Raumnutzung des Libellen-Schmetterlingshafts nördlich der Alpen
Abstracts
Da die Verinselung von Lebensräumen nach wie vor zu den Hauptbedrohungen der Biodiversität gehört, ist es für den Naturschutz von zentraler Bedeutung, Verbundsysteme zu entwickeln, die den Austausch zwischen isolierten Habitaten ermöglichen.
Der Libellen-Schmetterlingshaft (Libelloides coccajus) ist nördlich der Alpen durch seine enge Bindung an Trockenstandorte als Eiablagehabitat stark von der Verinselung betroffen. Der Beitrag untersucht die Raumnutzung dieser geschützten Art, um aufzuzeigen, wie sie sich im Raum bewegt, ob Ausbreitungshindernisse bestehen und welche Habitattypen eine Ausbreitung zulassen.
In bestehenden Populationen sind windgeschützte Offenlandflächen wichtig für die Lebensraumqualität, da sie bevorzugt als Jagd- und Reproduktionsräume genutzt werden. Dabei spielt der Vegetationstyp eine eher untergeordnete Rolle. Die Art bewegt sich auch über Offenlandflächen mit intensiver landwirtschaftlicher Nutzung. Diese scheinen ebenso wenig wie Strauchhecken gravierende Ausbreitungshindernisse darzustellen.
Diese Resultate liefern artspezifische Grundlagen für eine praxisorientierte Planung von Artenschutz- bzw. Verbundprojekten.
Spatial Use of the Owly Sulphur north of the Alps Consequences for species protection
Isolation of habitats remains one of the most severe threats to biodiversity, and it is vital to enable exchange among habitat islands by creating suitable stepping stones or connecting habitat corridors.
The Owly Sulfur Libelloides coccajus has been heavily affected by isolation north of the Alps since it depends on dry and warm open forests and on grasslands which should not be cut before early August. The study investigated flying habits of this protected species in order to elucidate both its core habitat and adjacent areas used for hunting and mating, with special regard to obstacles.
The results revealed that wind protection seems to be one of the prime requirements for L. coccajus, whereas land use and thus vegetation type play a minor role, at least as far as hunting is concerned. For instance, L. coccajus was frequently observed to fly and hunt above corn fields and other crops. Apparently, neither hedge rows nor crop fields seem to seriously hamper the dispersal of L. coccajus.
This finding is very important for designing and creating effective stepping stones and connecting corridors between the remaining L. coccajus populations.
- Veröffentlicht am

1 Einleitung
Durch die Intensivierung der Landwirtschaft wurden viele Lebensräume zerschnitten und in ihrem Wert vermindert. Diese Entwicklung führte zu vielfältigen negativen Effekten auf die Biodiversität (z.B. Settele et al. 1996). Die Verinselung von Lebensräumen ist trotz verschiedener Bemühungen des Naturschutzes noch immer eine der Hauptursachen für den Rückgang der Artenvielfalt (z.B. Baier et al. 2006). Dementsprechend steht in vielen Ländern der Gedanke, isolierte Biotope beispielsweise über Biotopverbundsysteme miteinander zu verbinden, im Zentrum der Naturschutzbemühungen (Jedicke 1994, Kissling 2011).
Um einen effizienten Beitrag zum Arten- bzw. Biodiversitätsschutz zu leisten, sollte ein Verbundsystem den Ansprüchen verschiedener Arten gerecht werden, welche jedoch teilweise sehr unterschiedlich sind. Was den einen Arten die Ausbreitung erleichtert, kann für andere Arten ein unüberwindbares Hindernis darstellen (Dover & Settele 2009). Diese teilweise artspezifischen Eigenschaften sind für verschiedene Arten bereits gut dokumentiert (Tagfalter: Settele & Reinhardt 1999; Amphibien: Günther 1999), während sie für viele andere Tierarten noch weitgehend unbekannt sind. Aus diesem Grund ist es insbesondere für stark von der Verinselung bedrohte Arten wichtig zu klären, wie sich diese Arten im Raum bewegen und welche Habitate für eine Ausbreitung geeignet sind.
Das Eiablagehabitat des Libellen-Schmetterlingshafts (Libelloides coccajus [Denis & Schiffermüller 1775]), magere Halbtrocken- und Trockenwiesen (Fetz 2002 & Wolf 2005), wurde in den letzten ca. 100 Jahren flächenmäßig massiv verringert und die bis heute übrig gebliebenen Flächen sind meist stark isoliert und in ihrer Qualität vermindert (Lachat et al. 2010). Dadurch kommt die Art derzeit nördlich der Alpen meist nur noch in räumlich isolierten Populationen vor (Fetz 2002, Müller et al. 2012, Wolf 2005) und ist in der Schweiz und in Süddeutschland gefährdet (Schweiz: Duelli 1994, Baden-Württemberg: Tröger 1998, Bayern: Pröse & Gruppe 2003).
Welche Lebensräume die Art als Fluggebiet nutzt, ist derzeit noch nicht restlos geklärt. Fetz (2002) beschreibt die Art als wenig ausbreitungsfähig und Wolf (2005) berichtet, dass L. coccajus nur über offenem Gelände fliegt und keine hochbewachsenen Lesesteinriegel oder Waldstücke überquert. Diese Aussagen widersprechen zumindest teilweise den Beobachtungen von Fischer (2009) und Müller et al. (2012), die fliegende Individuen von L. coccajus unter anderem auch in einem lichten Föhrenwald nachgewiesen haben. Diese Diskrepanzen verdeutlichen, dass der Wissensstand über die Raumnutzung von L. coccajus noch sehr gering ist und somit die Wirkungen von Biotopverbundprojekten auf diese Art kaum realistisch abgeschätzt werden können.
Im Rahmen dieser Untersuchung wurden folgende Fragenstellungen bearbeitet:
1. Welche verschiedenen Habitattypen werden von L. coccajus als Fluggebiet genutzt?
2. Unterscheidet sich die Intensität verschiedener von L. coccajus genutzter Habitate und, falls ja, gibt es einen Zusammenhang zu bestimmten Standortfaktoren (z.B. Vegetationstyp, Wind, Beuteangebot)?
3. Wie mobil ist die Art in den untersuchten Gebieten? Gibt es Landschaftselemente, welche die Mobilität einschränken (z.B. intensiv bewirtschaftetes Grünland, Ackerland, Hecken)?
Diese Informationen können dabei helfen, artspezifische Maßnahmen zur Aufwertung des Lebensraumes zu formulieren, bestehende Biotopverbundsysteme bezüglich ihrer Auswirkungen auf L. coccajus zu überprüfen und gegebenenfalls aufzuzeigen, wie diese hinsichtlich der Förderung der Art optimiert werden können.
2 Material und Methoden
2.1 Artenportrait
Der Libellen-Schmetterlingshaft (Libelloides coccajus) ist ein mittelgroßer Netzflügler mit einer Spannweite von 42 bis 45 mm. Er weist auf den Vorder- und Hinterflügeln ausgeprägte gelbe Flecken auf. Von anderen Libelloides-Arten lässt er sich durch den schwarzen, bis zum Analwinkel reichenden Fleck der Hinterflügel (vgl. Abb. 1) eindeutig unterscheiden (Aspöck et al. 1980). Die Imagines ernähren sich räuberisch von kleinen fliegenden Insekten (Aspöck et al. 1980), die sie im Flug erbeuten.
Nördlich der Alpen kommt die Art nur in klimatischen Gunstlagen vor und steigt bis auf eine Höhe von rund 1200 m üb. NN (Gonseth 1991), während für das Wallis eine vertikale Ausbreitung bis 2200 m üb. NN erwähnt wird (Aspöck et al. 1980). Viele Autoren vermuten für L. coccajus eine zweijährige Entwicklungsdauer der Larven (Aspöck et al. 1980, Fetz 2002, Gonseth 1991, Wolf 2005), diese konnte aber noch nie bestätigt werden. Gesicherte artspezifische Angaben zum Larvalhabitat von L. coccajus fehlen bislang.
Die Eigelege werden in einer Doppelreihe rund 20cm über der Bodenoberfläche an lebende oder abgestorbene Pflanzenteile abgelegt (Fetz 2002, Müller et al. 2012a).
2.2 Untersuchungsgebiete
Die Untersuchungen wurden an vier Populationen (Glattfelden = GL, Merishausen = ME, Hessenberg = HE, Rheinheim = RE) in einem Umkreis von rund 25km durchgeführt, von denen drei in der Nordschweiz und eine in Süddeutschland liegen. In Abb. 2 sind die Untersuchungsgebiete mit den wichtigsten Habitattypen dargestellt. Die Untersuchungsgebiete wurden basierend auf einer ersten informellen Begehung so ausgeschieden, dass das gesamte zu diesem Zeitpunkt genutzte Fluggebiet großzügig vom Untersuchungsgebiet eingeschlossen wurde. Zum Untersuchungsgebiet GL ist anzumerken, dass die Parzelle 3.2 größer ist als auf dem Luftbild dargestellt, die angedeutete Zweiteilung der Parzelle war zum Untersuchungszeitpunkt nicht vorhanden. Die zur Eiablage genutzten Habitattypen jedes Untersuchungsgebietes sind in der Legende von Abb. 2 fett markiert.
2.3 Fluggebiet
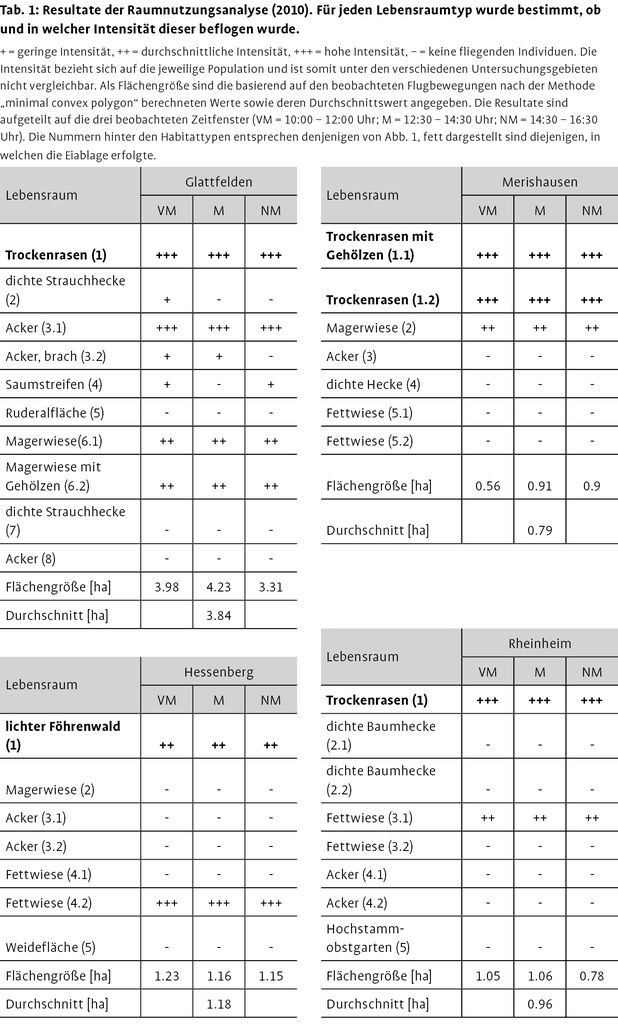
Zur Erfassung des genutzten Fluggebiets in den Untersuchungsgebieten wurde bei optimalen Flugbedingungen (sonnig, wenig Wind) in drei Zeitfenstern (VM = 10:00 12:00 Uhr; M = 12:30 14:30 Uhr; NM = 14:30 16:30 Uhr) jeweils das gesamte Untersuchungsgebiet nach fliegenden Individuen abgesucht; die beobachteten Flugbewegungen wurden auf einem Plan im Maßstab 1:1000 festgehalten. Diese Untersuchungen fanden an folgenden Tagen statt: GL: 23.05.2010, ME 24.05.2010, HE 05.06.2010, RH 07.06.2010. Alle in Abb. 2 verzeichneten Habitattypen wurden in jedem Zeitfenster einmal abgesucht (ca. 15 min/0,5 ha). Besondere Flugbeobachtungen wurden ebenfalls protokolliert. Die Einteilung der Flugaktivität in eine der drei Klassen gering, durchschnittlich und hoch erfolgte jeweils unmittelbar nach Abschluss einer Erfassung. Die Klassen beziehen sich immer auf die gesamte Aktivität im bearbeiteten Untersuchungsgebiet und sind somit zwischen verschiedenen Untersuchungsgebieten nicht vergleichbar.
2.4 Habitatnutzung
Die quantitative Erfassung der Flugaktivität erfolgte im Rahmen einer Semesterarbeit an der Zürcher Hochschule für Angewandte Wissenschaften Wädenswil (ZHAW) in den drei Untersuchungsgebieten HE, GL und ME vom 07.05. bis 26.05.2011 (Tschumi 2011). In jedem Untersuchungsgebiet fanden an jeweils drei Begehungsterminen Untersuchungen in je drei Habitattypen statt: in einer landwirtschaftlich intensiv und einer extensiv genutzten Parzelle sowie im Eiablagehabitat. In jedem Habitattyp wurden je drei Transekte definiert, für die während je 15min alle Überflüge von L. coccajus gezählt wurden. Für weitere methodische Details siehe Tschumi (2011).
2.5 Mobilität
In den Untersuchungsgebieten GL und ME wurde ein Markierungsexperiment an weiblichen Individuen durchgeführt. Die Untersuchung beschränkte sich auf ein Geschlecht, um eine Verzerrung der Resultate durch mögliche geschlechterspezifische Unterschiede zu vermeiden. Dazu wurden 2 (ME) bzw. 3 (GL) je 100 m2 große Bereiche (10 x 10 m, vgl. Abb. 4 und 5) ausgeschieden, innerhalb derer je zehn Individuen gefangen, auf dieselbe Art (entweder auf dem linken, dem rechten oder beiden Hinterflügeln) an den Hinterflügeln markiert und wieder freigelassen wurden. Die Markierung erfolgte so, dass im Flug festgestellt werden konnte, in welchem der Bereiche das Individuum gefangen wurde.
Die Markierung der Individuen erfolgte am 17.05.2011 (GL) und am 18.05.2011 (ME) mit gelbem, verdünntem Nagellack (1:5 verdünnt mit Aceton, vgl. Keller et al. 2010). Die nachfolgende Suche nach markierten Individuen fand danach jeweils am ersten Tag mit optimalen Flugbedingungen statt. Im gesamten Untersuchungsgebiet wurden so viele Flugbewegungen von markierten Individuen wie möglich auf einem Plan (1:1000) eingezeichnet und die maximale Anzahl gleichzeitig fliegender Individuen mit identischer Markierung in den einzelnen Flächen festgehalten.
2.6 Auswertungen
Die Digitalisierung der Beobachtungen des Fluggebiets und des Markierungsexperiments erfolgte mittels ArcMap 10.0, ebenso die Berechnung der minimal genutzten Fläche für jedes Zeitfenster anhand der minimal convex Polygon-Methode (Dalk & Sime 1938). Darauf basierend wurde für jedes Untersuchungsgebiet eine Übersicht erstellt, welche Lebensräume in welchem Zeitabschnitt beflogen wurden und wie groß die Intensität war.
Für jede Begehung und jeden Habitattyp wurde der Anteil an den gesamthaft an diesem Tag beobachteten Flugaktivität berechnet. Für jede Flugbewegung des Markierungsexperiments wurde eruiert, welche der in Abb. 4 und 5 aufgeführten Lebensräume beflogen wurden. Zudem wurde die größte Entfernung zwischen dem Zentrum eines Markierungsgebietes und den in diesem Bereich markierten Individuen gemessen.
3 Resultate
3.1 Fluggebiet
In Tab. 1 sind die als Fluggebiete genutzten Lebensräume der vier Untersuchungsgebiete aufgeführt. Die Flugaktivität der verschiedenen als Fluggebiet genutzten Lebensräume veränderte sich im Tagesablauf nur minimal.
In allen Untersuchungsgebieten wurde festgestellt, dass die Fluggebiete relativ gleichmäßig beflogen wurden (vgl. auch Anhang 1 bis 4 unter http://www.nul-online.de Service Download). In GL flogen viele Individuen über der westlichen Ackerfläche, einem Rapsfeld (Abb. 2: GL 3.1), ebenso entlang der Autobahnböschung (Abb. 2: GL 1) und über den beiden nördlich bzw. nordöstlich gelegenen Magerwiesen (Abb. 2: GL 6.1 & 6.2 und Anhang 1; Download). In diesem Untersuchungsgebiet überflogen zudem einige Individuen eine dichte, ca. 3 bis 4m hohe Strauchhecke (Abb. 2: GL 2).
In HE konnten einige fliegende Individuen im lichten Föhrenwald beobachtet werden, jedoch immer in Bereichen mit sehr geringem Kronenschlussgrad bzw. auf offenen Lichtungen (vgl. Anhang 2; Download). Diese Individuen flogen regelmäßig bis in die Wipfel der Föhren (ca. 68m) und darüber hinaus. Allerdings konnte nicht verfolgt werden, ob diese Individuen zurückkehrten oder das Waldgebiet verließen. Die Flugaktivität war über der Fettwiese nordöstlich des lichten Waldes (Abb. 2: HE 4.2) am größten. Bei einem informellen Besuch am 09.06.2010 in HE wurde festgestellt, dass diese Fettwiese gemäht war. Zu diesem Zeitpunkt war die Flugintensität über der Magerwiese südlich des lichten Waldes (Abb. 2: HE 2) am größten, über der gemähten Wiese konnte kein fliegendes Individuum beobachtet werden.
Dagegen konzentrierte sich die Flugaktivität in ME hauptsächlich auf zwei Trockenrasen, die sich an einem Hang in Buchten des Waldrandes befanden (vgl. Anhang 3; Download). Während die östliche Fläche einige Gebüsche aufwies (Abb. 2: ME 1.1) war die westliche praktisch unverbuscht (Abb. 2: ME 1.2). In der Fläche ME 1.1 wurden verschiedene Individuen beim Überflug von bis zu 5m hohen Gebüschgruppen beobachtet. Zudem bewegten sich einige Individuen in großer Höhe Richtung Waldrand, es konnte jedoch nicht eruiert werden, ob diese Individuen tatsächlich über den Wald flogen oder zuvor abdrehten.
In RH flogen die meisten Individuen im Bereich von zwei durch eine Baumhecke geschützten Buchten (Abb. 2: RH 1.1) und eine angrenzende Fettwiese (Abb. 2: RH 3.1). Überflüge über die Baumhecken konnten hier jedoch nicht beobachtet werden, auch die an die Fettwiese angrenzenden Getreideäcker (u.a. Gerste / Mais) (Abb. 2: RH 4.1) wurden nicht als Fluggebiet genutzt (vgl. Anhang 4; Download).
In GL war das mittels minimum convex polygon ermittelte durchschnittliche Fluggebiet flächenmäßig am größten (3,8 ha), die Werte der anderen drei Untersuchungsgebiete waren etwa vergleichbar (HE: 1,2 ha, ME: 0,8 ha, RH: 1,0 ha). Wie wir beobachteten, kam in allen Untersuchungsgebieten ein großer Teil der Flugaktivität durch jagende Individuen zustande, was an der sehr schnellen Flugweise erkennbar war, verbunden mit kurzen, ruckartigen Flugbewegung zum Fangen der Beute. Zudem wurden immer wieder Paarungsversuche und vereinzelt auch erfolgreiche Paarungen beobachtet.
3.2 Habitatnutzung
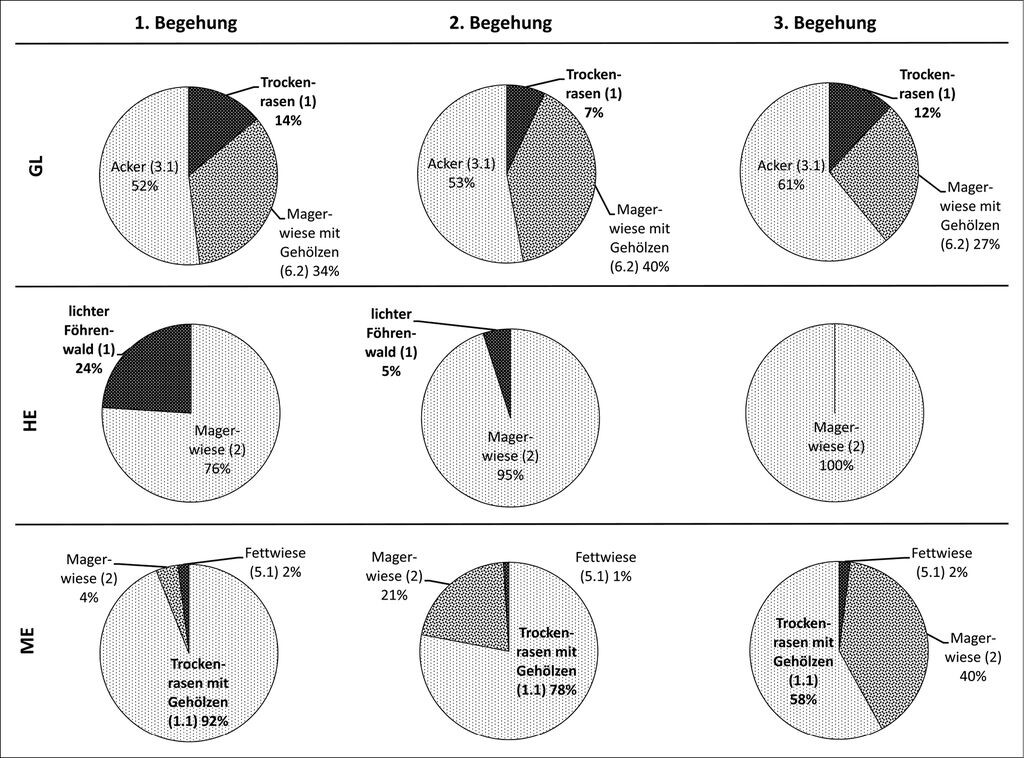
InAbb. 3 ist dargestellt, wie sich die Flugaktivität 2011 auf die verschiedenen Lebensräume aufgeteilt und im Laufe der drei Begehungen zwischen dem 07. und 26. Mai verändert hat. Im Durchschnitt wurden über einem Transekt 4,8 Überflüge pro Minute beobachtet, in GL im Mittel 8,2 und in HE 2,2 sowie in ME 1,3 Überflüge/min. Witterungsbedingt konnte die Erfassung der Fettwiese in HE bei der ersten Erfassung (08.05.2011) nicht durchgeführt werden. Diese wurde 21.05.2011 zusammen mit der zweiten Begehung nachgeholt. Allerdings wurden über der zu diesem Zeitpunkt bereits gemähten Wiese keine Überflüge mehr festgestellt. Während in GL und HE im Eiablagehabitat maximal ein Viertel der erfassten Flugaktivität stattfand, wurde in ME mehr als die Hälfte der Flugaktivität im Eiablagehabitat beobachtet. Hierzu gilt es anzumerken, dass in ME zwischen der ersten und der dritten Begehung eine deutliche Verlagerung der Aktivität vom Eiablagehabitat (Trockenrasen mit Gehölzen) zur Magerwiese erfolgte.
3.3 Mobilität
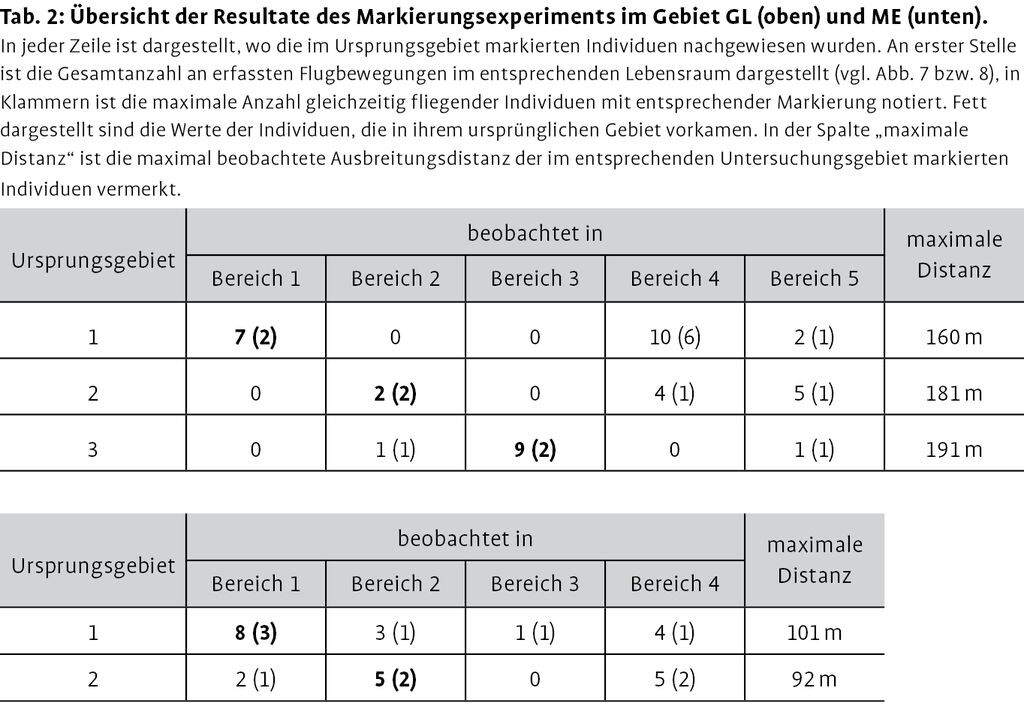
Insgesamt konnten 59 Flugstrecken von markierten Individuen festgehalten werden. In GL wurden von den 30 markierten Individuen an einem Tag 40 Flugstrecken protokolliert, in ME von 20 markierten Individuen 19 Flugstrecken. Abb. 4 und 5illustrieren, in welchen Bereichen markierte Individuen erfasst wurden, Tab. 2 gibt einen quantitativen Überblick der beobachteten markierten Individuen. Die maximalen nachgewiesen Distanzen waren in GL deutlich größer als in HE (vgl. Tab. 2). In ME konnte ein markiertes Individuum beobachtet werden, welches von der Fläche 1 über die Strauchhecke in die Fläche 3 flog. Ein weiteres, nicht markiertes Individuum wurde dabei beobachtet, wie es den Gehölzbereich zwischen den Flächen 1 und 2, welcher ca. 30m breit und 6 bis 8m hoch ist, überquert hat.
4 Diskussion
4.1 Fluggebiet und Ausbreitungsverhalten
Unsere Studien zu den von L. coccajus genutzten Habitattypen zeigen, dass ein breites Spektrum verschiedener Lebensräume als Fluggebiet genutzt wird. Wir konnten fliegende Individuen in einem lichten Föhrenwald, über Trockenwiesen mit verschieden großem Anteil an Gebüschen und Bäumen, über einem Maisacker, einem Rapsfeld, einer Fettwiese sowie an einer künstlichen Autobahnböschung, deren Vegetation als Trockenrasen charakterisiert werden kann, nachweisen. Dieses widerspricht zumindest teilweise den Beobachtungen von Fetz (2002) und Wolf (2005), welche im Taubertal (Bayern) die Art nur über Esparsetten-Halbtrockenrasen und Salbei-Glatthaferwiesen beobachteten und daher als stenotop einschätzen.
Die Beobachtung fliegender Individuen über intensiv bewirtschafteten Parzellen (Fettwiese, Mais- bzw. Rapsacker) ist insbesondere für die Planung von Biotopverbundsystemen von Bedeutung. Es verdeutlicht, dass intensiv bewirtschaftete Flächen für L. coccajus nicht zwingend eine stark isolierende Wirkung haben vorausgesetzt, dass weitere geeignete Habitate in einer für die Art überwindbaren Entfernung vorhanden sind.
Vergleicht man die Fläche des gesamten Fluggebiets der verschiedenen Untersuchungsgebiete miteinander, ist augenfällig, dass die Population GL ein Gebiet nutzte, welches mit rund 3.8 ha mehr als doppelt so groß war wie diejenigen der restlichen Populationen (Tab. 1). Zudem war die durchschnittliche Flugaktivität in diesem Untersuchungsgebiet etwa um den Faktor 4 größer als in den übrigen Untersuchungsgebieten. Mit hoher Wahrscheinlichkeit war diese Population die individuenstärkste von allen, wobei unklar ist, worauf das zurückzuführen ist. Mögliche Einflussfaktoren könnten die Größe des Fluggebiets, die Verfügbarkeit von Nahrung oder die Qualität der Eiablagehabitate sein.
Auf eine Interpretation bezüglich der Flächenansprüche von L. coccajus wird verzichtet, da die Genauigkeit der verwendeten minimum convex polygon-Methode kontrovers diskutiert wird (vgl. Nilsen et al. 2008) und zudem nebst der Flächengröße auch die Qualität des Habitats dabei eine zentrale Rolle spielt (Thomas et al. 2001).
Bezüglich der Ausbreitungsmöglichkeiten der weiblichen Individuen von L. coccajus muss die von uns beobachtete maximale Ausbreitungsdistanz kritisch hinterfragt werden. Die maximal festgestellten Ausbreitungsdistanzen waren mit 191m (Glattfelden) bzw. 101m (Merishausen) eher gering, vergleich man sie mit anderen flugfähigen Insekten (z.B. Roter Scheckenfalter, Melitaea didyma (Nymphalidae): 9000m (Brunzel & Reich 1996); Helm-Azurjungfer Coenagrion mercuriale (Coenagrionidae) 1790m (Hassall & Thompson 2012)). Von Kissling (2011) wurde für das Esparsetten-Widderchen (Zygaena carniolica (Zygaenidae) jedoch ein Aktionsradius von 200 bis 300m angegeben, in dem sich alle Individuen einer Population bewegen.
Da unsere Methode nur Beobachtungen im Untersuchungsgebiet zuließ, sind die von uns ermittelten Distanzen nicht als maximale Ausbreitungsdistanzen von herumstreuenden Individuen zu verstehen, sondern vielmehr als maximaler Aktionsradius eines Individuums in seinem angestammten Habitat. Settele & Reinhardt (1999) weisen zudem darauf hin, dass die Mobilität von Schmetterlingen aufgrund von Freilandexperimenten fast immer unterschätzt wird und Ergebnisse von Markierungsexperimenten lediglich als minimal überwindbare Entfernungen anzusehen sind.
4.2 Habitatnutzung
Basierend auf unseren Untersuchungen der Habitatnutzung von L. coccajus konnten wir aufzeigen, dass die Flugaktivität nicht per se vom Vegetationstyp einer Fläche abhängt und sich im Verlauf einer Flugperiode deutlich verändern kann.
Dass kein direkter Zusammenhang zwischen dem Vegetationstyp und der Flugaktivität besteht, zeigt die Beobachtung, dass in Glattfelden ein Acker in beiden Untersuchungsjahren sehr intensiv beflogen wurde, während auf den Ackerflächen in Merishausen und Rheinheim keine Überflüge stattfanden. In Glattfelden wurde 2010 Raps und 2011 Mais angebaut, in Merishausen und Rheinheim in beiden Untersuchungsjahren unter anderem Mais und Gerste. Weiter konnten wir in Hessenberg und Rheinheim viele fliegende Individuen über einer Fettwiese beobachten, während die Fettwiesen in Merishausen gemieden wurden. Wir gehen davon aus, dass diese selektive Habitatnutzung mit den lokalen Windverhältnissen zusammenhängt. Während der Acker in Glattfelden durch den umgebenden Wald relativ windgeschützt war, präsentierten sich die Äcker in Merishausen und Rheinheim eher windexponiert. Aufgrund dieser Beobachtungen erscheint es naheliegend, dass windgeschützte Bereiche von L. coccajus bevorzugt werden. Auch Fetz (2002) geht davon aus, dass windberuhigte Habitate für die Art vorteilhaft sind. Derartige positive Effekte windgeschützter Bereiche wurden auch für verschiedene Tagfalterarten nachgewiesen (z.B. Dover et al. 1997, Rosin et al. 2011).
Eine Anpassung der Flugaktivität von L. coccajus an veränderte Standortbedingungen konnte unter anderem in Hessenberg beobachtet werden. Im Jahr 2010, zu einem Zeitpunkt, als sowohl die Fettwiese nordöstlich des lichten Waldes (Abb.2 HE 4.2) als auch die Magerwiese südwestlich des lichten Waldes (Abb. 2 HE 2) noch nicht gemäht waren, wurden fliegende Individuen ausschließlich über der Fettwiese festgestellt. Nachdem die Fettwiese 4.2 jedoch gemäht war, wurden fliegende Individuen nur noch über der Magerwiese nachgewiesen. In diesem Fall könnte die verminderte Flugaktivität mit einem Mangel an Beutetieren zusammenhängen oder mit der fehlenden Möglichkeiten für die Tiere, sich bei Bedarf in die Vegetation zu setzen, wie sie das z.B. bei aufkommendem Wind tun. Dieselben Gründe dürften ebenfalls erklären, wieso sich die Flugaktivität in Glattfelden 2010 fast komplett auf den Maisacker beschränkte, während die vegetationslose Ackerparzelle östlich davon fast keine Flugaktivität aufwies. Einen gravierenden Einfluss des Windes auf die Flugaktivität über der östlichen Ackerparzelle kann aufgrund von Beobachtungen während des Markierungsexperimentes 2011 größtenteils ausgeschlossen werden. In dieser Zeit wurden auch über dem östlichen Acker, auf dem zu dieser Zeit eine nicht näher bestimmte, rund 20cm hohe Feldfrucht wuchs, einige fliegende Individuen beobachtet.
Da ein großer Teil der beobachteten Flugaktivität auf jagende Individuen zurückzuführen war, gehen wir davon aus, dass die Verfügbarkeit von Beutetieren ein weiterer Faktor bei der Auswahl der Aktivitätsgebiete ist. Aufgrund des sehr geringen Kenntnisstandes des Nahrungsspektrums von L. coccajus ist es jedoch schwierig, diesbezüglich Aussagen zu machen. Es ist aber anzunehmen, dass die fehlende Flugaktivität über der gemähten Fettwiese zumindest teilweise damit zusammenhängt, da beispielsweise die Anzahl blütenbesuchender Insekten mit Sicherheit drastisch vermindert war.
Sowohl unsere Beobachtungen als auch diejenigen von Wolf (2005) weisen darauf hin, dass die Kopulation räumlich in denselben Bereichen stattfindet wie die Jagd. Die von uns erfassten Fluggebiete können somit als Jagdgebiete, in welchen auch die Fortpflanzung stattfindet, bezeichnet werden. Charakteristisch für diese Lebensräume sind ihre windgeschützte Lage, das Vorkommen der bislang nicht näher bekannten Beute von L. coccajus und eine Vegetation, die bei widrigen Flugbedingungen Rückzugsmöglichkeiten bietet.
4.3 Mobilität
Eine weitere wichtige Erkenntnis liegt darin, dass Strauchhecken und schmale Waldbereiche (bis 30m Breite) überwunden werden können, wobei wir in Rheinheim eine Einschränkung der Mobilität durch eine sehr hohe Baumhecke (>10m) beobachteten. Diese Beobachtungen widersprechen teilweise den Erkenntnissen von Fetz (2002) und Wolf (2005), die für das von ihnen untersuchte Gebiet (Taubertal bei Rothenburg o.d.T) feststellten, dass die Art nur über offenen Grünlandflächen fliegt und Hecken und bewachsene Steinriegel aus intrinsischen Gründen nicht überquert. Sehr wahrscheinlich hat die Höhe eines Gehölzelementes einen zentralen Einfluss darauf, ob es als Ausbreitungshindernis wirkt. Da es sich bei L. coccajus um eine sehr wärmebedürftige Art handelt, ist denkbar, dass in einem kühleren Klima auch die Mobilität reduziert ist. Dieses könnte erklären, weshalb die Populationen, die von Fetz (2002) und Wolf (2005) untersucht wurden und rund 200km weiter nördlich liegen als die von uns untersuchten, größeren Einschränkungen bezüglich ihrer Mobilität unterlagen.
4.4 Weiterer Forschungsbedarf
Zur Ausbreitungsfähigkeit von L. coccajus ist der Wissensstand nach wie vor lückenhaft. Bezüglich einer Limitierung der Ausbreitungsmöglichkeit durch Wind erlauben unsere Daten keine abschließende Beurteilung und auch die maximale Ausbreitungsdistanz ist nach wie vor unklar. Da der beobachtete Zusammenhang zwischen den Windverhältnissen und der Flugaktivität auch dadurch zustande kommen könnte, dass bei aufkommendem Wind die Beutetiere ihre Flugaktivität einstellen, kann nicht direkt daraus gefolgert werden, dass die Ausbreitung der Art durch Wind limitiert wird. Vertiefte Studien zum Beutespektrum des Libellen-Schmetterlingshafts und zur großräumigen Ausbreitungsfähigkeit könnten weitere wertvolle Informationen zum Schutz dieser attraktiven Art liefern. Zudem muss auch beachtet werden, dass wir in unserer Untersuchung nur weibliche Tiere markiert haben, vergleichende Untersuchungen mit männlichen Individuen wären auf jeden Fall angezeigt.
Dank
Wir möchten uns ganz herzlich bei Frau I. Flöss (Kanton Aargau) für die fachliche Unterstützung und bei U. Kuhn (Kanton Zürich) und H. Billing (Kanton Schaffhausen) für die Bewilligung des Markierungsexperiments bedanken.
Literatur
Aspöck, H., Aspöck, U., Hölzel, H. (1980): Die Neuropteren Europas, Bd.1. Goecke & Evers, Krefeld, 494S.
Baier, H., Erdmann, F., Holz, R., Walterstraat, A. (2006): Freiraum und Naturschutz. Die Wirkungen von Störungen und Zerschneidungen in der Landschaft. Springer, Berlin, 684S.
Brunzel, S., Reich, M. (1996): Zur Metapopulationsstruktur des Roten Scheckenfalters (Melitaea didyma, Esper 1779) auf der Schwäbischen Alb. Zeitschr. Ökol. Naturschutz 5, 243-253.
Dalk, P.D., Sime, P.R. (1938): Home and seasonal ranges of the eastern cotton tail in Connecticut. Transcripts of the Northern American Wildlife Conference 3, 659-669.
Dover, J.W., Settele, J. (2009): The influence of landscape structure on butterfly distribution and movement: a review. Journal of Insect Conserv. 13, 3-27.
, Sparks, T.H., Geratorex-Davies, J.N. (1997): The importance of shelter for butterflies in open landscapes. Journal of Insect Conserv. 1, 89-97.
Duelli, P. (1994): Rote Liste der gefährdeten Netzflügler der Schweiz. In: Duelli, P., Hrsg., Rote Listen der gefährdeten Tierarten der Schweiz, Bundesamt für Umwelt, Bern, 48-51.
Fetz, T. (2002): Zoologische Aspekte des Magerrasen-Managements am Beispiel des Libellen-Schmetterlingshafts (Libelloides coccajus). Bayer. Landesamt Umweltschutz 167, 73-79.
Fischer, R. (2009): Der Schmetterlingshaft Libelloides coccajus [DENIS & SCHIFFERMÜLLER, 1775] im Aargau-Schaffhausen-Zürich. Unveröff. Bacholor-Thesis, ZHAW, Zürcher Hochschule für angewandte Wissenschaften Wädenswil, 76S.
Gonseth, Y. (1991): LAscalaphe, Libelloides coccajus (Denis & Schiff., 1775), (Neuropteroidae, Planipennia) dans le Canton de Neuchâtel. Bulletin Romand dEntomologie 9, 49-59.
Günter, R. (1999): Die Amphibien und Reptilien Deutschlands. Gustav Fischer, Jena, 825S.
Hasall, C., Tompson, D.J. (2012): Study design and mark recapture estimates of dispersal: a case study with the endangered damselfly Coenagrion mercuriale. Journal of Insect Conserv. 16, 111-120.
Jedicke, E. (1994): Biotopverbund. Ulmer, Stuttgart, 2. Aufl., 287 S.
Keller, D., Brodbeck, S., Flöss, I., Vonwil, G., Holderegger, R. (2010): Ecological and genetic measurements of dispersal in a threatened dragonfly. Biol. Conserv. 143, 2658-2663.
Kissling, S. (2011): Bedeutung des Biotopverbundes für die Zielart Esparsetten-Widderchen. Naturschutz und Landschaftsplanung 43 (2), 46-54.
Lachat, T., Pauli, D., Gonseth, Y., Klaus, G., Scheidegger, C., Vittoz, P. Walter, T. (2010): Wandel der Biodiversität in der Schweiz seit 1900. Ist die Talsohle erreicht? Bristol-Stiftung, Zürich, Haupt, Bern, 435S.
Müller, M., Schlegel, J., Krüsi, B.O. (2012): Der Libellen-Schmetterlingshaft (Libelloides coccajus [Denis & Schiffermüller 1775] Neuroptera: Ascalaphidae) im Kanton Aargau: Aktuelle Vorkommen und Empfehlungen zum Artenschutz. Mitt. Schweiz. Ent. Ges. 85, 177-199.
, Schlegel, J., Krüsi, B.O. (einger.): Selection of oviposition sites by Libelloides coccajus (Neuroptera: Ascalaphidae) north of the Alps: implications for nature conservation. Insect Conservation and Diversity.
Nilsen, E.B., Pederson, S., Linell, J.D.C. (2008): Can minimum convex polygon home range be used to draw biologically meaningful conclusions? Ecol. Research 23, 635-639.
Pröse, H., Gruppe, A. (2003): Rote Liste gefährdeter Netzflügler (Neuropteroidae) Bayerns. Schr.-R. Bayer. Landesamt Umweltschutz 166, 95-98.
Rosin, Z.M., Myczko, L., Skorka, P., Lenda, M., Moron, D., Sparkts, T. H. Tryjanowski, P. (2011): Butterfly responses to environmental factors in fragmented calcareous grasslands. Journal of Insect Conserv., published with open access, DOI 10.1007/s10841-011-9416-5.
Settele, J., Margules, C., Poschlod, P., Henle, K. (1996): Species Survival in Fragmented Landscapes. Kluwer, Dordrecht, 381pp.
, Reinhardt, R. (1999): Ökologie der Tagfalter Deutschlands: Grundlagen und Schutzaspekte. In: Settele, J., Feldmann, R. Reinhardt, R., Hrsg., Die Tagfalter Deutschlands, Ulmer, Stuttgart, 452S.
Thomas, J.A., Bouron, N.A.D., Clarke, R.T., Stewart, K.E., Simcox, D.J., Pearman, G.S., Curtis, R., Goodger, B. (2001): The Quality and isolation of habitat patches both determine where butterflies persist in fragmented landscapes. Proceedings of the Royal Society B, 268, 1791-1796.
Tröger, E.J. (1998): Gesamtverzeichnis der Netzflügler (Neuropteroidea) und Rote Liste gefährdeter Arten in Baden-Württemberg. Stand Juli 1997. In: Köppel, C., Rennwald, E., Gigion, A., Hrsg., Rote Listen auf CD-Rom Deutschland, Österreich, Lichtenstein, Schweiz, Süd-Tirol, Verlag für interaktive Medien, Gaggenau, 78-80.
Tschumi, E. (2011): Raumnutzungsanalyse des Libellen-Schmetterlingshafts (Libelloides coccajus) in der Nordschweiz. Unveröff. Bachelor-Thesis, ZHAW Zürcher Hochschule für angewandte Wissenschaften, Wädenswil, 53S.
Wolf, W. (2005): Der Libellen-Schmetterlingshaft Libelloides coccajus im bayerischen Taubertal Lebensraum, Ansprüche, Entwicklungsdaten. Laufener Seminarbeitr. 1/04, 61-66.
Anschrift der Verfasser: Markus Müller, SKK Landschaftsarchitekten AG, Lindenplatz 5, CH-5430 Wettingen 1, Schweiz, E-Mail markus.mueller@skk.ch; Prof. Dr. Bertil O. Krüsi und Jürg Schlegel, Institut für Umwelt und Natürliche Ressourcen, ZHAW Zürcher Hochschule für Angewandte Wissenschaften, CH-8820 Wädenswil, Schweiz.
-
Abb. 1: Ein weiblicher Libellen-Schmetterlingshaft, der in Glattfelden auf dem rechten Hinterflügel markiert wurde. Die Markierung wurde so großflächig aufgetragen, damit sie auch im Flug aus mehreren Metern Entfernung noch von Auge erkennbar war. Foto: Markus Müller -
Abb. 2: Ortho-Luftphotos der vier Untersuchungsgebiete. Die Untersuchungsgebiete sind gestrichelt eingezeichnet. Zusätzlich sind die wichtigsten Habitattypen bezeichnet (Erklärungen siehe Tab. 1). Geodaten © swisstopo (DV084370). Reproduziert mit Bewilligung von swisstopo (BA12039). -
Abb. 3: Veränderung des durchschnittlichen prozentualen Anteils der Flugaktivität innerhalb der verschiedenen Habitate der drei Untersuchungsgebiete (Daten: Tschumi 2011). Die Ziffern in Klammern entsprechen den Lebensräumen aus Abb. 2. Die Habitattypen, in welchen die Eiablage erfolgte, sind jeweils fett dargestellt. Die Zahl der erfassten Überflüge pro Erfassung lag zwischen 98 und 1748. Die Erfassungen erfolgten zwischen dem 07.05. und dem 26.05.2011. -
Abb. 4: Darstellung der Mobilität der markierten weiblichen Schmetterlingshafte in GL. Die schwarzen Quadrate entsprechen den Bereichen, in welchen markiert wurde. In den gestrichelt eingezeichneten Flächen wurde nach drei Tagen nach markierten Individuen gesucht. Die Pfeile zeigen an, in welchen Gebieten markierte Individuen beobachtet wurden. Dünne Pfeile entsprechen 15, dicke Pfeile >5 gleichzeitig fliegenden markierten Individuen in den neu genutzten Gebieten. Reproduziert mit Bewilligung von swisstopo (BA12039). -
Abb. 5: Darstellung der Mobilität der markierten weiblichen Schmetterlingshafte in ME. Weitere Erläuterung wie in Legende zu Abb. 4. -
Raumnutzung des Libellen-Schmetterlingshafts nördlich der Alpen -
Raumnutzung des Libellen-Schmetterlingshafts nördlich der Alpen -
Raumnutzung des Libellen-Schmetterlingshafts nördlich der Alpen





Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.