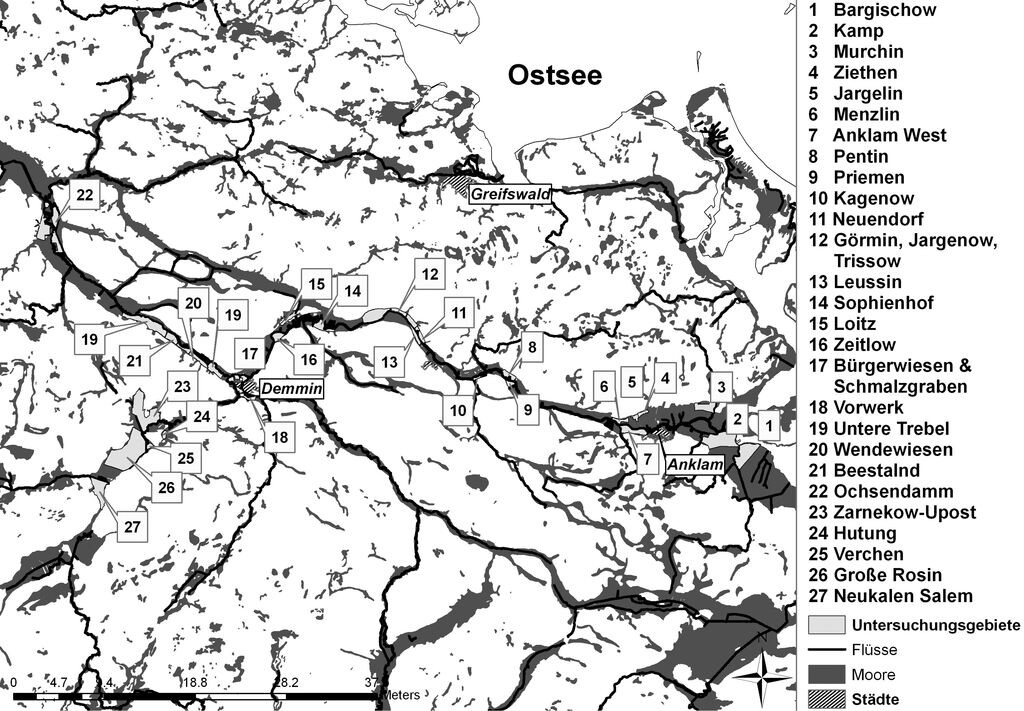
Die Untersuchungen wurden auf wiedervernässten Mooren im Peene- und Trebeltal Mecklenburg-Vorpommerns durchgeführt (Abb. 1), welche zum System der großen Flusstalmoore im Nordosten Deutschlands gehören und primär mehrere Meter mächtige mesotroph-subneutrale Durchströmungsmoore darstellen. Schon Ende des 18. Jahrhunderts unterlagen fast alle Bereiche der Flusstalmoore einer Entwässerung und Kultivierung. Die entscheidende Zäsur kam jedoch durch die Intensivierung der Landwirtschaft mit einhergehender Komplexmelioration zwischen 1960 und 1980. Waren vorher durch kleinbäuerliche Wiesen- und Weidenbewirtschaftung noch artenreiche Moor- und Feuchtwiesen vorhanden, wurde nun die Vegetation durch tiefe Entwässerung, Umbruch, Anlage von Decksandkulturen und Ansaat von Kulturgräsern stark verändert. Es kam zu einer starken Degradation der Moorstandorte mit dann durchweg eutrophem bis polytrophem Charakter (Succow 2001b).
Die Vegetation der Untersuchungsflächen bestand vor 1990 fast ausschließlich aus intensiv genutztem Niedermoorgrünland mit zwei- bis dreimaliger Schnittnutzung und entsprechend hohen Phosphor- und Stickstoffdüngergaben. Dabei handelte es sich überwiegend um Saatgrasland (Plantagini majoris-Lolietum perennis Beger 1932 nom. invers. propos., Pätzolt & Jansen 2004) mit einem hohen Anteil von Rohrglanzgras (Phalaris arundinacea) und weiteren eu- bis polytraphenten Grünlandarten wie Knäuelgras (Dactylis glomerata), Rohrschwingel (Festuca arundinacea) und Quecke (Elytrigia repens). Die Grundwasserstände im Moor lagen vor der Wiedervernässung in den Sommermonaten meist bis mehr als 1 m unter Flur (LUNG 1994-1997). Aufgrund der Zersetzung und Sackung der oberen Moorbodenhorizonte unter das Niveau der angrenzenden Gewässer wurden die Flächen gepoldert und mit Pumpanlagen ausgerüstet. Dadurch kam es zur weiteren Mineralisierung, Verdichtung und Sackung der Torfschichten, nach Succow (2001a) der Teufelskreis der Moornutzung.
3 Methoden
3.1 Erfassung der Vegetations entwicklung und Ableitung von Sukzessionstrends
Für die Vegetationskartierung wurden zwei verschiedene Verfahren angewandt, die Kartierung mit hochauflösenden Satellitendaten (Quickbird: aus dem Jahr 2007; World View 1 und SPOT 2: aus dem Jahr 2008) und die terrestrische Kartierung. Für beide Kartierungsmethoden wurde als Grundlage ein einheitlicher Kartierungsschlüssel unter besonderer Berücksichtigung dominant vorkommender Pflanzenarten bzw. Pflanzenartengruppen verwendet (Steffenhagen et al. 2008a). Die Auswertung der Quickbird-Daten erfolgte mit dem von Frick (2006) entwickelten und hier modifizierten Verfahren zur Vegetationsklassifizierung. Insgesamt wurden mit diesem Verfahren 12 Vegetationstypen unterschieden, die insgesamt 7 dominierende Pflanzenarten und 5 Lebensformengruppen umfassten. Durch Abgleich der Klassifizierungsergebnisse mit terrestrisch gewonnenen Vegetationsaufnahmen wurde die durchschnittliche Genauigkeit der Kartierungsergebnisse bestimmt.
Für die Ableitung allgemeiner Entwicklungstrends der Vegetation nach der Wiedervernässung wurden zahlreiche vorhergehende Einzelbeobachtungen (z.B. Diplomarbeiten am Institut für Botanik und Landschaftsökologie der Universität Greifswald) und Dauerflächenuntersuchungen (z.B. Timmermann et al. 2006) herangezogen. Dafür wurde das Datenmaterial mit dem Programm TURBOVEG bearbeitet und standardisiert. Die anschließende pflanzensoziologische Auswertung erfolgte mittels des Programms JUICE 7.0. Die Ergebnisse wurden mit den eigenen zeitlich späteren, großflächigen Erhebungen verglichen und zu Prognosen von Sekundärsukzessionen in Abhängigkeit von Grundwasserständen herange zogen.
3.2 Bestimmung der Biomasse dominanter Moorpflanzen
3.2.1 Helophyten (Sumpfpflanzen)
Zur Bestimmung der oberirdischen Netto-Primärproduktion (Gesamtproduktion minus Verluste) der dominant vorkommenden Helophyten wurde die Biomasse der Arten Breitblättriger Rohrkolben (Typha latifolia), Schilf (Phragmites australis), Großer Wasserschwaden (Glyceria maxima), Rohrglanzgras (Phalaris arundinacea), Ufer-Segge (Carex riparia) und Sumpf-Segge (Carex acutiformis) geerntet (Steffenhagen 2008). Um Maximalwerte der oberirdischen Biomasse zu ermitteln, erfolgte die Ernte jeweils zum Zeitpunkt beginnender Blüten- bzw. Fruchtbildung. Die auf diese Weise gewonnene Biomasse entspricht in etwa der jährlichen oberirdischen Netto-Primärproduktion von Helophyten-Beständen (Odonk & Kvet 1978) und enthält die höchsten Nährstoffgehalte (Bernhard & Hankinson 1979).
3.2.2 Hydrophyten (Wasserpflanzen)
Schwebematten und Schwimmdecken (Abgrenzung nach der Definition in Wegener 1998) wurden innerhalb der Untersuchungsgebiete an Standorten mit der Wasserstufe 6+ geerntet (Wasserstufen nach Koska 2001a), d.h. auf Flächen mit permanentem Überstau. Die Schwebematten wurden auf diesen Standorten fast ausschließlich von Ceratophyllum demersum gebildet. Lemna minor und Spirodela polyrhiza bildeten den überwiegenden Anteil der Schwimmdecken.
3.3 Analyse der Nährstoffgehalte in der Biomasse dominanter Moorpflanzen
Die Biomasse wurde nach der Trocknung mit einer Schlagkreuzmühle zerkleinert und mit einer Feinmühle (Pulverisette) zu Pulver gemahlen. Anschließend wurde die Kohlenstoff-, Stickstoff- und Phosphorgehalte bestimmt.
Der Gesamt-Kohlenstoff- und der Gesamt-Stickstoffgehalt (DIN ISO 10654 & DIN ISO 13878) wurde mit einem Elementaranalysator der Firma Elementar-Analysesysteme Hanau, mit dem Gerätetyp Vario EL III bestimmt. Das Pflanzenpulver wurde mit H2O2 und 5 M H2SO4 bei 170 °C aufgeschlossen. Die Proben wurden weiterhin mit Ascorbinsäure und Molybdatschwefelsäure versetzt, um anschließend die Extinktion am VARIAN-Photometer CARY 1E bei 710 nm zu messen. Für die Bestimmung des Phosphorgehalts (DIN EN 1189, modifiziert) der Pflanzen sei außerdem auf die detaillierte methodische Darstellung von Gelbrecht et al. (2008) verwiesen.
Die bestimmten Nährstoffmengen für die untersuchten dominanten Pflanzenarten bilden die Grundlage für die Einschätzung der kurzfristigen Nährstoffbindung während der Vegetationsperiode. Unter Stoffbindung soll hier also nur die mögliche saisonale, oberirdische Nährstoffmenge der Pflanzen verstanden werden.
3.4 Nährstoffbindung auf Landschaftsebene
Zur Darstellung der kurzfristig, oberirdischen Nährstoffbindung auf Landschaftsebene wurden die Daten der satellitenbildgestützten Vegetationskartierung mit den Ergebnissen für die Trockenmasse der oberirdischen Biomasse und den Nährstoffgehalten der Pflanzen verknüpft. Anhand der flächenhaften Verteilung der dominant auftretenden Pflanzenarten wurde die mögliche Nährstoffbindung während der Vegetationsperiode auf Karten in ArcGIS 3.2 dargestellt.
3.5 Einschätzung des Torfbildungspotenzials
Torfbildung (verstanden als Netto-Akkumulation organischer Substanz) findet nach bisherigem Kenntnisstand statt, wenn
(1) potenziell torfbildende Vegetation vorhanden ist, d.h. für die untersuchten nährstoffreichen Moore, dass Schilf oder Seggen vorherrschen (Oswit et al. 1976) und
(2) eine langjährig stabile langzeitige bis permanente Wassersättigung besteht, wodurch Abbauprozesse vermindert bzw. weitgehend unterbleiben (Grosse-Brauckmann 1990).
Zur Einschätzung des Torfbildungspotenzials wurden terrestrisch gewonnene Wasserstufenkarten anhand der Vegetationstypen (nach Koska 2001b) für fünf untersuchte Polder (165 ha) angefertigt. Diese wurden anschließend mit den Vegetationskarten in ArcGIS 3.2 verschnitten. Hinsichtlich der Torfbildungswahrscheinlichkeit wurden für die wiedervernässten Moore drei Flächentypen nach folgenden Kriterien abgeleitet:
Potenzielle Torfbildung auf Flächen in denen beide Voraussetzungen für Torfbildung erfüllt sind (s.o.). Torfbildung kann hier möglicherweise schon stattfinden.
Zukünftige potenzielle Torfbildung auf Flächen mit permanentem Überstau oder ganzjährig mit Wasser gesättigtem Wurzelbereich, in denen die vorherrschende Vegetation im Verlauf der Sekundärsukzession durch potenziell torfbildende Vegetation ersetzt werden kann (vgl. Timmermann et al. 2006).
Torferhaltung bzw. Torfzehrung auf Flächen mit Wasserstands-Medianen unter Flur bzw. mit stark wechselnden Wasserstands-Medianen, die sich größtenteils unter Flur befinden.
4 Ergebnisse
4.1 Vegetationsentwicklung nach der Wiedervernässung
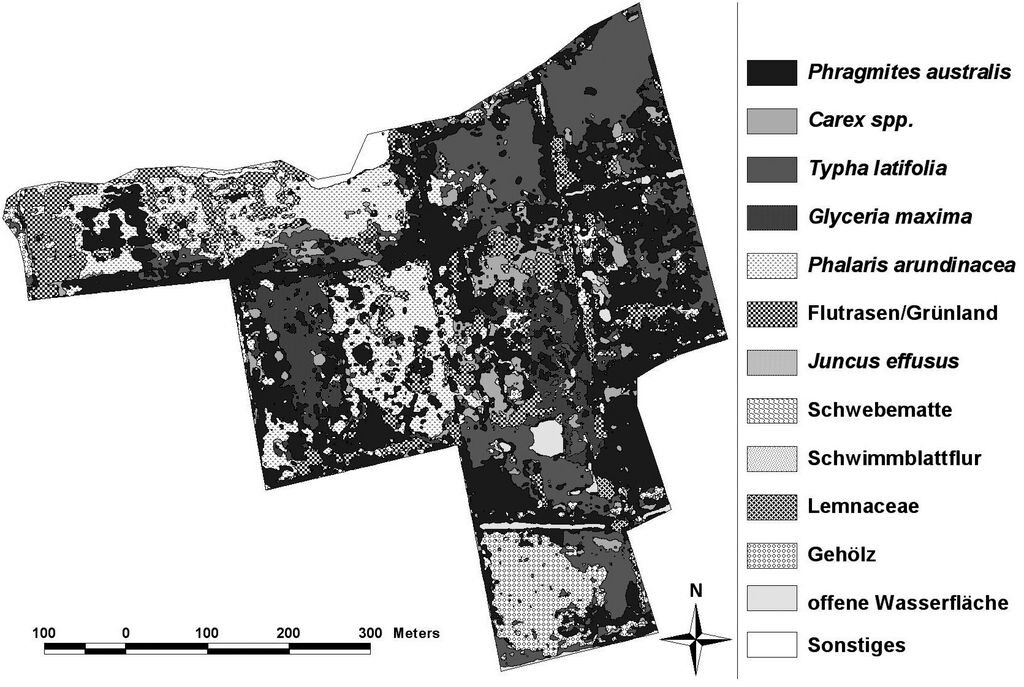
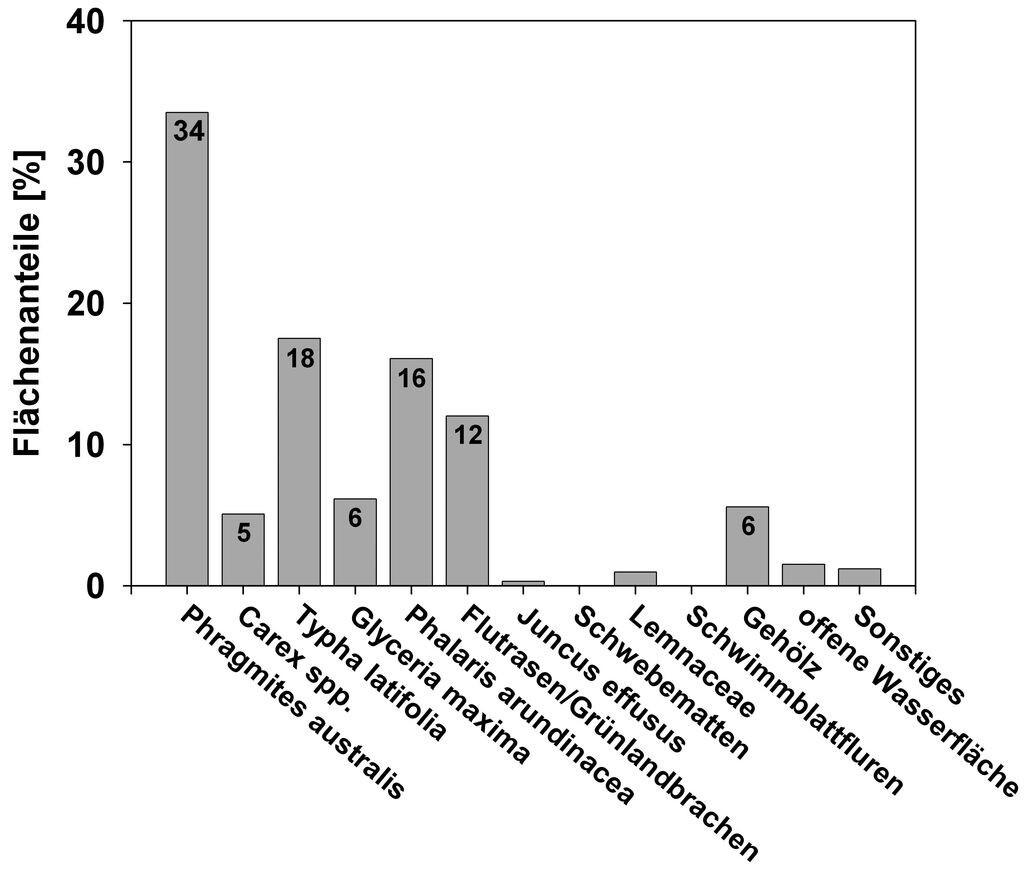
Insgesamt konnte für alle differenzierten Vegetationsklassen eine durchschnittliche Genauigkeit von 80 bis 85 % erreicht werden (vgl. Frick 2006). Exemplarisch werden hier jedoch nur die Kartierungsergebnisse für den 32,5 ha großen Polder Jargelin vorgestellt. Vor der Wiedervernässung im Jahr 1995 dominierte hier artenarmes Rohrglanzgras-Grünland. Abb. 2 stellt dagegen die flächenhafte Verteilung der Vegetation mit elf Vegetationstypen im Polder Jargelin im Jahr 2007 dar. Zwölf Jahre nach der Vernässung wird die Vegetation fast ausschließlich durch Röhrichte und Seggenriede bestimmt. Hauptbestandsbildner ist Schilf, welches bereits ein Drittel der Gesamtfläche einnimmt (Abb. 3). Den zweitgrößten Anteil nehmen die Röhrichte mit Breitblättrigem Rohrkolben mit einer Fläche von ca. 18 % ein. Großseggenriede mit Carex acutiformis und C. riparia sind nur mit einem Flächenanteil von rund 5 % vertreten. Die ursprüngliche Ausgangsvegetation (Rohrglanzgras) wurde demnach auf den nassesten Standorten nahezu vollständig durch Röhrichte und Seggenriede ersetzt und nimmt nur noch einen Flächenanteil von 16 % ein. Die terrestrische Vegetationskartierung ergab außerdem, dass sich im Zentrum des Polders bereits Tendenzen einer Nährstoffverarmung abzeichnen. Hier hat sich kleinflächig ein Mosaik aus Schnabel-Segge (Carex rostrata), Zweizeiliger Segge (C. disticha), Teich-Schachtelhalm (Equisetum fluviatile), Sumpf-Vergißmeinnicht (Myosotis scorpioides), Wasserschierling (Cicuta virosa), Sumpf-Sternmiere (Stellaria palustris) und Wasser-Minze (Mentha aquatica) entwickelt.
4.2 Biomasse und Nähr stoff bin dung der dominanten Moorpflanzen
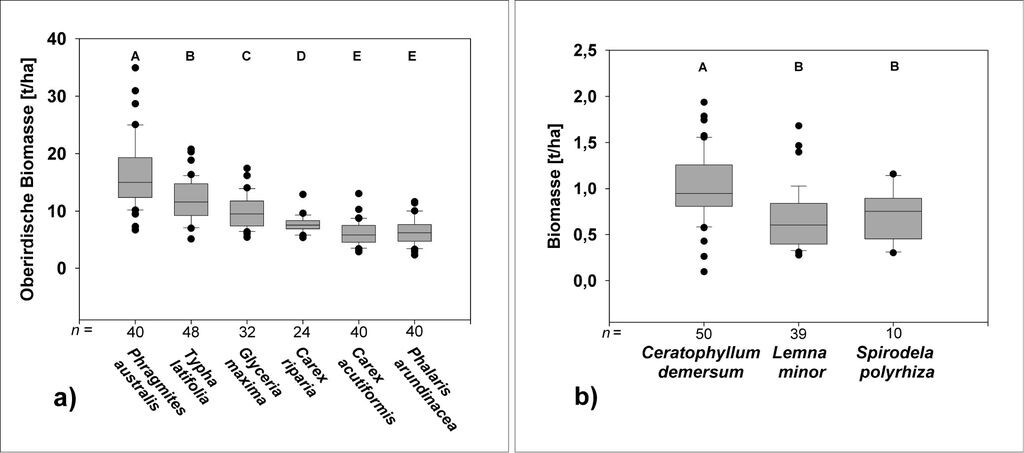
Die oberirdische Biomasseproduktion der Helophyten erreichte im Mittel Werte von 6,1 bis 16,6 t Trockensubstanz (TS) pro ha mit meist signifikanten Unterschieden zwischen den einzelnen Arten (Abb. 4a). Besonders hoch war die Biomasseproduktion von Schilf, welche mit 12,5 bis 23,8 t TS/ha über den Werten der anderen dominanten Pflanzenarten lag. Für Typha latifolia und Glyceria maxima ließ sich mit durchschnittlich 12,1 t TS/ha bzw. 9,8 t TS/ha ebenfalls ein hohes Produktionsniveau nachweisen. Phalaris arundinacea (6,5 t TS/ha) und Carex acutiformis (6,1 t TS/ha) unterschieden sich nicht signifikant.
Die Biomasseproduktion von Ceratophyllum demersum lag zwischen 0,86 und 1,19 t TS/ha (Abb. 4b). Die Werte der Lemnaceen lagen mit 0,67 (Lemna minor) und 0,71 t TS/ha (Spirodela polyrhiza) deutlich unter der Biomasseproduktion von Ceratophyllum demersum. Untereinander wiesen die beiden Lemnaceen kaum Unterschiede in der Biomasseproduktion auf.
Der Kohlenstoffgehalt der Helophyten lag bei durchschnittlich 46,0 % Kohlenstoff in der Trockensubstanz (= C/TS) (Tab. 1). Dabei war Phragmites australis aufgrund der hohen Biomasseproduktion in der Lage, die doppelte Kohlenstoffmenge in organische Substanz umzuwandeln als die Großseggenarten.
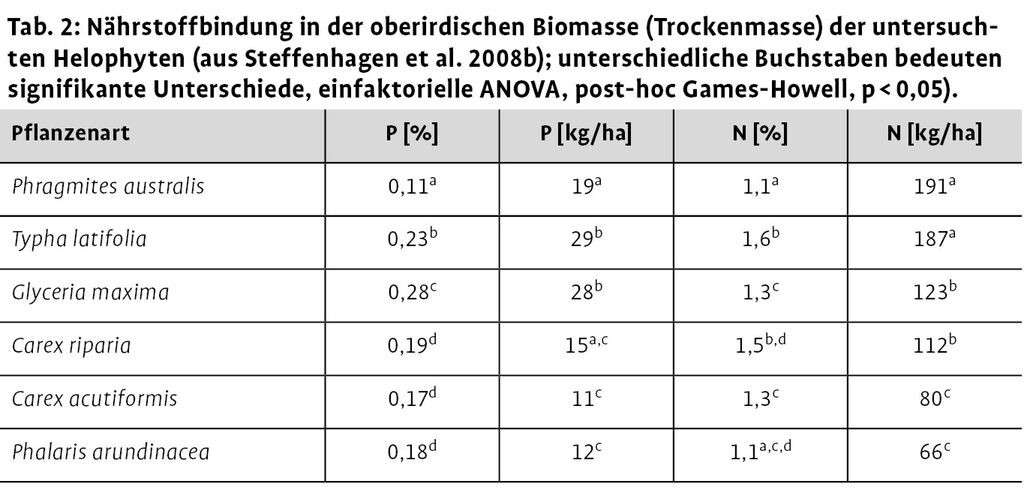
Für die in der oberirdischen Biomasse eingelagerten Phosphor- und Stickstoffgehalte zum maximalen Entwicklungszeitpunkt, ließen sich signifikante Unterschiede zwischen den Pflanzenarten nachweisen (Tab. 2). Auf Basis der in Tab. 2 angegebenen mittleren P-Gehalte in der Trockensubstanz und den Ergebnissen der oberirdischen Biomasseermittlung (Abb. 4a) ergab sich, dass Typha latifolia und Glyceria maxima die höchsten Phosphormengen mit 29 und 28 kg P/ha in der oberirdischen Biomasse enthielten. Die höchsten Stickstoffmengen in der oberirdischen Biomasse konnten für Phragmites australis (191 kg N/ha) und Typha latifolia (187 kg N/ha) nachgewiesen werden.
4.3 Nährstoffbindung auf Landschaftsebene
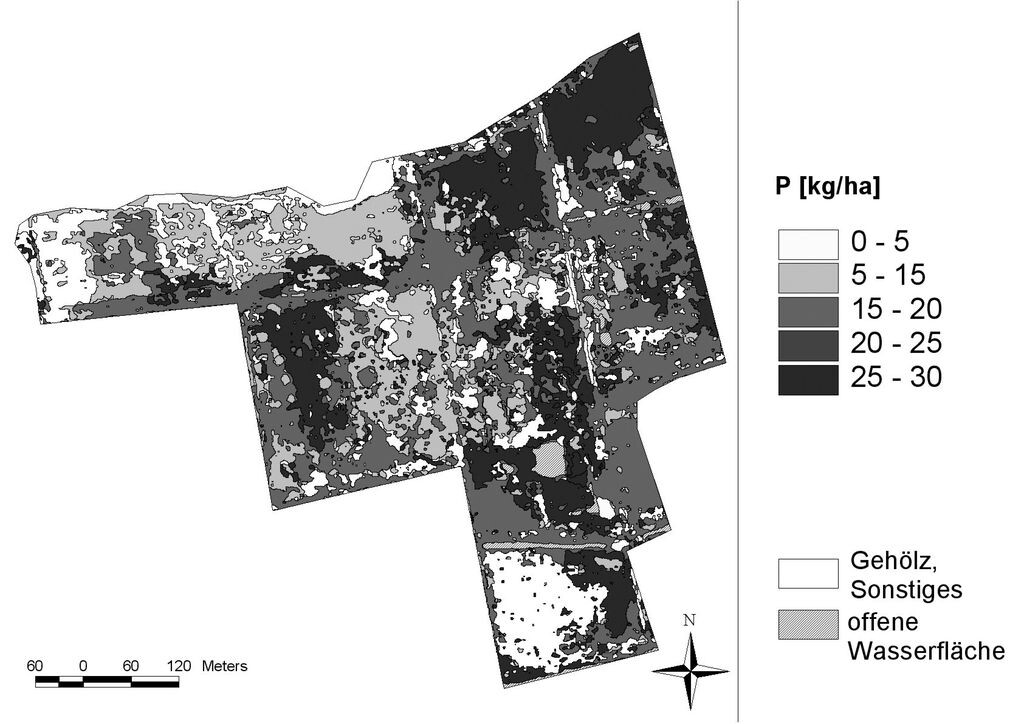
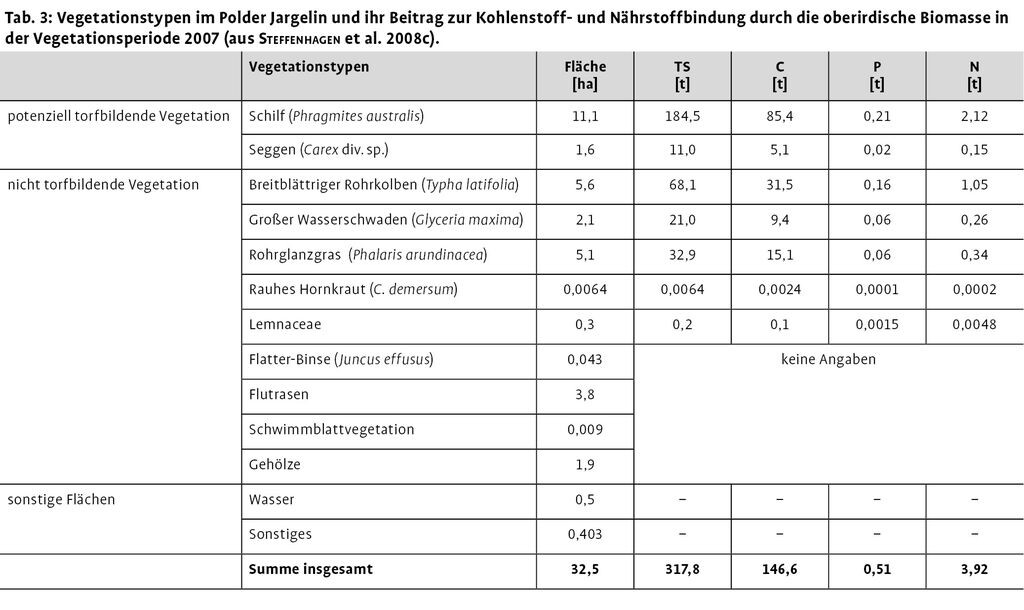
Beispielhaft soll hier für den Polder Jargelin die flächenhafte Verteilung der dominant auftretenden Pflanzenarten genutzt werden, um die Stoffbindung in der oberirdischen Biomasse der Hauptbestandsbildner auf Landschaftsebene beurteilen zu können (Tab. 3). Die in einer Vegetationsperiode oberirdisch gebundene Kohlenstoffmenge in dem 32,5 ha großen Polder Jargelin lag bei insgesamt ca. 145 t, was gleichzeitig einer Kohlendioxidfixierung von ca. 540 t durch die oberirdische Biomasse entspricht. Zum Zeitpunkt der maximalen Biomasseausbildung wurden außerdem rund 4 t Stickstoff, 2 t Kalium und 0,5 t Phosphor in der oberirdischen Vegetation gebunden. Das Phosphorbindungspotenzial durch die pflanzliche oberirdische Biomasse wird am Beispiel des Polders Jargelin in Abb. 5 veranschaulicht. Die Daten liefern einen ersten Überblick zur Menge sowie räumlich-standörtlichen Differenzierung der in der Sprossbiomasse gebundenen Nährstoffe zum maximalen Entwicklungszeitpunkt der oberirdischen Biomasse innerhalb eines gesamten Polders.
4.4 Einschätzung des Torfbildungspotenzials
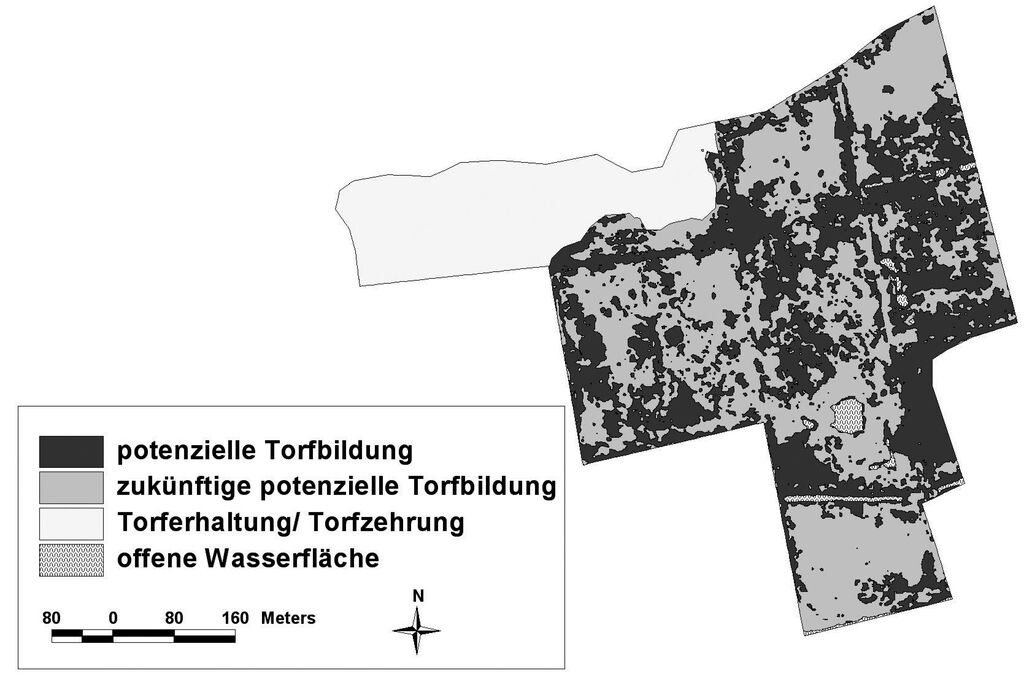
Bei der Einschätzung des Torfbildungspotenzials wurde deutlich, dass fünf bis zwölf Jahre nach der Wiedervernässung die Flächen mit potenzieller Torfbildung ein Viertel bis fast die Hälfte der Gesamtfläche der Untersuchungsgebiete einnehmen können. Flächen mit zukünftiger potenzieller Torfbildung in den fünf Untersuchungsgebieten wiesen mit 27 bis 49 % ähnlich hohe Anteile auf wie die Flächen mit potenzieller Torfbildung. Den weitaus niedrigsten Flächenanteil in allen Untersuchungsgebieten nahmen die Areale ein, in denen aufgrund unzureichender Vernässung nicht mit einer Torfbildung zu rechnen ist (17 bis 23 %). Diese Flächen befanden sich in Talrandnähe bzw. an den Deichanlagen des jeweiligen Polders. Beispielhaft wurden hier die ermittelten Torfbildungspotenziale des Polder Jargelins dargestellt (Abb. 6).
5 Diskussion
5.1 Vegetationsentwicklung und Sukzessionstrends
Mit dem von Frick (2006) entwickelten Analyseverfahren von Satellitendaten konnte erstmals die Vegetation wiedervernässter Niedermoore in Mecklenburg-Vorpommern großflächig mittels Fernerkundung in hoher Auflösung kartiert werden. Die Vegetationsklassifizierung bietet zahlreiche Vorteile wie z.B. hohe Klassifikationsgenauigkeit, Reproduzierbarkeit, Kartierung auch von schwer zugänglichen Gebieten und eine direkte simultane Flächenquantifizierung. Damit stehen auch Grundlagen für ein zeitsparendes und kontinuierliches Monitoring zur Verfügung. In Abhängigkeit von der ursprünglichen Nutzungsintensität bzw. der Moordegradation und der hydrologischen Bedingungen nach der Wiedervernässung (z.B. Wasserstände) haben sich für jedes Untersuchungsgebiet individuelle Vegetationsmuster ergeben. Insgesamt zeichneten sich jedoch für die untersuchten wiedervernässten Moore ähnliche Tendenzen bzw. Gesetzmäßigkeiten bei der Vegetationsentwicklung in folgenden Punkten ab:
Die Ausgangsvegetation mit Phalaris arundinacea-Grünland, Flutrasengesellschaften oder Grünlandbrachen war nur noch in Resten an höher gelegenen Bereichen wie Talrändern oder Inseln mit mineralischem Boden zu finden.
Das Vorhandensein von Seggen und Schilf in der Ausgangsvegetation beschleunigt wesentlich die Ausbreitung von potenziell torfbildender Vegetation nach der Wiedervernässung.
Nahezu die Hälfte der Flächen wurde von offenen Wasserflächen eingenommen und es bildeten sich Flachseen über Niedermoortorf, deren Verlandung durch Ausbildung von Schwebematten und Lemnaceae eingeleitet wird, welche zu starker Schlammbildung beitragen können.
Bereits relativ kurze Zeit nach der Wiedervernässung (nach etwa fünf Jahren) konnten sich Röhrichte und Seggenriede etablieren und waren z.T. Hauptbestandsbildner. Insbesondere Schilf (Phragmites australis), aber auch andere Ried- und Röhrichtarten konnten sich vielerorts von den Grabenrändern ausbreiten, sowie inselartig neue Populationen begründen.
Bereits fünf bis zwölf Jahre nach der Wiedervernässung nahm potenziell torfbildende Vegetation (v.a. mit Phragmites australis und Carex div. sp.) im Mittel 15 bis 20 %, stellenweise ein Viertel bis sogar die Hälfte der wiedervernässten Moore ein.
Kleinflächig waren bereits Entwicklungstendenzen zu artenreicherer Vegetation (Braunmoos-Seggenriede) und mesotrophen Standortsverhältnissen zu beobachten.
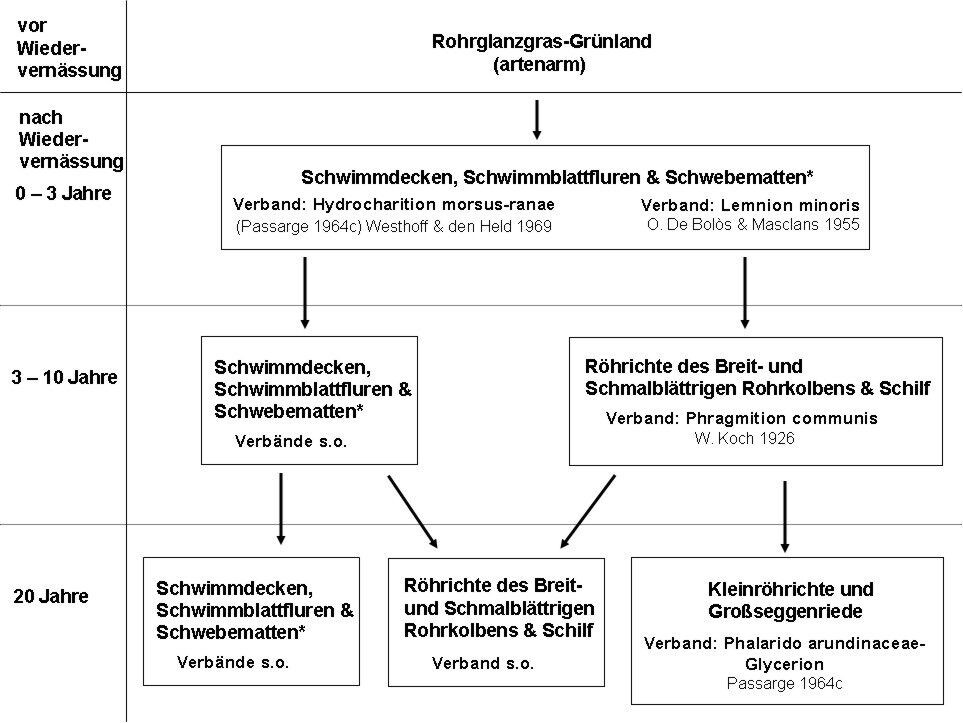
Insbesondere die Vegetationsentwicklung in den schon länger vernässten Untersuchungsgebieten (ab 1995 bis 1999) wies einen hohen Anteil an potenziell torfbildender Vegetation und kleinflächig nährstoffärmeren Bereichen mit artenreicheren Pflanzengesellschaften auf (vgl. Polder Jargelin, Abschnitt 4.1). Diese positiv zu bewertenden Entwicklungen decken sich mit den Zielstellungen des Moorschutzkonzeptes Mecklenburg-Vorpommerns (Lenschow & Thiel 2000). Trotz der schnellen Etablierung von Röhrichten und Seggenrieden wird jedoch die Verlandung der größeren offenen Wasserflächen, aufgrund der ungünstigen Bedingungen für die Ausbreitung und Etablierung von Helophyten (durch stärkere Wasserbewegung, Lichtmangel und dauerhaft hohem Überstau), in Abhängigkeit von der Wassertiefe, wahrscheinlich mindestens einige Jahrzehnte in Anspruch nehmen. In Abb. 7 werden beispielhaft für hoch überstaute Moore als Entwicklungsstadien bzw. Zeitschnitte frühe Initialstadien (ca. 0-3 Jahre nach Vernässung), mittlere Initialstadien (etwa drei bis zehn Jahre nach Vernässung) und ein hypothetisches Folgestadium (ca. 20 Jahre nach Vernässung) dargestellt.
5.2 Nährstoffbindung in der Biomasse
Hinsichtlich der Biomasseproduktion und der Bindung von Kohlenstoff, Phosphor und Stickstoff konnte insgesamt für die Helophyten sowie Schwimmdecken und Schwebematten eine beträchtliche Filterwirkung während der Vegetationsperiode nachgewiesen werden. Dabei zeigten sich insbesondere bei den Helophyten deutliche artspezifische Unterschiede. Insgesamt wurden Werte für die oberirdische Biomasseproduktion erreicht und zum Teil übertroffen, die für Niedermoore mit intensiver Grünlandnutzung (überwiegend Phalaris arundinacea-Grünland) vor der Wiedervernässung mit 9 bis 23 t TS/ha Erntebiomasse belegt sind (Schrautzer 2001). Die hohe Biomasseproduktion von Schilf wird auch durch andere Autoren mit Angaben von Maximalwerten von 20 bis 43 t TS/ha bestätigt (Kühl & Kohl 1992). Mit Blick auf die Hydrophyten muss von erheblichen Schwankungen der Biomasseproduktion während der Vegetationsperiode und in Abhängigkeit des Nährstoffgehalts des Wassers ausgegangen werden (Körner 1996). Dies gilt auch für die Lemnaceen bei denen eine Verdopplung der Biomasse innerhalb von mehreren Tagen bis Wochen erfolgen kann (Landolt 1996).
Die Ergebnisse zur Bindung von Kohlenstoff und Nährstoffen unterstreichen das Vermögen von Schilf, aufgrund seiner hohen Produktivität entsprechend hohe Nährstoffmengen in der oberirdischen Biomasse während der Vegetationsperiode einzulagern (Dykyjová 1978). Die Fähigkeit der Helophyten, kurzfristig erhebliche Phosphormengen zu binden, senkt während der Vegetationsperiode das potenzielle Austragsrisiko von gelöstem P aus wiedervernässten und überstauten Niedermooren (Poldern) in angrenzende Gewässer (Gelbrecht et al. 2008). Aufgrund des hohen Nährstoffentzugspotenzials werden Helophyten vielfach genutzt, um nährstoffbelastetes Abwasser in künstlich angelegten Feuchtgebieten zu filtern, einerseits durch Nährstoffentzug und andererseits durch Schaffung eines günstigen Milieus für Mikroorganismen, die am Nährstoffabbau beteiligt sind (Vymazal 2006).
Durch Auswaschung und Zersetzung werden die verbliebenen Nährstoffe nach dem Absterben der oberirdischen Biomasse sehr schnell wieder freigesetzt (Vymazal 2006). Vor dem Absterben der oberirdischen Biomasse wird aber ein Teil (25 50 %) der Nährstoffe durch Translokation in die Wurzeln verlagert (Gumbricht 1993). Die Blattstreu von Phragmites australis und Typha latifolia kann innerhalb von 1,5 bzw. 2,5 Jahren vollständig zersetzt sein (Mason & Bryant 1975) und es ist davon auszugehen, dass etwa 90 % der in der oberirdischen Biomasse verbliebenen Phosphormenge aus der Pflanzenstreu wieder freigesetzt wird (Koerselman & Verhoeven 1992). In den untersuchten wiedervernässten und dauerhaft überstauten Niedermooren des Peenetals gelangt jedoch ein Teil der abgestorbenen oberirdischen Biomasse der Helophyten am Sediment unter anaerobe Bedingungen. Dadurch läuft die Zersetzung im Vergleich zur belüfteten Bodenzone vermutlich nur unvollständig und stark verlangsamt ab. Das würde zu einer deutlich höheren dauerhaften P- und C-Akkumulation im neu gebildeten limnischen Sediment führen.
Eine entscheidende Rolle für die Stoffspeicherung in den wiedervernässten Mooren spielt der Flächenanteil der Röhrichtarten Phragmites australis, Typha latifolia und Glyceria maxima, da diese aufgrund ihrer hohen Produktivität sowie z.T. sehr hohen Nährstoffgehalte die höchsten Nährstoffmengen in der oberirdischen Biomasse binden können. Der hohe Flächenanteil von Schilf ist als besonders positiv zu bewerten, da Schilf einerseits ein hohes Torfbildungspotenzial besitzt (Oswit et al. 1976) und andererseits in der Lage ist, eine enorme unterirdische Biomasse auszubilden, die während des Winterhalbjahrs entsprechend hohe Nährstoffmengen speichern kann (Kvet & Husák 1978). Bei einer weiteren Ausbreitung von Schilf im Verlauf der Sekundärsukzession (vgl. Abb. 7) ist deshalb von einer zunehmenden Nährstoffspeicherung in den wiedervernässten Poldern auszugehen.
5.3 Torfbildungspotenziale
Ein Ziel der Wiedervernässung ist, dass Moore langfristig wieder Nährstoffe und Kohlenstoff speichern können, d.h. als Senke (sink) in der Landschaft fungieren (Succow 2001a). Dabei sind Torfbildung und Sedimentation die einzigen Stoffspeicherprozesse, die langfristig wirksam sind (Vymazal 2006). Die Untersuchungen haben gezeigt, dass eine Torfbildung potenziell schon in großen Bereichen der wiedervernässten Moore stattfinden kann, da mit der Anhebung des Wasserspiegels und dem Vorhandensein torfbildender Vegetation, zwei wichtige Grundvoraussetzungen gegeben sind. Außerdem ist anzunehmen, dass sich die Großseggen und insbesondere Schilf im Verlauf der Sekundärsukzession weiter stark ausbreiten werden (vgl. Abschnitt 5.1) und sich somit die Fläche der Standorte mit potenzieller Torfbildung stetig vergrößern wird. Die Wiedervernässung führte aber auch auf Flächen mit Wasserstands-Medianen unter Flur bzw. mit stark wechselnden Wasserstands-Medianen zu einer Verringerung bzw. Unterbindung der Torfzehrung. Damit sind auch diese Flächen hinsichtlich der Klimawirkung und einer zu erwartenden verminderten Nährstofffreisetzung im Vergleich zum intensiv genutztem Niedermoorgrünland als positiv zu bewerten (Meyer et al. 2001). Insgesamt muss hervorgehoben werden, dass der mit ca. 25 bis 50 % hohe Flächenanteil von Arealen mit potenzieller Torfbildung einen wesentlichen Erfolg des Moorschutzprogramms von Mecklenburg-Vorpommern darstellt.
6 Schlussfolgerungen
Im Hinblick auf die hier betrachteten Ökosystemleistungen wiedervernässter Moore kann zusammenfassend festgestellt werden, dass das Moorschutzprogramm in Mecklenburg-Vorpommern nach über zehn Jahren Wiedervernässung eine erfolgreiche Bilanz ziehen kann:
(1) Auf den wiedervernässten Moorflächen zeigt sich im landesweiten Vergleich eine Vegetationsentwicklung hin zu einem vielfältigen Mosaik aus Sumpf- und Wasserpflanzen, welches für moor- und feuchtgebietsspezifische Arten einen bedeutenden Lebensraum darstellt.
(2) Die hohe saisonale Stoffbindung in der Vegetation ermöglicht bei Biomasseentzug (Kim & Geary 2001) oder bei einsetzender Torfbildung eine erhebliche Reduktion der Nährstoffflüsse in einer durch Eutrophierung gekennzeichneten Landschaft.
(3) Bereits jetzt zeichnet sich ein erhebliches Potenzial der Torfbildung ab, eines der wesentlichen Ziele eines nachhaltigen Moorschutzes.
Dank
Die Untersuchungen wurden vom Landesamt für Umwelt, Naturschutz und Geologie in Mecklenburg-Vorpommern (LUNG MV) finanziell unterstützt. Namentlich danken wir Dr. Uwe Lenschow für die Unterstützung und wertvolle Anregungen zu der Forschungsarbeit. Dr. Jörg Gelbrecht und Dr. Dominik Zak (Institut für Gewässerökologie und Binnenfischerei Berlin) sowie Ulrich Möbius (Universität Greifswald) danken wir für die Unterstützung der umfangreichen Laboranalysen.
Literatur
Berg, C., Dengler, J., Abdank, A., Isermann, M. (Hrsg., 2004): Die Pflanzengesellschaften Mecklenburg-Vorpommerns und ihre Gefährdung Textband. Weissdorn, Jena, 606 S.
Bernhard, J.M., Hankinson, G. (1979): Seasonal changes in standing crop, primary production and nutrient levels in a Carex rostrata wetland. Oikos 32, 328-336.
Dykyjová, D. (1978): Nutrient uptake by littoral communities of helophytes. In: Dykyjová, D., Kvet, J., Pond Littoral Ecosystems Structure and functioning, Springer, Berlin, Heidelberg, New York, 257-277.
Frick, A. (2006): Beiträge hochauflösender Satellitenfernerkundung zum FFH-Monitoring Entwicklung eines wissensbasierten Klassifikationsverfahrens und Anwendung in Brandenburg. Diss. TU Berlin, 226 S.
Gelbrecht, J., Zak, D., Augustin, J. (Hrsg., 2008): Phosphor- und Kohlenstoff-Dynamik und Vegetationsentwicklung in wiedervernässten Mooren des Peenetals in Mecklenburg-Vorpommern Status, Steuergrößen und Handlungsmöglichkeiten. Ber. IGB 26, 1-177.
Grosse-Brauckmann, G. (1990): Ablagerungen der Moore. In: Göttlich, K., Hrsg., Moor- und Torfkunde, Schweizerbartsche Verlagsbuchh., Stuttgart, 175-236.
Gumbricht, T. (1993): Nutrient removal processes in freshwater submersed macrophytes systems. Ecological Engineering 2, 1-30.
Kim, S.Y., Geary, P.M. (2001): The impact of biomass harvesting on phosphorus uptake by wetland plants. Water Science and Technology 44, 61-67.
Körner, S. (1996): Selbstreinigungsprozesse imKlärwerksableiter Wuhle unter besonderer Berücksichtigung der Submersen Makrophyten. Diss. Humboldt- Universität Berlin,122 S.
Koerselman, W., Verhoeven, J.T.A. (1992): Nutrient dynamics in mires of various trophic status: nutrient inputs and outputs and the internal nutrient cycle. In: Verhoeven, J.T.A., ed., Fens and Bogs in the Netherlands: Vegetation, History, Nutrient Dynamics and Conservation, Geobotany 18, 397-432.
Koska, I. (2001a): Ökohydrologische Kennzeichnung von Moorstandorten. In: Succow, M., Joosten, H., Hrsg., Landschaftsökologische Moorkunde, Schweizerbartsche Verlagsbuchhandlung, Stuttgart, 92-111.
(2001b): Standortkundliche Kennzeichnung und Bioindikation. In: Succow, M., Joosten, H., Hrsg., Landschaftsökologische Moorkunde, Schweizerbartsche Verlagsbuchh., Stuttgart, 128-143.
Kowatsch, A. (2007): Moorschutzkonzepte und -programme in Deutschland. Ein historischer und aktueller Überblick. Naturschutz und Landschaftsplanung 39, 197-204.
Kühl, H., Kohl, J.G. (1992): Nitrogen accumulation, productivity and stability of reed stands (Phragmites australis (Cav.) Trin. ex Steudel) at different lakes and sites of the lake districts Uckermark and Mark Brandenburg. Internat. Rev. ges. Hydrobiol. 77, 85-107.
Kvet, J., Husák, . (1978): Primary data on biomass and production estimates in typical stands of fishpond littoral plant communities. In: Dykyjová, D., Kvet, J., eds., Pond littoral ecosystems structure and functioning, Springer, Berlin, Heidelberg, New York, 211-216.
Landolt, E. (1996): Duckweeds (Lemnaceae): Morphological and ecological characteristics and their potential for recycling of nutrients. Environmental Research Forum 5-6, 289-296.
Lenschow, U., Thiel, W. (2000): Das Moorschutzkonzept des Landes Mecklenburg-Vorpommern. Ansätze zur Lösung der durch Entwässerung verursachten ökonomischen und ökologischen Probleme. Natur und Landschaft 75, 317-322.
LUNG (19941997): Moorstandortkatalog Mecklenburg-Vorpommern. Landesamt für Umwelt, Naturschutz und Geologie, Datenkatalog, Abt. 4, Güstrow.
Mason, J.Y., Bryant, R.J. (1975): Production, nutrient content and decomposition of Phragmites communis Trin. and Typha angustifolia L. J. Ecol. 63, 71-95.
Meyer, K., Höper, H., Blankenburg, J. (2001): Spurengashaushalt und Klimabilanz bei Vernässung. In: Kratz, R., Pfadenhauer, J., Hrsg., Ökosystemmanagement für Niedermoore: Strategien und Verfahren zur Renaturierung, Ulmer, Stuttgart, 104-110.
Odonk, J.P., Kvet, J. (1978): Selection of sampling areas in assessment of production. In: Dykyjová, D., Kvet, J., Pond Littoral Ecosystems Structure and functioning, Springer, Berlin, Heidelberg, New York, 163-174.
Oswit, J., Pacowski, R., S. Zurek (1976). Characteristics of more important peat species in Poland. In: Peatlands and their utilization in Poland, V. International Peat Congress Poznan, NOT, Warsaw, 51-60.
Pätzolt, J., Jansen, F. (2004): Molinio-Arrhenatheretea Tx. 1937 Wirtschaftsgrünland. In: Berg, C., Dengler, J., Abdank, A., Isermann, M., Hrsg., Die Pflanzengesellschaften Mecklenburg-Vorpommerns und ihre Gefährdung, Weißdorn, Jena, 336-353.
Schrautzer, J. (2001): Niedermoore Schleswig-Holsteins: Charakterisierung und Beurteilung ihrer Funktion im Landschaftshaushalt. Habil.-Schr., Christian-Albrechts- Universität, Kiel, 350 S.
Steffenhagen, P. (2008): Probenahme der Helophyten (Sumpfpflanzen) und Hydrophyten (Wasserpflanzen). In: Gelbrecht, J., Zak, D., Augustin, J., Hrsg., Phosphor- und Kohlens toff-Dynamik und Vegetationsentwicklung in wiedervernässten Mooren des Peenetals in Mecklenburg-Vorpommern Status, Steuergrößen und Handlungsmöglichkeiten, Ber. IGB 26, 44-45.
, Frick, A., Timmermann, T., Zerbe, S. (2008a): Satellitenbildgestützte Vegetationsklassifizierung unter besonderer Berücksichtigung dominanter Pflanzenarten. In: Gelbrecht, J., Zak, D., Augustin, J., Hrsg., Phosphor- und Kohlenstoff-Dynamik und Vegetationsentwicklung in wiedervernässten Mooren des Peenetals in Mecklenburg-Vorpommern Status, Steuergrößen und Handlungsmöglichkeiten, Ber. IGB 26, 143-145.
, Timmermann, T., Schulz, K., Zerbe, S. (2008b): Biomasseproduktion sowie Kohlenstoff- und Nährstoffspeicherung durch Sumpfpflanzen (Helophyten) und Wasserpflanzen (Hydrophyten). In: Gelbrecht, J., Zak, D., Augustin, J., Hrsg., Phosphor- und Kohlenstoff-Dynamik und Vegetationsentwicklung in wiedervernässten Mooren des Peenetals in Mecklenburg-Vorpommern Status, Steuergrößen und Handlungsmöglichkeiten, Ber. IGB 26, 145-150.
, Frick, A., Timmermann, T. & Zerbe, S. (2008c): Satellitenbildgestützte Bewertung auf Landschaftsebene. In: Gelbrecht, J., Zak, D., Augustin, J., Hrsg., Phosphor- und Kohlenstoff-Dynamik und Vegetationsentwicklung in wiedervernässten Mooren des Peenetals in Mecklenburg-Vorpommern Status, Steuergrößen und Handlungsmöglichkeiten, Ber. IGB 26, 155-158.
Succow, M. (2001a): Zusammenfassende Beurteilung der Folgen tiefgreifender agrarischer Nutzungsintensivierung der letzten Jahrzehnte auf die Niedermoorstandorte Nordostdeutschlands. In: Succow, M., Joosten, H., Hrsg., Landschaftsökologische Moorkunde, 2. überarb. Auflage, Schweizerbartsche Verlagsbuchh., Stuttgart, 463-470.
Succow (2001b): Ökologisch (-phytozoenologische) Moortypen. In: Succow, M., Joosten, H., Hrsg., Landschaftsökologische Moorkunde, 2. überarb. Auflage, Schweizerbartsche Verlagsbuchh., Stuttgart, 229-234.
Timmermann, T., Joosten, H., Succow, M. (2009): Restaurierung von Mooren. In: Zerbe, S., Wiegleb, G., Hrsg., Renaturierung von Ökosystemen in Mitteleuropa, Spektrum, Heidelberg, 55-93.
, Margoczi, K., Takács, G., Vegelin, K. (2006): Restoration of peat-forming species-poor fen grasslands. Applied Vegetation Science 9, 241-250.
Vymazal, J. (2006): Removal of nutrients in various types of constructed wetlands. Science of the Total Environment 380, 48-65.
Wegener, U. (1998): Naturschutz in der Kulturlandschaft Schutz und Pflege von Lebensräumen. Gustav Fischer, Jena, 456 S.
Anschriften der Verfasser(innen): Peggy Steffenhagen, Karsten Schulz und Dr. Tiemo Timmermann, Institut für Botanik und Landschaftsökologie, Universität Greifswald, Grimmer Straße 88, D-17487 Greifswald, E-Mail tiemo@uni-greifswald.de ; Prof. Dr. Stefan Zerbe, Fakultät für Naturwissenschaften und Technik, Freie Universität Bozen-Bolzano, Universitätsplatz 5, I-39100 Bozen, Italien; E-Mail: stefan.zerbe@unibz.it ; Dr. Annett Frick, LUP GmbH, Potsdam, E-Mail: annett.fr
-
Abb. 1: Lage der Untersuchungsgebiete im Peene- und Trebeltal in Mecklenburg-Vorpommern. -
Abb. 2: Satellitenbild-gestützte Vegetationskarte des Polders Jargelin im Jahr 2007 (aus Steffenhagen et al. 2008a). -
Abb. 3: Flächenanteile der Vegetationstypen im Polder Jargelin im Jahr 2007 (vgl. Abb. 2) mit Flächenanteilen von Eleocharis palus tris = 0 %, Juncus effusus = 0,3 %, Lemnaceae = 1,0 %, Schwimmblattfluren = 0,05 %, offener Wasserfläche = 1,5 %, Sonstiges = 1,2 %. -
Abb. 4: a) Oberirdische Biomasse der untersuchten Helophyten und b) Gesamt-Biomasse der Schwebematten und Schwimmdecken, bezogen auf die Moorfläche. Box-plots mit unterschiedlichen Buchstaben zeigen signifikante Unterschiede, One-Way ANOVA, Post-Hoc-Scheffé-Test, p < 0,05, = Ausreißer und Extremwerte (aus Steffenhagen et al. 2008b). -
Abb. 5: Phosphor menge in der ober irdischen Biomasse zum maximalen Entwicklungszeitpunkt der Vegetation im Polder Jargelin 2007 (aus Steffenhagen et al. 2008c). -
Abb. 6: Satellitenbildgestützte Ableitung von Torfbildungs potenzialen auf der Grundlage der Wasserstufen und Vegetation im Polder Jargelin im Jahr 2007 (aus Steffenhagen et al. 2008c). -
Abb. 7: Prognose einer Sekundärsukzession für wiedervernässtes Moorgrünland mit permanent hoch überstauten Vernässungsflächen (Wasserstufe 6+ nach KOSKA 2001a; Grundwasser-Median 120-60 cm über Flur) 0-3, 3-10 und 20 Jahre nach der Wiedervernässung; die pflanzensoziologische Zuordnung erfolgte nach Berg et al. (2004), * Definition nach Wegener (1998). -
-
Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.