Nutzung von Windwurfflächen durch Vögel
Abstracts
Für eine natürliche Walderneuerung besitzen Windwurfflächen eine wichtige Bedeutung. Sie erhöhen die Strukturvielfalt und tragen damit zu einem größeren Artenreichtum bei. Die Nutzung von Windwürfen durch Vögel wird entscheidend durch die Faktoren Flächengröße, Vegetationsausprägung, Totholzanteil und Art der Umgebung beeinflusst.
Der Sturm Kyrill hinterließ im Januar 2007 einige Windwurfflächen im nordhessischen Nationalpark Kellerwald-Edersee. Während der Brutsaison 2008 wurden die Vogelgemeinschaften von neun Windwurfflächen untersucht.
Ziel der Studie war es heraus zu finden, wie Unterschiede in der Struktur, der Umgebung und der forstlichen Behandlung der Windwürfe die Nutzung durch Vögel beeinflussen.
Die meisten Reviere konnten in Bereichen mit dicht übereinander gestapeltem Totholz und vielen aufgeklappten Wurzeltellern festgestellt werden. Viele Reviere lagen allerdings auch im Grenzbereich zwischen Windwurf und Wald. Geräumte Windwürfe wiesen im Vergleich weniger Arten auf als ungeräumte. Auf Windwürfen mit angrenzendem Offenland waren die Vogelgesellschaften artenreicher als auf solchen, die komplett von Wald umgeben waren. Die Untersuchung zeigt, dass die strukturelle Vielfalt von ungeräumten Windwurfflächen im Zusammenhang mit einer diversen Umgebung verantwortlich für eine höhere Nutzung durch Vögel ist.
Influence of Different Structures of Windthrows on Bird Habitats Investigations in the National Park Kellerwald-Edersee
As an important effect of natural disturbances windthrows increase structural diversity and enhance biodiversity. Area size, vegetation structure, proportion of deadwood and environmental characteristics are decisive variables influencing the suitability of windthrows for birds.
In January 2007 the storm Kyrill caused several windthrows in the national park Kellerwald-Edersee in Northern Hesse. During the breeding season 2008 the bird communities of nine windthrows were investigated.
The aim of the study was to reveal how differences in structure, environment and forestal treatment of the windthrows affect their use by birds.
Areas of big stacked logs with large root plates sheltered the majority of bird territories on windthrows whereas cleared areas showed lower species richness and individual abundances. Furthermore a lot of territories were found in the transition of windthrow and forest. Windthrows near open areas had higher numbers of species. This study provides further evidence that the structural diversity of windthrows and their environment is one of the most significant reasons for the use of these areas by birds.
- Veröffentlicht am

1 Einleitung
Durch den Orkan Kyrill, der am 17. und 18. Januar 2007 über weite Teile Europas hinweg zog, wurden auch in Deutschland in vielen Gegenden großflächig Bäume entwurzelt. Im nordhessischen Nationalpark Kellerwald-Edersee entstanden vorwiegend in den zwischen Buchenwald eingestreuten Fichtenbeständen einige teils mehrere Hektar große Windwurfflächen.
Durch das Sturmereignis wurde die Waldstruktur punktuell deutlich verändert. Geschlossene Waldbestände wurden aufgelichtet und bereits vorhandene Offenlandbereiche weiter ausgedehnt.
Windwurfflächen unterscheiden sich hinsichtlich vieler ökologischer Faktoren von geschlossenen Waldbeständen. Es herrscht eine stärkere Windanfälligkeit und durch höhere Sonneneinstrahlung ist auch das Mikroklima deutlich verändert. Die Vegetationsstruktur ist durch liegendes und stehendes Totholz, welches vielen Tierarten günstige Brut- und Nahrungsbedingungen bietet, modifiziert. Zu den wichtigsten Folgen von Windwürfen zählt die Erhöhung der strukturellen Vielfalt und der damit einhergehende Anstieg der Artenvielfalt (u.a. Brawn et al. 2001, Fischer et al. 1998).
Das Zitat von Gatter (2004) Änderungen der Wälder verändern die Vogelwelt beschreibt die Bindung von Vogelarten und -gemeinschaften an strukturelle Eigenheiten von Baumbeständen (Scherzinger 2006) und verweist auf direkte Reaktionen der Arten auf Umweltveränderungen. Ergänzend bezeichnet Flade (1994) Vögel als schnelle Besiedler neu entstandener Lebensräume. Sie stellen demnach gute Indikatoren des Sukzessionsverlaufs auf Windwürfen dar und eignen sich zur Untersuchung der Nutzung von Windwurfflächen.
In 1989 beginnend untersuchte Scherzinger (2006) über einen Zeitraum von zwölf Jahren die Reaktion von Vogelgemeinschaften auf ein Sturmereignis des Jahres 1983. Dort gab es in den Folgejahren starke Borkenkäferkalamitäten. Nach den Stürmen Vivian und Wiebke in 1990 wurden in Baden-Württemberg ganze Waldbiozönosen von Windwurfflächen eingehend untersucht (Fischer 1998). Werth et al. (1998) stellten dabei Unterschiede der Avifauna von geräumten und ungeräumten Windwürfen heraus. Mit dem gleichen Ansatz arbeitete Hohlfeld (2006) bei seinen Untersuchungen von Bannwäldern ebenfalls in Baden-Württemberg nach dem Sturm Lothar in 1999. Auch in der Schweiz gab es Studien zu Vögeln auf Windwurfflächen (Glutz von Blotzheim 2001), insbesondere auch zum Einfluss forstlich unterschiedlich behandelter Flächen (Duelli et al. 2002).
Bis auf Leder et al. (2005) mit ihrer Beobachtung der Entwicklung von Vegetation und Tierwelt auf Windwurf-Dauerbeobachtungsflächen in Nordrhein-Westfalen finden sich in der Literatur keine Arbeiten, welche systematisch die Nutzung und Verteilung von Vögeln auf bestimmte Bereiche und Strukturen von Windwürfen untersuchen. Der Einfluss der umgebenden Landschaft findet in den meisten Studien keine Beachtung.
Die vorliegende Arbeit befasst sich daher mit folgenden Fragestellungen:
Welche Strukturen können auf den Windwurfflächen unterschieden werden? Sind Präferenzen bei der Nutzung durch die Avifauna zu erkennen?
Gibt es Unterschiede in der Zusammensetzung der Vogelgemeinschaften von Windwürfen, die von Offenland bzw. von Wald umgeben sind?
Unterscheiden sich die Vogelgemeinschaften von geräumten und nicht geräumten Windwurfflächen?
Waldbereiche, die wie die meisten der hier untersuchten Probeflächen zu 100 % dem Prozessschutz unterliegen, lassen sich in normal bewirtschafteten Wäldern nicht finden. Hier bietet der Nationalpark Kellerwald-Edersee die seltene Gelegenheit, die natürliche Entwicklung der Sukzession und der Artengemeinschaften von Windwürfen, auch in Zukunft, zu verfolgen.
2 Untersuchungsgebiet
2.1 Naturraum
Der im hessischen Landkreis Waldeck-Frankenberg gelegene Nationalpark Kellerwald-Edersee wurde am 01. Januar 2004 gegründet und erstreckt sich auf einer im Norden durch den Edersee begrenzten Fläche von 5724 ha (Frede 2007).
Das Landschaftsbild des Nationalparks ist geprägt durch einen Wechsel bewaldeter Mittelgebirgszüge mit teils tief eingeschnittenen Bachtälern. Dabei werden Höhen von 200 m bis knapp 680 m ü. NN erreicht. Tonschiefer und Grauwacke sind die vorherrschenden Gesteine. Die Böden sind flach- bis mittelgründig und von überwiegend saurer bis nährstoffarmer Ausprägung (Frede 2007).
Die Leelage östlich von Rothaargebirge und Westerwald bestimmt die klimatischen Verhältnisse. So werden im Bereich des subatlantisch-subkontinentalen Übergangsklima Jahresmitteltemperaturen von 6,5 °C bis 8,0 °C sowie Jahresniederschlagsmengen von 600 mm bis 800 mm erreicht (Becker et al. 1996).
Der nährstoffarme Hainsimsen-Buchenwald (Luzulo-Fagetum) dominiert das Waldbild. Neben einer bemerkenswerten Vielfalt an Begleit- und Sonderbiotopen stellen Altholzbereiche mit teils über 160 Jahre alten Rotbuchen (Fagus sylvatica) keine Seltenheit dar (Frede 2007).
2.2 Witterungsbedingungen
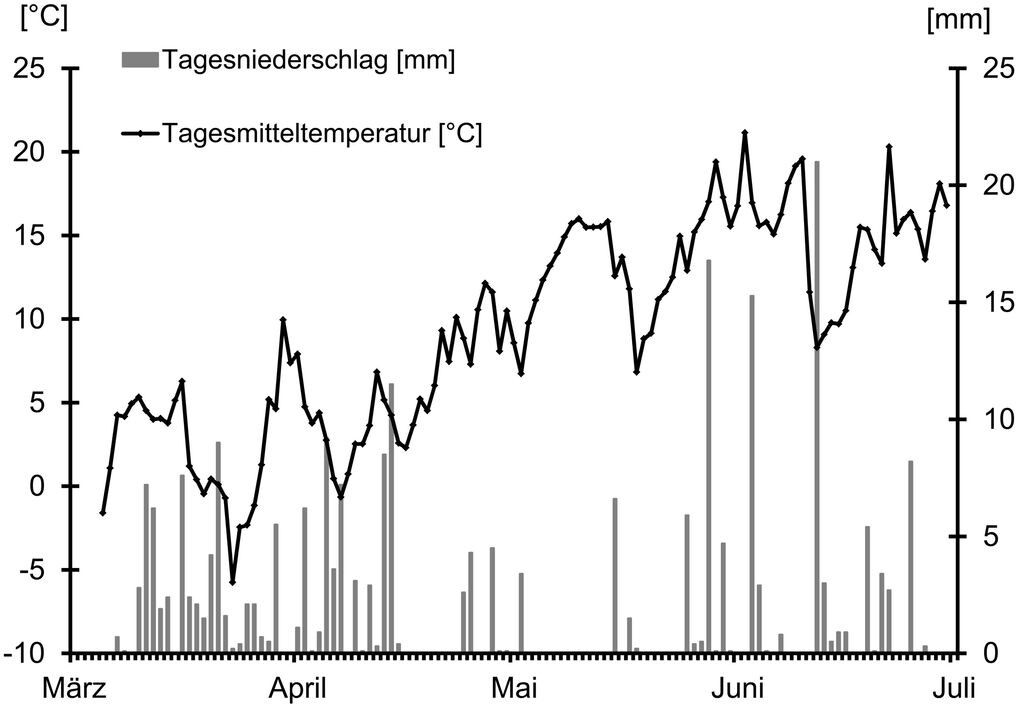
Die Witterungswerte (Abb. 1) während des Untersuchungszeitraumes wurden an der Messstation Kellerwald des Hessischen Landesamtes für Umwelt und Geologie (2008) erhoben. Diese liegt im Osten des Nationalparks auf 460 m ü. NN (Rechtswert: 3495010, Hochwert: 569 9619).
2.3 Untersuchungsflächen
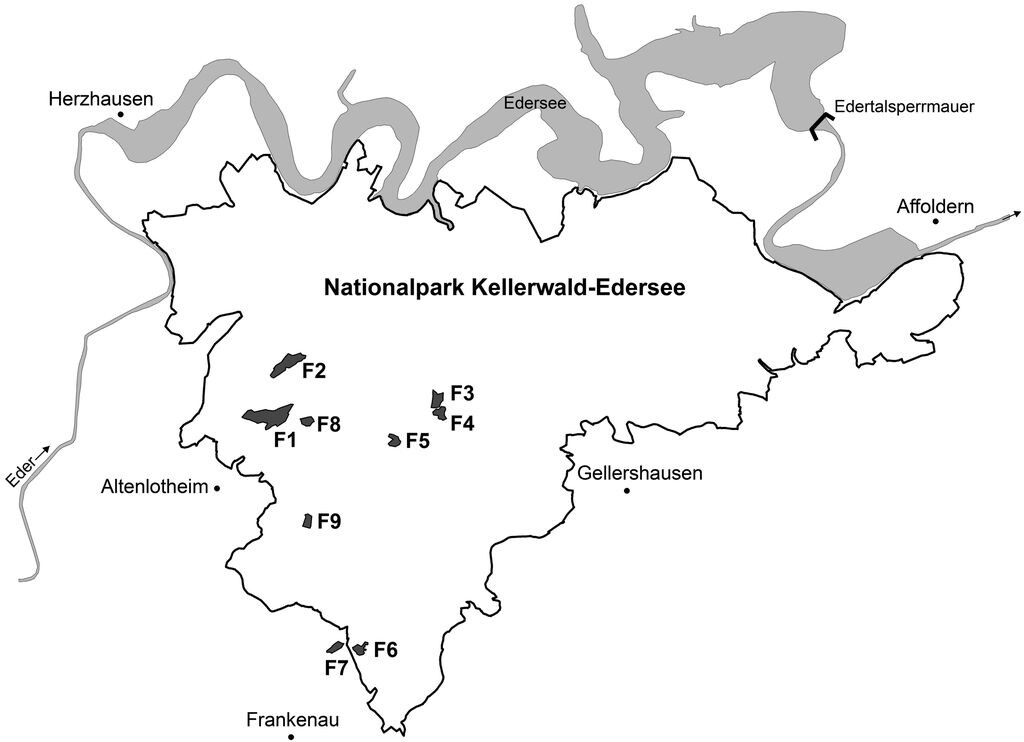
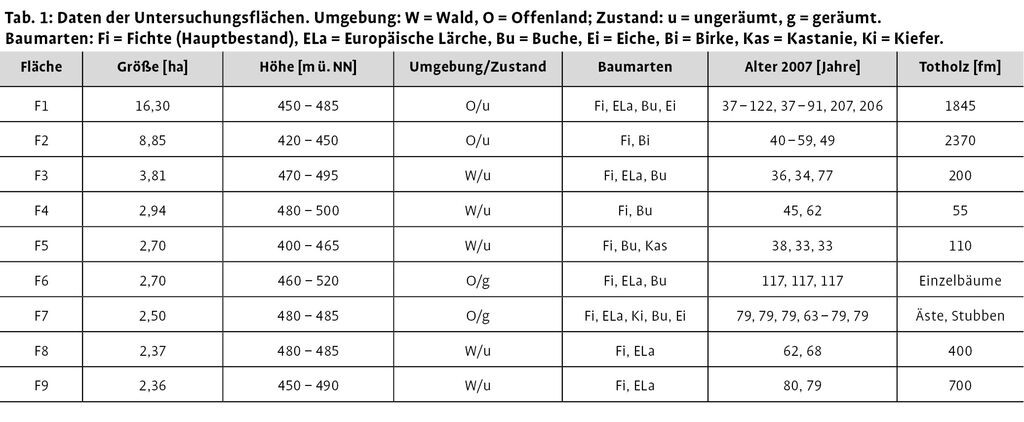
Die Untersuchungsflächen (Abb. 2 und 3) wurden anhand ihrer Größe (Mindestgröße 2 ha) sowie ihrer Erreichbarkeit ausgewählt. Entsprechend der Fragestellung sollten von Wald umgebene bzw. an Offenland angrenzende und geräumte sowie ungeräumte Windwürfe vertreten sein. Anhand dieser Kriterien standen neun Windwurfflächen mit einer Gesamtfläche von 44,5 ha zur Verfügung (Tab. 1).
Sofern es sich bei den Untersuchungsflächen um Waldflächen handelt, sind sie größtenteils von Hainsimsen-Buchenwald umgeben. Kleinräumig gibt es angrenzende Fichtenbereiche, die nicht vom Sturm umgeworfen wurden, sowie ehemalige Aufforstungsflächen oder Buchendickungen.
An die Windwürfe angrenzende Offenlandbereiche bestehen meist aus Rotschwingel- und Rotstraußgraswiesen (Festuca rubra-Agrostis tenuis-Gesellschaft) oder auch frischem Grünland, wie im Fall von F1 und F2. In naher Umgebung von F6 und F7 befinden sich, da sie direkt an der Nationalparkgrenze liegen, landwirtschaftliche Nutzflächen. Auch kleine Schlagflur- und Sukzessionsbereiche schließen direkt an die Windwürfe an.
Auf den Windwürfen selbst dominieren, besonders in den offeneren Bereichen, überwiegend Digitalis purpurea, Rubus fruticosus agg., Rubus ideaus und Deschampsia flexuosa.
In den Jahren vor Kyrill wurden aus einigen Beständen der Untersuchungsflächen teilweise gewisse Mengen von Fichten entnommen (mdl. Nationalparkförster R. Chartschenko).
3 Methoden
3.1 Vogelerfassung
Zur Erfassung der Vögel wurde die Revier- bzw. Probeflächenkartierung (Südbeck et al. 2005) angewandt. Im Zeitraum vom 05.03. bis 23.06.2008 wurde jede Windwurffläche insgesamt zehn Mal begangen. Grundsätzlich wurden alle Vogelarten kartiert, die während der Kontrollen auf oder in unmittelbarer Umgebung der Windwürfe akustisch oder visuell erfasst werden konnten. Aufgrund der Reviergröße von Eulen und tagaktiven Greifvögeln im Vergleich zu den größtenteils eher kleinen Windwürfen wurde auf deren Revierkartierung verzichtet. Tagaktive Nahrungsgäste wurden registriert.
Zur Festlegung von Revieren der Brutvögel wurden mindestens drei revieranzeigende Beobachtungen vorausgesetzt (Flade 1994, Südbeck et al. 2005).
3.2 Auswertungsmethoden
Um die Auswertung zur Windwurfstrukturnutzung zu vereinfachen, wurde bei den Vögeln zwischen Brutvogel, Nahrungsgast, Durchzügler und Überflieger unterschieden. Zur genaueren Analyse wurden die Brutvögel, je nach Lage ihrer Reviere in
Brutvögel mit mindestens einmal einem vollständigen Revier auf einem Windwurf (Bw),
Brutvögel mit Revieren auf der Windwurfgrenze und Revieren außerhalb (Br),
Brutvögel mit Revieren, die komplett außerhalb der Windwurfflächen liegen (Ba) und
Brutvögel mit großräumigeren Revieren, die die Windwürfe mit einschließen (Bg) unterteilt.
3.3 Gildeneinteilung
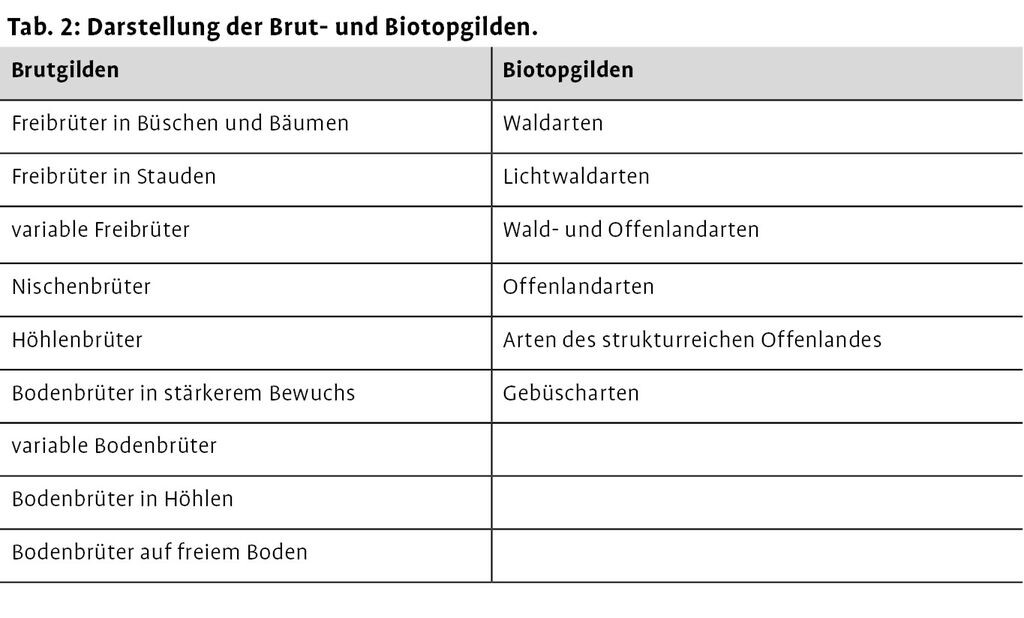
Zur Charakterisierung des Brutverhaltens und der Ansprüche an ihren bevorzugten Lebensraum wurden die auf den Windwürfen vorkommenden Vögel in entsprechende ökologische Gilden eingeteilt (Tab. 2).
3.4 Siedlungsdichte
Siedlungsdichtewerte wurden berechnet, um die Artenzusammensetzung und relativen Häufigkeiten der Vögel bzw. Arten einer Fläche bestimmen und mit anderen Flächen vergleichen zu können. Die Siedlungsdichte gibt die Reviere einer Brutvogelart oder einer Brutvogelgemeinschaft pro Hektar an. Bei der Auszählung wurden Reviere, welche sich über die Grenzen der Windwürfe hinaus erstrecken, nur zur Hälfte in Bezug auf die jeweilige Fläche gewertet. Südbeck et al. (2005) stellen dies für nicht projektgebundene Untersuchungen frei.
3.5 Strukturabgrenzung und Strukturdiversität
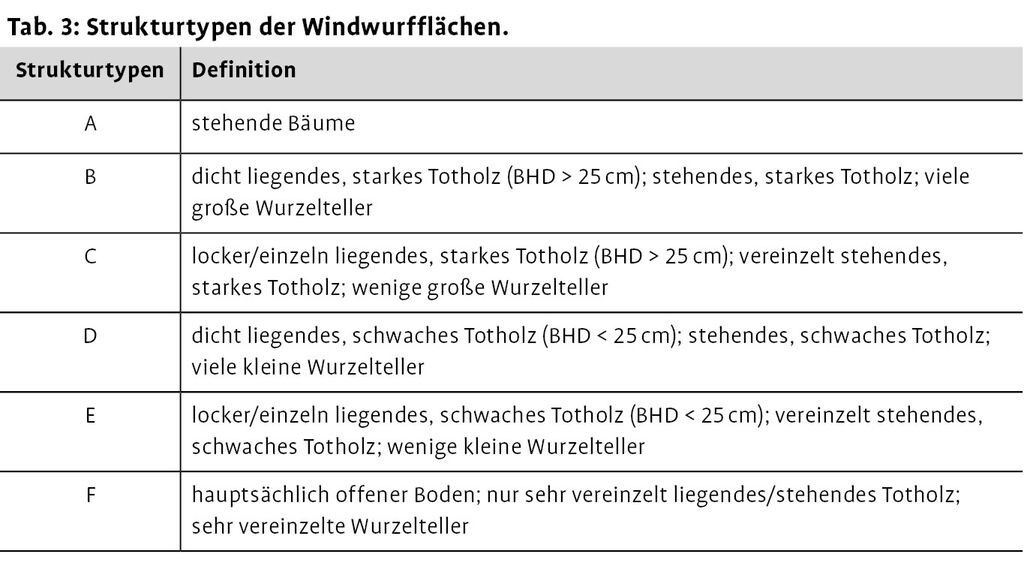
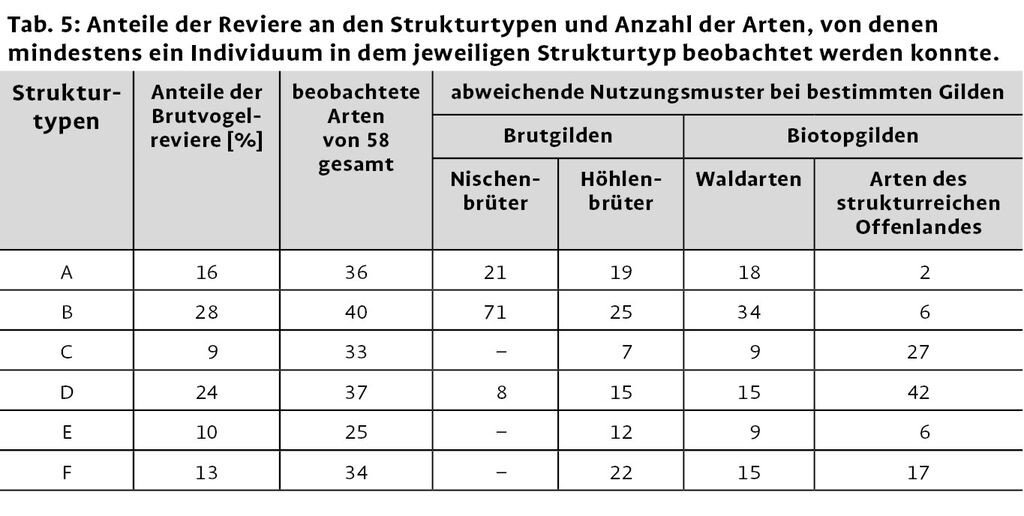
Die Struktur von Windwürfen ist z.T. schon auf kleinem Raum verschieden. Zur Klärung der Frage, inwieweit Vögel bestimmte Bereiche häufiger oder seltener nutzen, wurden die Untersuchungsflächen in Strukturtypen unterteilt (Tab. 3). Besondere Kriterien waren Größe und Dichte der gefallenen Bäume sowie Menge und Ausprägung von senkrecht aufgeklappten Wurzeltellern. Bereiche mit viel liegendem Totholz beinhalteten gleichzeitig auch einen großen Anteil an teils abgebrochenem, aber noch stehendem Totholz.
Im Feld waren die unterschiedlichen Strukturbereiche, aufgrund mangelnden Überblicks, kaum auszumachen, daher wurde auf digitale Luftbilder (Fa. Aerosense) aus dem Jahr 2007 zurückgegriffen.
Mithilfe des Shannon-Diversitätsindex wurde für jede Fläche die Strukturdiversität berechnet:

Je größer der Wert HS, desto höher die Strukturvielfalt einer Windwurffläche. S bezeichnet die Gesamtzahl der vorkommenden Strukturtypen. pi stellt den relativen Anteil des Vorkommens eines Strukturtyps (i) an der Gesamtzahl aller Strukturtypen der Fläche dar. Als di wird der absolute Anteil eines Strukturtyps (i) an der Gesamtzahl aller Strukturtypen bezeichnet. D steht für das gesamte Vorkommen aller Strukturtypen einer Fläche.
4 Ergebnisse
4.1 Windwurfstrukturen
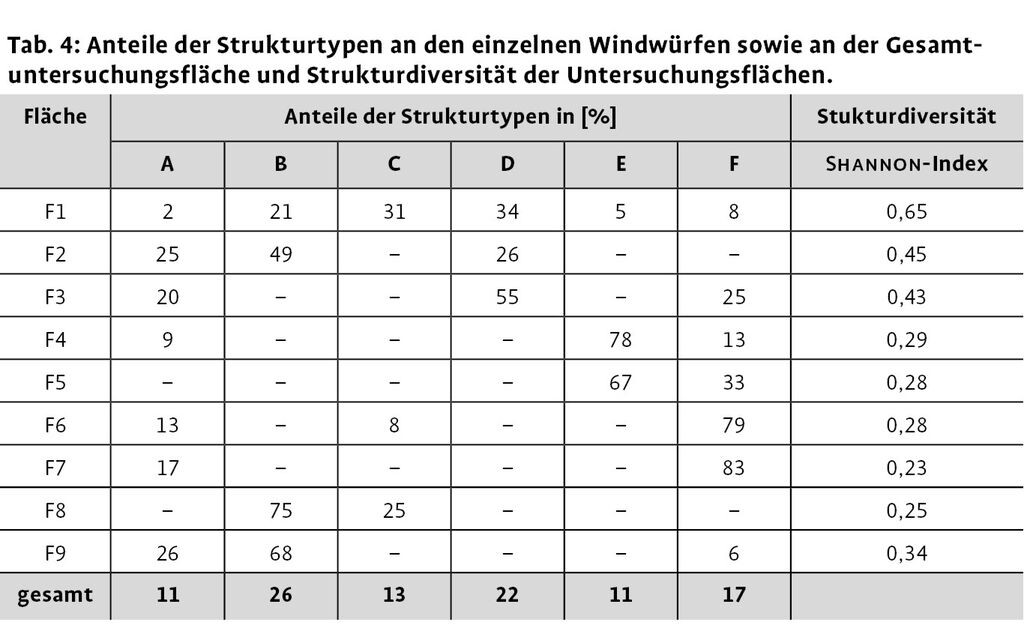
Die Anteile der verschiedenen Strukturtypen an den Untersuchungsflächen variieren relativ stark (Tab. 4). Die Strukturtypen B und D, welche durch Bereiche mit dicht liegendem Totholz gekennzeichnet sind, haben den größten Anteil an der Gesamtuntersuchungsfläche, kommen aber nur auf vier Probeflächen vor. Fläche F1 ist die einzige, auf der alle sechs Strukturtypen zusammen vorkommen, was sich auch in dem höchsten Wert der Strukturdiversität niederschlägt. Die beiden geräumten Windwürfe F6 und F7 weisen jeweils sehr hohe Anteile an offenen Bereichen (F) auf, was wiederum eine geringere Strukturdiversität zur Folge hat.
4.2 Strukturnutzung der Vögel
Bei der Untersuchung konnten 58 Vogelarten festgestellt werden. 37 Arten können als Brutvögel bezeichnet werden. Bei den restlichen 21 handelt es sich um Nahrungsgäste oder kurzzeitig rastende Durchzügler. 23 Arten der Brutvögel haben Reviere auf den Windwürfen (Bw), sieben in den Randbereichen (Br), fünf großräumige Reviere (Bg) und zwei nur außerhalb der Windwürfe (Ba). Von den 587 Revieren der Brutvögel Bw und Br lagen 154 komplett auf den Windwürfen, 197 außerhalb und 236 im Grenzbereich.
Als häufigste Vogelarten, nach Anzahl ihrer Reviere, können Buchfink (Fringilla coelebs), Zaunkönig (Troglodytes troglodytes), Kohlmeise (Parus major), Amsel (Turdus merula) und Zilpzalp (Phylloscopus collybita) genannt werden.
Die beiden Arten mit den häufigsten Revieren komplett auf den Windwurfflächen sind Zaunkönig und Heckenbraunelle (Prunella modularis). Sie weisen, auf die Gesamtuntersuchungsfläche bezogen, auch die höchsten Siedlungsdichtewerte mit je 9,0 Rev./10 ha bzw. 7,0 Rev./10 ha auf.
Die Windwurffläche F2 ist mit einem Wert von 91,5 Rev./10 ha am dichtesten besiedelt. F1, die größte Fläche, kommt lediglich auf 46,3 Rev./10 ha.
Die Strukturtypen mit dicht liegendem Totholz (B, D) scheinen insgesamt, wie auch zur Reviergründung für Brutvögel, am attraktivsten zu sein (Tab. 5). Die Typen mit locker liegendem Totholz (C, E) ziehen die Vögel am wenigsten an. Dieses allgemeine Nutzungsmuster lässt sich, bis auf wenige Ausnahmen, auch auf die Gilden übertragen. Nischenbrüter, zu denen nur der Grauschnäpper von F2 zählt, und Höhlenbrüter zeigen eine leicht veränderte Strukturnutzung. Für die Höhlenbrüter scheinen nach dem dichten Totholzbereich auch Bereiche mit stehenden Bäumen (A) und Offenbodenbereiche (F) von größerer Bedeutung zu sein. Ähnlich sieht das Bild bei den Waldarten der Biotopgilden aus. Hingegen scheinen die offeneren bzw. Bereiche mit schwächerem Totholz den Arten des strukturreichen Offenlandes eher zu liegen.
4.3 Unterschiede zwischen Offenland- und Waldwindwürfen
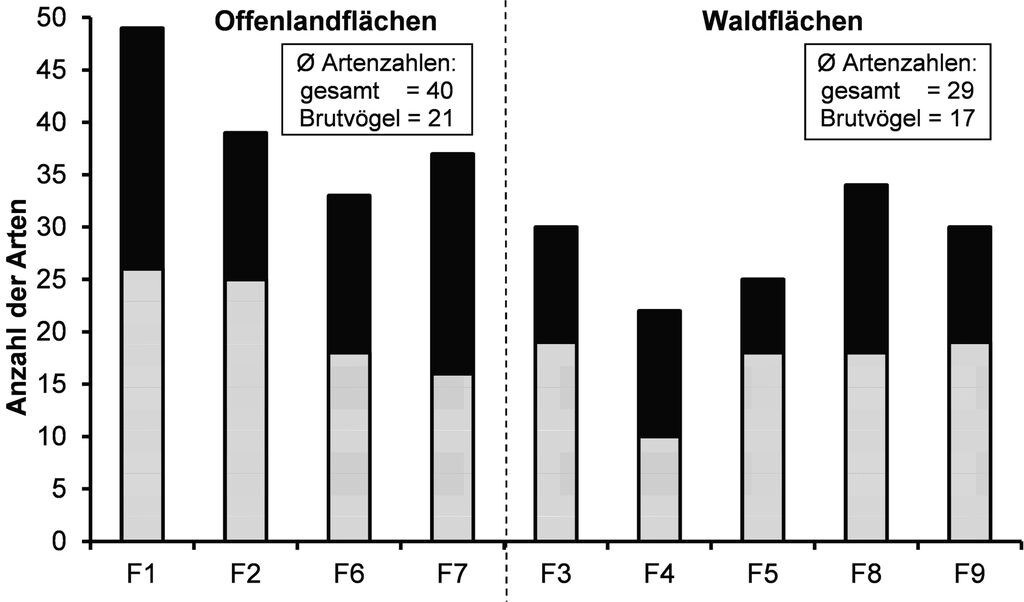
Im Unterschied zu den Waldwindwürfen zeichnen sich die Offenlandwindwürfe durch hohe Gesamtartenzahlen aus. Beim Vergleich der Brutvogelzahlen fällt dieser Unterschied allerdings nicht ganz so deutlich aus (Abb. 4). Während der zehn Begehungen konnten die Offenlandflächen durchgehend höhere Gesamt- wie auch Brutvogelartenzahlen aufweisen.
Auf den beiden großen Offenlandflächen F1 und F2 konnten insgesamt die meisten Vogelarten nachgewiesen werden (Abb. 4). Die geräumten Flächen F6 und F7 ähneln in der Anzahl der Brutvogelarten den Waldflächen.
Die Artenzusammensetzung, besonders auf die Biotopgilden bezogen, stellt ein weiteres Unterscheidungskriterium dar. Die Waldarten sind auf beiden Flächenkategorien am stärksten vertreten, in der weiteren Gegenüberstellung werden allerdings Unterschiede deutlich. An Offenland angepasste Arten treten auf den Offenlandwindwürfen auch stärker in Erscheinung, Gebüscharten sind hingegen seltener.
In der allgemeinen Zusammensetzung der Brutgilden sind die freibrütenden Arten auf den Offenlandflächen die dominierende Gruppe. Bei den Brutvögeln liegen sie allerdings auch hier, wie auf den Waldflächen, hinter den Höhlenbrütern.
4.4 Unterschiede zwischen geräumten und ungeräumten Windwürfen
Auf den geräumten Flächen (F6, F7) dominiert der Strukturtyp F (offener Boden) ganz deutlich (Tab. 4). Sie sind beide stark mit Brombeer- und Himbeerstauden bewachsen. Die Gesamtartenzahlen an beobachteten Vögeln sind hier relativ hoch, bei den Brutvogelarten allerdings eher niedrig. Als Brutvögel treten größtenteils Höhlenbrüter und Freibrüter in Büschen und Bäumen in Erscheinung. Waldvogel- und Gebüscharten haben den größten Anteil an den Biotopgilden.
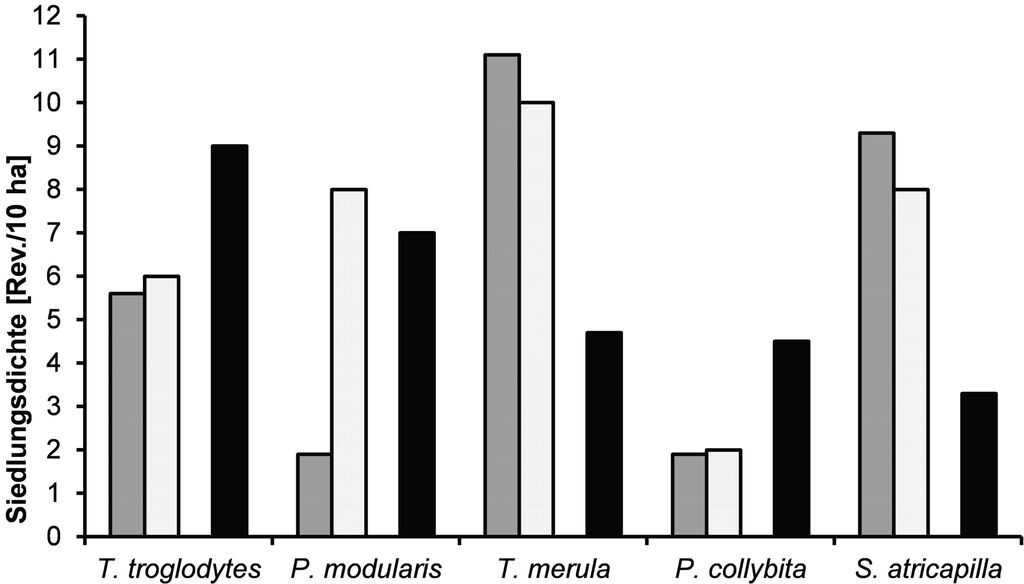
Einige Arten haben auf den geräumten Windwürfen im Vergleich zur Gesamtsiedlungsdichte relativ stark abweichende Werte (Abb. 5). Zaunkönig, Heckenbraunelle und Zilpzalp weisen auf diesen Flächen sehr geringe Siedlungsdichten auf. Bei Amsel und Mönchsgrasmücke (Sylvia atricapilla) ist genau das Gegenteil der Fall. Diese beiden Arten haben hier stark erhöhte Werte. Der Baumpieper (Anthus trivialis) kommt auf den geräumten Windwürfen nicht vor.
5 Diskussion
5.1 Windwurfstrukturen und ihre Nutzung durch Vögel
Die Windwurffläche F1 hat im Vergleich zu den anderen Untersuchungsflächen die höchste Strukturdiversität. Dass in ihrem Bereich auch die höchste Zahl an Vogelarten festgestellt wurde, hängt vermutlich überwiegend mit der Flächengröße zusammen. Eine hohe Strukturvielfalt hat aber auch, wie bereits von einigen Autoren festgestellt, immer eine höhere Artendiversität zur Folge (u. a. Brawn et al. 2001, Duelli et al. 2002, Fischer et al. 1998, Jedicke 1996).
Das Totholz scheint für die Nutzung der verschiedenen Windwurfbereiche durch die Vögel der entscheidende Faktor zu sein. Gatter (2004) schreibt dem Vorkommen von Totholz eine höhere Verfügbarkeit an Nahrung, neue Nischen sowie geringere interspezifische Konkurrenz zu. Auch Zillmann (2004) sieht insbesondere im liegenden Totholz eine wichtige Nahrungsressource für Vögel sowie eine hohe Schutzfunktion. Die Dichte scheint stärkeren Einfluss als der Stammdurchmesser des liegenden Totholzes zu haben, da die Strukturtypen B und D am häufigsten aufgesucht werden. Das dicht liegende Totholz bietet den Vögeln nicht nur beim Brüten, sondern auch bei der Nahrungssuche einen hohen Schutz vor Feinden. Je mehr liegendes Totholz vorhanden ist, desto höher ist auch die Anzahl der Wurzelteller sowie die Zahl der abgebrochenen stehenden Totholzstämme. Gerade für Arten wie die Heckenbraunelle, die Flade (1994) zu den Leitarten von Kahlschlägen zählt, oder auch den Zilpzalp ist nach Fuller (2000) eine dichte Ausprägung des Unterholzes auf Bestandeslücken wichtig. Die stärkere Nutzung des Typs B spricht allerdings für eine höhere Priorität des stärkeren Totholzes. Utschick (1991) stuft die Bedeutung von starkem Totholz für Vögel als zwei- bis dreimal wichtiger als die von schwachem Totholz ein. Der Zaunkönig, der gefallene Bäume und Wurzelteller als Bruthabitat bevorzugt, bestätigt dies mit seinen hohen Siedlungsdichtewerten, besonders auf den Flächen F2, F8 und F9. Die Wurzelteller des dickeren Totholzes sind viel größer als die des dünneren und bieten bessere Brutmöglichkeiten. Auch die Ausdehnung der liegenden Kronen ist bei den dicken Bäumen größer und dichter. Utschick (1991) stellt fest, dass die Bedeutung von starkem Totholz für die Bruthöhlennutzung allerdings eher gering ist. Gesunde starke Laubbäume werden häufiger genutzt. Daraus resultiert womöglich der im Vergleich zum allgemeinen Nutzungsmuster höhere Anteil der Reviere im Strukturtyp A, nach B und F. Insgesamt kann davon ausgegangen werden, dass die Strukturtypen B und D den Vögeln die größere Strukturvielfalt bieten.
Die Waldarten nutzen größtenteils auch den Strukturtyp B. Hingegen erklärt sich der höhere Anteil der Reviere der Arten strukturreichen Offenlandes in den eher offeneren Strukturen dadurch, dass diese Vögel größtenteils von Warten aus nach Insekten oder Kleintieren am Boden jagen. Sie benötigen demnach eine Kombination aus Sitzwarten und offenen Bodenstellen, die sie gerade in den Bereichen C und D sowie in Grenzbereichen zum Strukturtyp F vorfinden.
Die etwas stärkere Nutzung der Strukturtypen A und F gegenüber C und E erklärt sich möglicherweise gar nicht dadurch, dass sie den Vögeln große Vorteile bieten, sondern dass sie viel häufiger auf den Untersuchungsflächen vorkommen.
Für die Nutzung bestimmter Strukturen durch Vögel spielen neben der Flächengröße auch immer noch andere Faktoren wie Verfügbarkeit von Brutplätzen, Nahrung und Feinddruck eine entscheidende Rolle.
5.2 Unterschiede zwischen Offenland- und Waldwindwürfen
Die Dynamik und die Prozesse innerhalb von Lebensgemeinschaften einer bestimmten Fläche beruhen generell nicht nur auf den dort vorliegenden Begebenheiten, sondern werden im Wesentlichen auch von der umgebenden Landschaft beeinflusst und mitbestimmt (Hirao et al. 2008).
Der Grund für die insgesamt höheren Artenzahlen der Offenlandflächen ist die höhere Lebensraum- bzw. Strukturvielfalt, allein schon durch den Wechsel und Grenzbereich zwischen Wald und Offenland. Die bereits schon vorhandenen Offenlandbereiche werden durch die Windwurfflächen erweitert und durch das liegende und stehende Totholz strukturreicher. Die Waldwindwürfe sind vermutlich für typische Offenlandarten zu klein, um ihren gesamten Lebensraumansprüchen gerecht zu werden.
Mit höherer Lebensraumvielfalt geht auch eine höhere Nahrungsvielfalt einher. Die Offenlandbereiche dürften durch ein wärmeres Mikroklima, speziell höhere Sonneneinstrahlung, für viele Insektenarten ein geeignetes Habitat darstellen. Fuller (1995) bestätigt einen höheren Insekten- wie auch Fruchtreichtum in offenen Bereichen im Vergleich zu geschlossenem Bestand. In der Umgebung von F1 und F2 gibt es im Sommer viele blühende Pflanzen, die das Nahrungsangebot für Insekten und somit auch für Vögel erhöhen.
Die Begründung für die von Beginn der Untersuchung an auf den Offenlandflächen festgestellten höheren Artenzahlen könnte bei einem insgesamt etwas wärmeren Mikroklima liegen. Die Witterung war zu Beginn der Kartierung noch sehr unbeständig, mit häufigeren Niederschlägen, teils als Schnee, und mit relativ niedrigen Temperaturen. Für die Veränderung der Artenzusammensetzung auf Kahlschlägen ist im Vergleich zu der des restlichen Waldes das durch höhere Sonneneinstrahlung wärmere und trockenere Mikroklima verantwortlich (Gatter 2000). So sind die mittleren Luft- und Bodentemperaturen, gerade auf größeren Windwürfen im Vergleich zu kleineren beschatteten Waldwindwürfen, etwas höher (Sousa 1984). Hierdurch könnten sich im Bereich des Offenlandes schon früher als auf den Waldflächen erste Insekten aufhalten und den Vögeln als Nahrungsgrundlage dienen. Dies ist gerade zum Ende des Winters, der Jahreszeit mit den höchsten Verlusten (Fuller 1995), wenn viele der Nahrungsressourcen aufgebraucht sind, eine große Erleichterung. Die im Frühjahr höhere Anzahl von Waldvogelarten auf den Offenlandflächen lässt somit auf eine Verlagerung der Nahrungssuche ins Offenland schließen. Vögel, deren Lebensraum sich durch geschlossene Wälder oder Buschland auszeichnet, sind ohnehin, zumindest zur Brutzeit, größtenteils insektivor (Fuller 1995). Nach Gatter (2000) nimmt die Artenvielfalt allgemein, besonders aber die von Insekten und Reptilien und damit auch von Vögeln als deren Räuber, auf neu entstandenen, waldfreien Flächen zu.
Durch die größere Lebensraumvielfalt erhöht sich auch die Vielfalt innerhalb der Biotopgildengemeinschaft auf den Offenlandbereichen. Gatter (2000, 2004) schreibt, dass natürliche waldfreie Stellen, die es ohne Sturmereignisse in den letzten Jahren in Mitteleuropa gar nicht gäbe, die Vielfalt an Arten fördern und sich dadurch Vogelgemeinschaften durch Wandlungen in der Waldstruktur ändern könnten. Auch anthropogen verursachte Freiflächen wie Kahlschläge können dazu beitragen, dass Offenlandarten zeitweise in Wälder vordringen (Jedicke 1995, 1997a). Je strukturreicher ein Wald, desto höher ist die Zahl der dort vorkommenden Vogelarten (Fuller 1995, Jedicke 1997a).
Neben Aspekten wie Mikroklima, Vegetationsstruktur und Form hat die Größe von Störungsflächen ebenfalls Anteil daran, ob sich bestimmte Arten, hier Offenlandarten, ansiedeln oder nicht (Brawn et al. 2001, Hirao et al. 2008). Roth (1976) und Denslow (1985) ergänzen, dass die Dominanz der Waldarten auf kleineren Störungsflächen meist erhalten bleibt, die Artenvielfalt insgesamt, im Vergleich zu unzerstörtem Wald, dort jedoch auch erhöht wird. Das Nationalparkgebiet dürfte mit der Aufweitung von Offenflächen und mehr Strukturreichtum durch den Windwurf besonders für Offenlandarten noch attraktiver geworden sein.
Größere Prädatoren wie Turmfalke (Falco tinnunculus) und Mäusebussard (Buteo buteo) konnten bei der Ansitzjagd auf den großen Windwürfen beobachtet werden. Scherzinger (2006) merkt an, dass gute Versteckmöglichkeiten auf den Windwürfen das Populationswachstum von Nagern fördern. Die Flächen sind vom Nahrungsangebot her somit attraktiv für Kleinsäugerprädatoren, stellen aber auch kein einfaches Jagdgebiet dar.
Anhand des höheren Anteils der Höhlenbrüter in der Brutvogelgemeinschaft, zu denen über ein Drittel der Waldvogelarten gehören (Fuller 1995), erkennt man, dass diese auch auf den Offenlandflächen größtenteils von der umgebenen Waldlandschaft geprägt ist. Wie Jedicke (1997b) und Paleit (1998) feststellten, ist der Anteil von Höhlenbrütern in Buchenalthölzern höher als der von Freibrütern. Ohne den Einfluss des umgebenden Waldes würden die Freibrüter, wie bei der Gesamtvogelgemeinschaft der Offenlandflächen, vermutlich dominieren. Je nach Verbuschungsgrad würden sich verstärkt Arten wie Grasmücken einfinden (Fuller 1995).
5.3 Unterschiede zwischen geräumten und nicht geräumten Windwurfflächen
Die Gesamtartenzahlen der beiden geräumten Windwürfe (F6, F7) sind zwar im Vergleich relativ hoch, während die Anzahl der Brutvogelarten, gerade auf den Windwürfen selbst, nicht höher als die der nicht geräumten Flächen ist.
Werth et al. (1998) stellten bei einem Vergleich auf ungeräumten Windwürfen sogar nahezu doppelt so viele Brutvogelarten fest wie auf geräumten.
Aufgrund der Tatsache, dass es sich bei den geräumten Flächen um Offenlandflächen handelt und sie recht hohe Gesamtartenzahlen aufweisen, könnte man erwarten, dass die Zahl der Brutvögel im Vergleich auch etwas höher ist als auf den Waldflächen. Das wäre wohl auch der Fall, wenn das Totholz hier liegengeblieben wäre und so die Strukturvielfalt der Flächen erhöht hätte.
Die einzigen Strukturen auf F6 und F7 sind einzeln stehende Bäume und Bereiche, in denen Brombeer- oder Himbeerstauden wachsen, ansonsten herrscht mit Gräsern bewachsener, offener Boden vor, wo aufgrund fehlenden Schutzes vor Räubern kaum ein Vogel brüten dürfte.
Auf Dauerbeobachtungsflächen im Nationalpark Bayerischer Wald wurde fünf Jahre nach einem Sturmereignis immer noch eine Dominanz von Himbeersträuchern auf geräumten Flächen festgestellt (Fischer et al. 1990). Schlagflurarten waren in der Vegetation dieser Windwürfe stark vertreten, während auf ungeräumten Flächen fast ausschließlich Waldpflanzen wuchsen. Auch hier wurde festgestellt, dass geräumte Windwürfe weniger strukturelle Vielfalt aufweisen, weil u.a. die senkrecht stehenden Wurzelteller, von Fischer et al. (1998) sowie Scherzinger & Schumacher (2004) als wertvolle Sonderstandorte bezeichnet, zurückgeklappt wurden (Fischer et al. 1990).
Die Abwesenheit des Baumpiepers auf den beiden Flächen könnte für die fehlenden Strukturen beispielhaft sein. Er benötigt hohe Singwarten sowie eine gut entwickelte und vielfältig strukturierte Krautschicht. Zaunkönig, Heckenbraunelle und Zilpzalp zeigen hier ebenfalls aufgrund ihrer niedrigen Siedlungsdichten die fehlenden Strukturen an. Gerade für Arten wie u.a. Zaunkönig und Heckenbraunelle sind senkrecht stehende Wurzelteller von großer Bedeutung, da sie als Singwarte, Nistplatz, Nahrungsraum oder auch Versteck dienen (Scherzinger & Schumacher 2004).
Klare Profiteure dieser Flächenausprägung scheinen jedoch Mönchsgrasmücke und Amsel zu sein, die hier auf beiden geräumten Windwürfen bei weitem ihre höchsten Siedlungsdichtewerte erreichen. Die Amsel als variabler Freibrüter, durchaus auch in Gebüschen bodennah brütend, und die Mönchsgrasmücke als Freibrüter in Stauden dürften von den Brombeer- und Himbeersträuchern und Gebüschen im Randbereich als Brutplätze profitieren. Sie bieten ihnen während der Brut Schutz vor Prädatoren. Außerdem dürften deren Beeren später im Jahr als Nahrungsquelle eine wichtige Rolle spielen.
Der hohe Anteil der Gebüscharten bei den Biotopgilden der Brutvogelgemeinschaft zeigt, dass sich die geräumten Flächen am Waldrand befinden. Dort sind Sukzessionsflächen und Schlagflurbereiche mit jüngeren Bäumen oder niedrigeren Büschen in unmittelbarer Umgebung lokalisiert. Der Wald scheint hier nicht mehr so großen Einfluss auf die Brutvogelgemeinschaft zu haben, so dass der Anteil der Waldarten geringer ist. Auch auf den beiden Waldwindwürfen F4 und F5, mit strukturärmerer Ausprägung, haben die Gebüscharten einen höheren Anteil. Die Waldarten scheinen allgemein die strukturreicheren Windwürfe zu bevorzugen.
Zu vermuten ist, dass geräumte Flächen auch eine geringere Nahrungsgrundlage bieten als ungeräumte. Totholzinsekten dürften hier kaum vorkommen. Borkenkäfer finden sich auf nicht geräumten Flächen in viel größerer Anzahl als auf geräumten und haben bei Untersuchungen von Wermelinger et al. (1995) zwei Jahre nach dem Sturmereignis ihren höchsten Populationsstand erreicht. Duelli & Obrist (1999) konnten beim Vergleich der Arthropodenfauna von geräumten und ungeräumten Windwürfen in der Schweiz allerdings kaum Unterschiede in der Artenzahl ausmachen. Jedoch war die Artenzusammensetzung auf den beiden Flächentypen vollkommen unterschiedlich. Dagegen vermuten Werth et al. (1998), dass Arthropoden auf nicht geräumten Flächen viel häufiger sind als auf geräumten, da sie auf den nicht geräumten Windwürfen fast das Zweifache an Vögeln als Nahrungsgäste beobachten konnten. Eine Erklärung hierfür liefern die durch das liegen gebliebene Totholz vielfältigeren Strukturen.
Bodenvögel, die man vielleicht aufgrund der Offenbodenbereiche erwarten würde, sind auf geräumten Windwürfen nicht häufiger anzutreffen als auf ungeräumten. Das Nahrungsangebot scheint daher auf beiden Flächentypen nicht schlecht zu sein. Nach Wermelinger et al. (1995) ist das Vorkommen unterschiedlicher Tiergruppen von der Art des Flächentyps abhängig. Demnach fanden sie bei ihrer Untersuchung heraus, dass Totholzinsekten auf den nicht geräumten Windwürfen am häufigsten sind. Spinnen, Blüten besuchende Insekten und Eidechsen traten dagegen auf den geräumten Windwürfen häufiger auf. Bei Schmetterlingen wurde kein Unterschied festgestellt. Ausschlaggebend hierfür dürften die Vegetationszusammensetzung sowie das unterschiedliche Mikroklima sein. Auf den geräumten Windwürfen können sich Blütenpflanzen viel besser ausbreiten und der offene Boden erwärmt sich durch direkte Sonneneinstrahlung besser, was den Reptilien zugute kommt. Für Ansitzjäger auf Reptilien, wie z.B. den Raubwürger (Lanius excubitor), fehlen hier allerdings wiederum die Sitzwarten, so dass die Vögel davon nicht unbedingt profitieren können.
Literatur
Becker, W., Frede, A., Lehmann, W. (1996): Pflanzenwelt zwischen Eder und Diemel. Flora des Landkreises Waldeck-Frankenberg mit Verbreitungsatlas. Naturschutz in Waldeck-Frankenberg 5. Korbach.
Brawn, J.D., Robinson, S.K., Thompson III, F.R. (2001): The role of disturbance in the ecology and conservation of birds. Annual Review of Ecology and Systematics 32, 251-276.
Denslow, J.S. (1985): Disturbance-mediated coexistence of species. In: Pickett, S.T.A., White, P.S., Hrsg., The ecology of natural disturbance and patch dynamics, New York, 307-323.
Duelli, P., Obrist, M.K. (1999): Räumen oder belassen? Die Entwicklung der faunistischen Biodiversität auf Windwurfflächen im schweizerischen Alpenraum. Verh.Ges. Ökol. 29, 193-199.
, Obrist, M.K., Wermelinger, B. (2002): Windthrow-induced changes in faunistic biodiversity in alpine spruce forests. Forest, Snow and Landscape Research 77 (1/2), 117-131.
Fischer, A. (Hrsg., 1998): Die Entwicklung von Wald-Biozönosen nach Sturmwurf. Ecomed, Landsberg.
, Abs, G., Lenz, F. (1990): Natürliche Entwicklung von Waldbeständen nach Windwurf. Ansätze einer Urwaldforschung in der Bundesrepublik. Forstwiss. Cbl. 109, 309-326.
, Honold, A., Reif, A., Funke, W., Bücking, W. (1998): Die Sturmwurf-Biozönosen in der Startphase der Bestandesentwicklung. In: Fischer, A., Hrsg., Die Entwicklung von Wald-Biozönosen nach Sturmwurf, Ecomed, Landsberg, 357-374.
Flade, M. (1994): Die Brutvogelgemeinschaften Mittel- und Norddeutschlands. Grundlagen für den Gebrauch vogelkundlicher Daten in der Landschaftsplanung. IHW, Eching.
Frede, A. (2007): Nationalpark Kellerwald-Edersee. Der Nationalpark Kellerwald-Edersee ein Buchenwald-Naturerbe von europäischem Rang. In: Nordhessische Gesellschaft für Naturkunde und Naturwissenschaften e.V. (NGNN), Hrsg., Naturschutzgebiete in Hessen. Bd. 4, Landkreis Waldeck-Frankenberg mit Nationalpark Kellerwald-Edersee, Cognitio, Niedenstein, 72-89.
Fuller, R.J. (1995): Bird life of woodland and forest. Cambridge University Press, Cambridge.
(2000): Influence of treefall gaps on distributions of breeding birds within interior old-growth stands in Biaowieza Forest, Poland. The Condor 102, 267-274.
Gatter, W. (2000): Vogelzug und Vogelbestände in Mitteleuropa. 30 Jahre Beobachtung des Tagzugs am Randecker Maar. Aula, Wiebelsheim.
(2004): Deutschlands Wälder und ihre Vogelgesellschaften im Rahmen von Gesellschaftswandel und Umwelteinflüssen. Vogelwelt 125 (3-4), 151-176.
Glutz von Blotzheim, U.N. (2001): Zur Entwicklung der Avifauna auf ehemaligen Sturmwurfflächen im Tannen-Buchenwaldareal am Schwyzer Nordalpenrand (1990-2000). Orn. Beob. 98, 81-112.
Hessisches Landesamt für Umwelt und Geologie (2008): Witterungsdaten der Messstation Kellerwald vom 01.03.2008 bis 01.07.2008. http://www.hlug.de/medien/luft/recherche/recherche.php?station=1350 (Stand: 13.03.2009).
Hirao, T., Murakami, M., Oguma, H. (2008): Functional spatial scale of community composition change in response to windthrow disturbance in a deciduous temperate forest. Ecol. Research 23 (2), 249-258.
Hohlfeld, F. (2006): Ornithologische Untersuchungen in einem Bann- und Wirtschaftswald vor und nach Sturm. Vogelwelt 127, 51-64.
Jedicke, E. (1995): Grenzstrukturen in Wäldern und ihr Einfluß auf die Avifauna. Ber. ANL 19, 115-123.
(1996): Brutvogelgemeinschaften in Buchen-Althölzern und -Schirmbestand im Krofdorfer Forst bei Gießen. Forstwiss. Cbl. 115, 163-173.
(1997a): Biotopverbund im Wald: Lässt sich dieses Naturschutz-Konzept von der freien Landschaft übertragen? Natur- und Kulturlandschaft 2, 90-96.
(1997b): Buchen-Altholzinseln als Naturschutz-Instrument im Wald. Vogel und Umwelt 9, 93-117.
Leder, B., Lehmann, A., Leonhardt, A. (2005): Vegetationsentwicklung und Avifauna auf Windwurfflächen. LÖBF-Mitt. 3/05, 39-42.
Paleit, J. (1998): Naturschutzfachlicher Beitrag zum Flächenmanagement im geplanten Nationalpark Kellerwald mit Hilfe der Avifauna. Unveröff. Dipl.-Arb. im Fachbereich Landschaftsarchitektur und Umweltplanung der Universi-tät Gesamthochschule Paderborn, Abteilung Höxter.
Scherzinger, W. (2006): Reaktionen der Vogelwelt auf den großflächigen Bestandeszusammenbruch des montanen Nadelwaldes im Inneren Bayerischen Wald. Vogelwelt 127 (4), 209-263.
, Schumacher, H. (2004): Der Einfluss forstlicher Bewirtschaftungsmaßnahmen auf die Waldvogelwelt eine Übersicht. Vogelwelt 125 (3-4), 215 - 250.
Sousa, W.P. (1984): The role of disturbance in natural communities. Annual Review of Ecology and Systematics 15, 353-391.
Südbeck, P., Andretzke, H., Fischer, S., Gedeon, K., Schikore, T., Schröder, K., Sudfeldt, C. (Hrsg., 2005): Methodenstandards zur Erfassung der Brutvögel Deutschlands. Radolfzell.
Utschick, H. (1991): Beziehungen zwischen Totholzreichtum und Vogelwelt in Wirtschaftswäldern. Forstwiss. Cbl. 110, 135-148.
Wermelinger, B., Duelli, P., Obrist, M., Odermatt, O., Seifert, M. (1995): Die Entwicklung der Fauna auf Windwurfflächen mit und ohne Holzräumung. Schweiz. Z. f. Forstwesen 146 (11), 913-928.
Werth, H., Werth, S., Funke, W. (1998): Vögel. In: Fischer, A., Hrsg., Die Entwicklung von Wald-Biozönosen nach Sturmwurf, Ecomed, Landsberg, 321-329.
Zillmann, F. (2004): Untersuchungen zur Struktur- und Habitatnutzung durch Vögel in Buchenwäldern. Unveröff. Dipl.-Arb. am Institut für Biologie, Humboldt-Universität Berlin.
Anschrift des Verfassers: Daniel Seitz, Im Ortsgrund 51, D-35066 Frankenberg (Eder), E-Mail [email protected] .
-
Abb. 1: Witterungsverlauf während des Untersuchungszeitraums. -
Abb. 2: Überblick über die Lage der Untersuchungsflächen im Nationalpark Kellerwald-Edersee. -
Abb. 3: Blick in die Untersuchungsfläche F1. -
Abb. 4: Vergleich der Gesamt- und Brutvogelartenzahlen zwischen Offenland- und Waldwindwürfen. Grau = Anzahl der Brutvogelarten, schwarz = Gesamtartenzahlen. -
Abb. 5: Siedlungsdichten ausgewählter Arten. Grau = Untersuchungsfläche F6, weiß = Untersuchungsfläche F7, schwarz = Sieldungsdichte bezogen auf die Gesamtuntersuchungsfläche. -
-
-
-
-
-
-


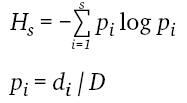
Bedienhilfen
Barrierefreiheit
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.