Auswirkungen der Windenergie auf Tierarten
Abstracts
In den letzten Jahren ist eine zunehmende Begleitung des Windenergieausbaus mit qualitätssichernden Publikationen zu der Frage der Auswirkungen auf die Tierwelt feststellbar. Das Ziel ist nicht nur, Kenntnisse zur bestmöglichen Vermeidung von negativen Wirkungsbeziehungen zwischen bestimmten Tierartengruppen und Windenergieanlagen (WEA) für die Anlagenplanung zur Verfügung zu stellen, wie beispielsweise zu der Frage der Kollisionen von Wildvögeln oder Störungen der Meeresumwelt. Auch das europäische und nationale Artenschutzrecht verlangt explizit, relevante Natur- und Artenschutzeffekte zu bewältigen.
Mit dem Ziel, synoptisch den aktuellen Stand der Forschung aufzuzeigen, wird in diesem Beitrag auf Basis einer an der TU Berlin durchgeführten Literaturanalyse dargelegt, welche Kenntnisse über die Auswirkungen der Windenergie auf die Tierwelt zwischenzeitlich als angenommen gelten können und wo weiterhin erhebliche Kenntnislücken bestehen, beispielsweise aufgrund von sich widersprechender Studienergebnissen.
Einige Effekte wurden bereits häufiger untersucht, wie etwa der Einfluss von Standortfaktoren auf die Raumnutzung von Greifvögeln, während zu anderen weiterhin stark im Diskurs stehenden Fragen, wie z.B. zu Effekten auf der Populationsebene, weniger belegte Kenntnisse bestehen. So ergeben sich eine Reihe weiterführender Fragestellungen; Monitoring-Daten sollten z.B. auf geeignete Art zur Verfügung gestellt werden, um beim weiteren Ausbau der Windenergie Auswirkungen auf die Tierwelt noch besser informiert begegnen zu können.
Effects of wind power on animal species Synoptical overview
During the last years the development of wind energy has been accompanied by publications aiming to ensure quality control on the question of effects on wildlife species. The aim is not only to provide knowledge for the best possible avoidance of negative relations between certain animal species and wind energy for facility planning, such as the question of bird collisions or biotic disturbances in the marine environment. Additionally, both European and national species protection laws explicitly require to deal with all relevant effects on nature and endangered species. The paper aims to synoptically illustrate the state of research. On the one hand it shows what kind of knowledge about the impacts of the wind energy on wildlife species can be considered as accepted for practical planning. On the other hand it aims to identify remaining knowledge gaps, for example caused by contradicting study results. The results presented here are based on an international synopsis carried out at the Institute of Technology Berlin (TU Berlin).
Some effects have already been investigated several times, such as the influence of locational factors on the spatial behaviour of birds of prey, whilst only little secure knowledge has been gained on other important questions, such as the habituation of migratory birds to wind parks or impacts on population level.
These findings have led to a number of subsequent questions, e.g. the availability of monitoring data in a suitable way. This would help to increase information of potential impacts on wildlife during the further development of wind energy.
- Veröffentlicht am

Not a rare picture in Northern Germany migrating cranes (Grus grus) in front of wind power plantsARSU GmbH
1 Einleitung
Mit dem weltweiten Ausbau der Windenergie an Land und auf dem Meer verbleiben Bedenken und Herausforderungen hinsichtlich der unbeabsichtigten Nebenwirkungen auf die Tierwelt (Abb.1). Gleichzeitig kommt stetig neuer Erkenntnisbedarf hinzu, etwa bezüglich der beträchtlich gewachsenen Anlagengrößen und den kumulativen Effekten des Zubaus. Um die Zielsetzungen des international vereinbarten Ausstiegs aus der Nutzung fossiler Energieträger auch bei der Stromerzeugung zu gewährleisten, muss dies dennoch zu den geringstmöglichen Natur- und Artenschutz-Kosten pro kWh erfolgen (May et al., angen., vgl. van Kuik et al. 2016), d.h. dass Belange von geschützten Arten nicht beeinträchtigt werden. Dies verlangt einen umfassenden Einblick in die Effekte von Windenergieanlagen (WEA) und Windparks auf die Tierwelt, wie Störungen, Habitatverlust, Kollisionsopfer (z.B. Bailey et al. 2010, Kunz et al. 2007, Smallwood & Thelander 2008), sowie eine Diskussion möglicher Auswirkungen auf die Populationsebene betroffener Arten (vgl. Grünkorn et al. 2015).
Seit den frühen 2000-er Jahren hat sich dazu die Anzahl wissenschaftlich qualitätsgesicherter Publikationen international mehr als verzehnfacht, gemessen etwa anhand der Eintragungen in der Wind-Wildlife Impacts Literature Database ( https://wild.nrel.gov/ ) des National Renewable Energy Laboratory in den USA (May et al., angen.). Wesentlich dazu beigetragen hat in Europa eine Konferenzserie, die 2011 in Trondheim/Norwegen das erste Mal stattfand und nach 2013 in Stockholm/Schweden schließlich 2015 an der TU Berlin ausgetragen wurde ( http://www.cww2015.tuberlin.de ).
In jüngerer Zeit kamen tiergruppenspezifische synoptische Arbeiten hinzu ein Ausdruck des gewachsenen Bedürfnisses, aus der Vielzahl der Arbeiten einen Überblick zu erhalten, was wir bislang zur Thematik zu wissen scheinen bzw. auch noch nicht schlussfolgern können (z.B. Cryan & Barclay 2009 für Fledermäuse sowie Marques et al. 2014 für Vögel; ähnlich zu möglichen Vermeidungsmaßnahmen May et al. 2015 v.a. für Vögel, Peste et al. 2015 für Fledermäuse). Dies ist letztlich möglich geworden aufgrund einer wohl einmaligen Forschungsdichte zu diesem Thema. Auch in Deutschland ist erheblich in die ökologische Begleitforschung beim Ausbau der Windenergie investiert worden. Gleichermaßen stoßen solche Zwischenfazits jedoch aufgrund des anhaltend dynamischen Zuwachses an betreffenden Studien an ihre Grenzen, so dass Wissens-Plattformen wie Thethys ( http://tethys.pnnl.gov/ ) zunehmend an Bedeutung gewinnen, wie auch die internationale Kooperation WREN auf dieser Plattform (Copping et al. 2015.).
Dabei stellt z.B. das europäische und nationale Artenschutzrecht mit der FFH- und Vogelschutzrichtlinie auch in Deutschland strenge Anforderungen an die Zulassung von WEA, insbesondere sofern relevante Arten durch ein signifikantes Tötungsrisiko durch den Bau und Betrieb der Windenergienutzung betroffen sein können (Reichenbach 2015). Zur Konzeption möglicher Vermeidungsmaßnahmen in allen in Frage kommenden Phasen der Planung und des Betriebs eines Windparks (Gartman et al. 2016) ist es erforderlich, hinreichend die möglichen Wirkungsweisen von WEA auf die betroffenen Tiergruppen und Arten zu erkennen. Schon deshalb schien es an der Zeit, den internationalen Wissensstand zu bündeln und eine Zwischenbilanz zum Stand des Wissens zu ziehen. Während bislang vorliegende Synopsen (z.B. Arnett et al. 2011a, Arnett & Baerwald 2013, Bergström et al. 2013a, Helldin et al. 2012, Lovich & Ennen 2013, Rydell et al. 2012) sich auf bestimmte Artengruppen konzentrieren oder nationale Schwerpunkte aufweisen, haben Schuster et al. (2015; open access) den internationalen Forschungsstand zu den Auswirkungen auf Fledermäuse, (Greif-, Zug- und Rast sowie Zug-) Vögel sowie Meeressäugetiere übergreifend aufgezeigt. Andere Wildtierarten wurden z.B. infolge zu weniger Studien und Publikationen dabei nicht betrachtet, etwa Insekten oder Landsäugetiere (vgl. Boldt & Hummel 2013).
Entstanden ist ein Überblick darüber, welche Hypothesen durch Studien als soweit belegt gelten, aber auch welche Wirkungszusammenhänge weiterhin schwer verständlich bleiben, aufgrund fehlender, unvollständiger oder auch sich widersprechender Studienergebnisse. Zur besseren Verbreitung beleuchten wir hier in einer zusammengefassten Form wesentliche Aspekte zum betreffenden Kenntnisstand auf Basis dieser bislang international publizierten Arbeit (Schuster et al. 2015).
Im Folgenden wird zunächst der Weg der Hypothesenbildung, welche Auswirkungen der Windenergie als eher bekannt bzw. als eher unsicher beurteilt werden müssen, beleuchtet. In einem zweiten Schritt werden ausgewählte Ergebnisse für die genannten Artengruppen vorgestellt, diskutiert und verbleibende Unsicherheiten exemplarisch aufgezeigt. Der Beitrag schließt mit einem knappen Ausblick, in welchen Bereichen beispielsweise ein weitergehender Erkenntnisbedarf besteht.
2 Methode
Dieser Synopse liegt in einem ersten Schritt eine systematische, qualitative Literaturanalyse aus einem internationalen Blickwinkel auf Basis der Ergebnisse von Schuster et al. (2015) zugrunde. Insgesamt wurden über 220 Dokumente unterschiedlicher Publikationen in Hinblick auf die Effekte der Windenergienutzung auf Vögel, Fledermäuse und Meeressäugetiere berücksichtigt. Neben Studien mit Peer-Review (1) sowie Synopsen (Reviews) mit Peer-Review (2) wurden ebenfalls Studien/Berichte bzw. graue Literatur (4), d.h. nicht verlagsgebundene Veröffentlichungen, und Konferenzbeiträge (6) analysiert, um in einen großen Umfang den auch jüngeren Kenntnisstand der Auswirkungen der Windenergie abzubilden. Beispielsweise stellten internationale Konferenzen, wie die o.a. CWW 2011, Trondheim, die CWE 2013, Stockholm, sowie die Wind Wildlife Research Meetings (NWCC, USA) hierbei eine Basis für die entsprechend gekennzeichnete Berücksichtigung noch nicht wissenschaftlich publizierter Beiträge dar.
Allerdings erfolgten die Recherchen und Auswertungen bei Schuster et al. (2015) lediglich bis zum Jahr 2014 und decken nur wenige Beiträge aus 2015 ab; viel ist zwischenzeitlich weiter hinzugekommen. In einem zweiten Schritt wurden auf Grundlage dieses Literaturreviews sich abzeichnende Hypothesen hinsichtlich der Effekte von Windenergie auf die jeweiligen Tiergruppen identifiziert. Stets auf die Interaktionen von Wildtieren und WEA einwirkende Faktoren, wie standortbezogene Auswirkungen oder artspezifisches Verhalten im Einzelnen, wurden dabei nur exemplarisch berücksichtigt. Diese Hypothesen sind tabellarisch hinsichtlich ihrer Plausibilisierung und der herangezogenen Quellen in Schuster et al. (2015) dokumentiert. Hierbei wurden Hypothesen z.B. als mit Unsicherheiten belegt diskutiert, welche widersprechende Studienergebnisse aufweisen, von besonders wenigen Autoren erst untersucht wurden oder lediglich geringe Untersuchungszeiträume umfassen. Diese grundlegende Arbeitsstruktur und Darstellungsweise gemäß Schuster et al. (2015) wird in Tab. 1 verdeutlicht. Für den vorliegenden Beitrag erfolgte eine weiter aggregierte Darstellung einiger wesentlicher Wirkungshypothesen.
Einschränkend ist festzuhalten, dass die verschiedenen Studien und Publikationen auf i.d.R. unterschiedlichen Studiendesigns und Untersuchungszeiträumen beruhen und dass vielfältige regionalspezifische und auch artspezifische Charakteristika zu beachten sind. So ist beispielsweise das Verhalten des Golden Eagle (Steinadler, Aquila chrysaetos) bei der Nutzung des Aufwindes entlang von Kammlinien am Altamont Pass in Kalifornien und einem an diesen Standorten erhöhten Kollisionsrisiko mit WEA nicht unmittelbar mit dem Verhalten eines Greifvogels des Tieflands und Mittelgebirges in Deutschland vergleichbar. Dennoch handelt es sich meist um weltweit wirksame Gefahrenquellen von WEA, hier etwa für die Raumnutzung von Thermikfliegern (engl. soaring birds) bzw. Greifvögeln.
Desgleichen ist trotz des Versuchs, auf möglichst aktuelle Studienergebnisse zurückzugreifen, auf eine stetige weitere Anlagenentwicklung und lernende Planungsprozesse von WEA und Windparks zu achten. Bei der Auswahl der als eher bekannt bzw. als mit Unsicherheiten einzustufende Hypothesen gilt es zu bedenken, dass der Fall, in dem bei der Literaturanalyse keine Studien mit einem gegenteiligen Ergebnis gefunden wurden, dieses nicht bedeutet, dass eine Hypothese allein deshalb bestätigt oder verworfen werden kann. Auch die Anfertigung von synoptischen Beiträgen kann also mit zahlreichen Unsicherheiten behaftet sein. So ist die o.a. Kennzeichnung der jeweiligen Veröffentlichungsart und -reife von zentraler Bedeutung, z.B. ob eine Untersuchung ein Peer-Review durchlaufen hat. Andererseits ist zu beobachten, dass zahlreiche Konferenzbeiträge früher oder später in betreffende peer-reviewte Veröffentlichungen münden; schon daher haben wir auch Augenmerk auf solche Beiträge gelegt. Weiterhin sei abschließend auf den ebenfalls frei zugänglichen Abschlussbericht einer synoptischen Studie für das Schweizer Bundesamt für Energie (BFE) hingewiesen, welche den internationalen Kenntnisstand zum Einfluss der Windenergie auf Fledermäuse und Vögel mit einer Spezifizierung für die Schweiz erörtert (BFE 2015).
3 Zusammenfassung wesentlicher Wirkungshypothesen
Die Windenergienutzung kann auf vielfaltige Weise einen Einfluss auf die Tierwelt nehmen. Nach Drewitt & Langston (2006) sind die wichtigsten Effekte: Kollisionen, Verdrängung, Barriereeffekte, Habitatveränderungen und -verlust, wobei Kollisionen von Vögeln (Abb. 2, 3) und Fledermäusen die prägnanteste und am häufigsten beachtete und berichtete Form der Auswirkungen darstellt. Stets gelten standort-, jahreszeit- und artenspezifische Ausdifferenzierungen jenseits der hier synoptisch angesprochenen Auswirkungen (Pearce-Higgins et al. 2009, Schuster et al. 2015, Steinborn et al. 2011). Während einige Wirkungszusammenhänge bereits gut nachvollzogen werden können, gilt dies für andere vermutete Effekte weit weniger.
3.1 Auswirkungen auf Fledermäuse
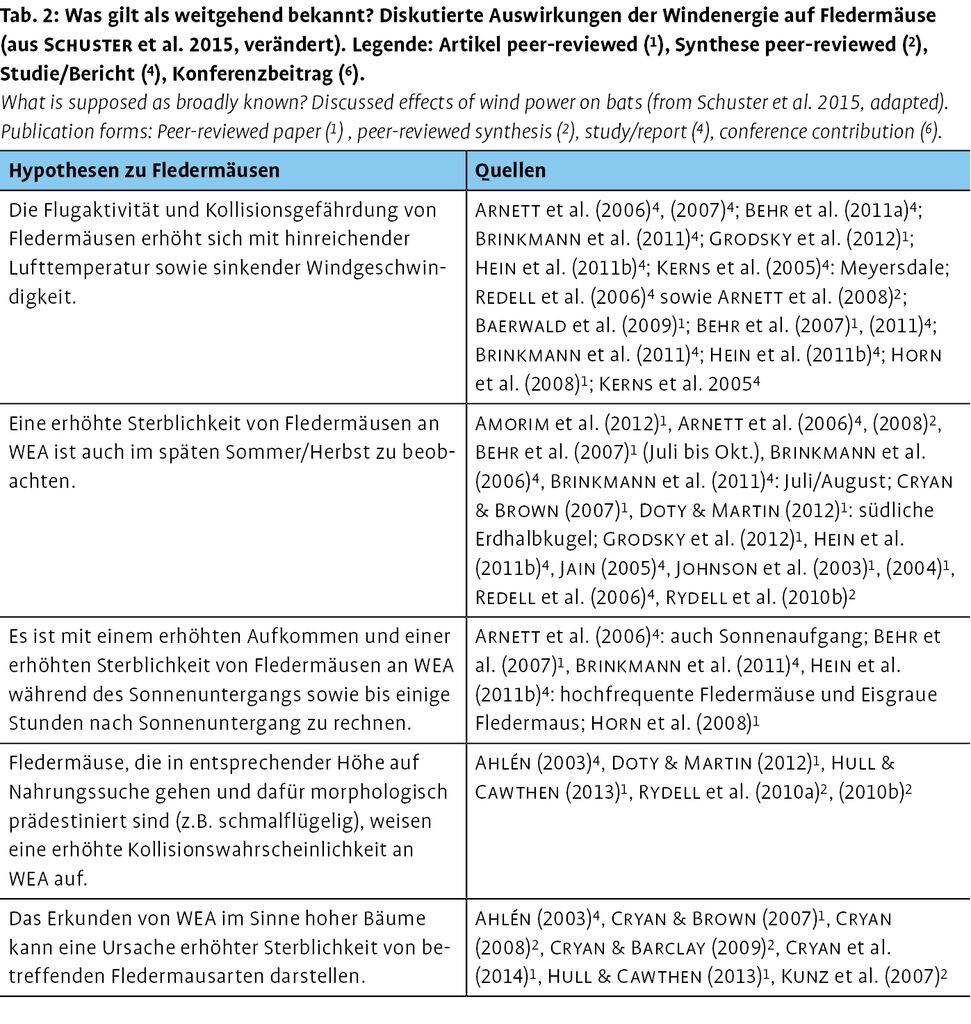
Die Synopse der Literaturanalyse spricht dafür, dass insbesondere die folgenden Hypothesen der Interaktionen zwischen Fledermäusen (Abb. 4) und WEA als plausibel gelten können (Tab. 2).
Enge Zusammenhänge zwischen der Aktivität von Fledermäusen und Windgeschwindigkeit, Niederschlag und Lufttemperatur wurden vielfach untersucht und diskutiert (z.B. Baerwald & Barclay 2011), so dass diese Parameter zunehmend in der Windenergieplanung zu einer Vorhersage eines erhöhten Kollisionsrisikos mit WEA und für die Konzeption und den Einsatz von Vermeidungsmaßnahmen während des Betriebs von WEA verwendet werden (z.B. Brinkmann et al. 2011). Beispielsweise sind in Deutschland in einer Folge von Forschungsvorhaben Algorithmen aus der Windgeschwindigkeit (Abnahme der Fledermausaktivität mit höherer Windgeschwindigkeit, z.B. ≥5ms1), dem Muster der nächtlichen Flugaktivität (Abb. 5) und der Temperatur (häufig nur geringe Flugaktivität unterhalb ca. 10°C) sowie Jahreszeit ermittelt worden, um zu einer Vorhersage erhöhter Fledermausaktivität zu gelangen und so anlagenspezifische Abschaltalgorithmen zu erlauben (vgl. Biehl et al. 2017). Ähnliche Studien belegen diesen Zusammenhang der Fledermausaktivität mit Lufttemperatur und Windgeschwindigkeit ebenso (z.B. Arnett et al. 2006, 2007; Behr et al. 2011a, Brinkmann et al. 2011, Grodsky et al. 2012, Hein et al. 2011b, Kerns et al. 2005, Redell et al. 2006).
Ferner wird eine erhöhte Sterblichkeit von Fledermäusen auch aufgrund des phänologischen Verhaltens determiniert. Dies ist damit zu erklären, dass während Migrationsperioden die Anzahl an verunglückten Individuen steigen kann (Cryan & Brown 2007). So zeigen Untersuchungen, dass während des späten Sommers und vorwiegend des Herbsts eine erhöhte Sterblichkeit von Fledermäusen besteht (Arnett et al. 2006, Doty & Martin 2012, Dürr 2002, Hull & Cawthen 2013). Beispielsweise stellte Endl et al. 2004 (in Rydell et al. 2010a) in einer Untersuchung fest, dass ca. 90 % der jährlichen Sterblichkeit von Fledermäusen im August und September und lediglich 10 % Anfang Juni zu dokumentieren waren.
Es wird angenommen, dass insbesondere das herbstliche Zugverhalten von Fledermäusen zu einer erhöhten Kollisionswahrscheinlichkeit führt. Dabei gilt zu beachten, dass Fledermäuse während der herbstlichen Migration über einen langen Zeitraum mit einer Vielzahl an Zwischenlandungen, etwa zur Nahrungssuche und Paarung, in Richtung ihrer Winterhabitate ziehen (Furmankiewicz & Kurcharska 2009). Dagegen wird das Kollisionsrisiko von Fledermäusen während der Frühlings-Migration insgesamt als geringer eingeschätzt, da Fledermäuse in dieser Zeit auf direktem Flugweg zu ihren Sommerlebensräumen und -schlafstellen gelangen. Dabei ist die Flughöhe tendenziell höher, so dass die Gefährdung durch die Windenergienutzung während dieser Jahreszeit offenbar sinkt (ebd.).
Ob migrierende Arten jedoch generell einem höheren Risiko ausgesetzt sind, ist noch nicht hinreichend geklärt (Schuster et al. 2015). Das erhöhte Aufkommen von Fledermäusen während des Sonnenuntergangs sowie bis einige Stunden nach Sonnenuntergang wiederum kann laut Horn et al. (2008) mit einem zu diesem Zeitpunkt erhöhten Insektenaufkommen erklärt werden.
Doch nicht nur die Phänologie, sondern auch die Morphologie von Fledermäusen sowie ihr betreffendes Verhalten haben nachweislich Einfluss auf das Kollisionsrisiko mit WEA. So haben Studien von Hull & Cawthen (2013) sowie Rydell et al. (2010b) aufgedeckt, dass kollidierte Fledermäuse unterschiedlicher Gattungen morphologische Ähnlichkeiten aufweisen. Fledermäuse, welche im freien Luftraum und somit i.d.R. in größeren Flughöhen mit Hilfe von hoch-intensiver Schmalband-Echoortung ihre Beute aufspüren sowie lange und schmale Flügel und entsprechende Flugeigenschaften aufweisen, zeigen in Europa (Nyctalus, Pipistrellus, Verspertilio, Eptesicus), Nordamerika (Lasiurus, Lasionycteris, Perimyotis) und Australien (Chalinolobus gouldii) eine höhere Kollisionswahrscheinlichkeit (Hull & Cawthen 2013, Rydell et al. 2010b). Tatsächlich gehen 98 % der an WEA getöteten Fledermäuse im freien Luftraum auf Jagd, während ca. 60 % der Arten lediglich in tiefen Flughöhen Nahrung suchen (Abb. 6) und damit einem geringeren Kollisionsrisiko in der durch den Rotor überstrichenen Fläche aufweisen (Rydell et al. 2010a).
Auffällig sind ebenso Untersuchungen von Cryan et al. (2014) und Kunz et al. (2007), welche zu dem Schluss kommen, dass womöglich von einer Verwechselung der Silhouette und Luftströmungen von WEA in der offenen Landschaft mit Bäumen ausgegangen werden kann. Fledermäuse, die Bäume als Schlafstellen nutzen, erkunden die WEA, so dass es zu einem erhöhten Kollisionsrisiko mit der Anlage kommen kann. Es wird vermutet, dass die Luftströmungen um stillstehende oder sich langsam drehende Rotorblätter einer WEA vergleichbar mit den Luftströmungen um große Bäume sein können (Cryan et al. 2014). Dies würde ebenfalls die Erkenntnis unterstützen, dass schnell drehende Rotorblätter von Fledermäusen weniger häufig angeflogen werden, da hierbei die Luftströmungen durch erratische, abwärts gerichtete Turbulenzen und somit ungünstige Jagdbedingungen gekennzeichnet sind.
Fledermäuse, welche sich in Höhlen anstatt in Bäumen zum Schlafen niederlassen, scheinen demgegenüber eine geringere Sterblichkeitsrate durch WEA aufzuweisen. Dies gilt ebenfalls vor dem Hintergrund, als dass Bennett & Hale (2014) für mit roter Befeuerung gekennzeichnete WEA eine geringere Sterblichkeit von Fledermäusen aufgrund einer womöglich besseren Unterscheidung zu hohen Bäumen feststellten. Eine Anlockung von Fledermäusen an befeuerten WEA infolge von Erkundungsflügen und Nahrungsvorkommen wie Insekten wird angenommen, allerdings sind die ursächlichen Gründe noch nicht gänzlich geklärt bzw. empirisch belegt (BFE 2015).
So verbleiben vielfach Unsicherheiten bezüglich der artbedingten Verhaltensmuster, aus denen erhöhte Kollisionswahrscheinlichkeiten mit WEA aufgrund von Aktivitätssteigerungen resultieren. So gehen beispielsweise Redell et al. (2006) und Arnett et al. (2006) von einer Abhängigkeit der Lufttemperatur hinsichtlich der Verwendung einer niedrigen oder höheren Frequenz der Echoortung aus. Fledermäuse, welche höhere Frequenzen zur Echoortung nutzen, zeigen nach Arnett et al. (2006) eine stärkere Aktivität als Fledermäuse mit einer Nutzung niedrigerer Frequenzen, während konträr nach Redell et al. (2006) die Fledermausaktivität mit einer Nutzung niedrigerer Frequenzen (zu 713 % ) ansteige und Fledermäuse mit hochfrequenter Echoortung lediglich zu 39 % aktiver werden (jeweils pro 1°C Lufttemperaturanstieg). So verbleibt z.B. für die Konzeption von Vermeidungsmaßnahmen unklar, ob tatsächlich eine erhöhte Kollisionswahrscheinlichkeit infolge erhöhter Aktivitäten für vorkommende Arten mit niedriger oder höherer Frequenz der Echoortung besteht.
Inwiefern vermehrt Fledermäuse unter hohen Luftdruckbedingungen, beispielsweise vor oder nach einer Sturmfront (Kerns et al. 2005), oder vielmehr unter niedrigen Luftdruckbedingungen fliegen (Baerwald & Barclay 2011, Cryan & Brown 2007), kann ebenfalls als nicht geklärt gelten (Schuster et al. 2015). Einige Studien gehen davon aus, dass bei Mondlicht die Fledermausaktivität steigt, während in einer anderen Untersuchungen ein geringes Mondlicht zusammen mit niedriger Windgeschwindigkeit sowie einer hohen Wolkenbedeckung zur Vorhersage einer erhöhten Fledermausaktivität beitragen konnte (Cryan & Brown 2007). Die Abhängigkeit von Klimaelementen ist weiterhin zu beobachten als auch artenspezifisch zu beurteilen (Baerwald & Barclay 2011, Behr et al. 2011a).
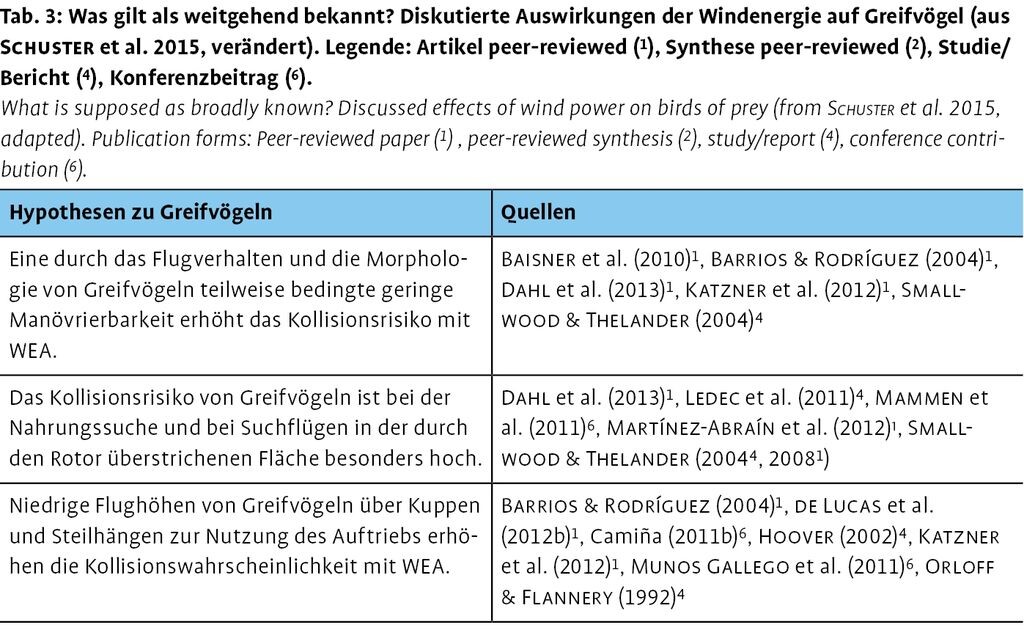
3.2 Auswirkungen auf Greifvögel
Vielfältige Untersuchungen zeigen, dass vornehmlich Greifvögel ein hohes Gefährdungspotenzial einer Kollision mit WEA aufweisen (vgl. z.B. Baisner et al. 2010, Dahl et al. 2013, Ledec et al. 2011, de Lucas et al. 2012b, Madders & Whitfield 2006, Martínez-Abraín et al. 2012, Smallwood & Thelander 2004, 2008; vgl. Tab. 3). So gingen z.B. Bellebaum et al. (2013) für Brandenburg von einem möglichen Rückgang von ca. 3 % des Rotmilan-Brutbestands (Milvus milvus) infolge der Windenergienutzung aus.
Insgesamt gilt zu beachten, dass Greifvögel ihre Größe und ihre Flügelspannweite zum Segeln und Gleiten (soaring flight) während mehr als 95 % ihrer Flugzeit nutzen. Der Vorteil dabei ist ein verringerter Energieverbrauch gegenüber dem Flügelschlag. Dies kann jedoch ihre Manövrierfähigkeit einschränken, so dass die Kollisionswahrscheinlichkeit mit WEA steigt (vgl. z.B. Baisner et al. 2010, Smallwood & Thelander 2004, Smallwood et al. 2009). Die Wetterbedingungen, vornehmlich das Wind- (als auch das Niederschlags-)aufkommen, üben also einen starken Einfluss auf das Flugverhalten von Greifvögel aus, da Luftströmungen für den Segelflug genutzt werden (Camiña 2011b, Hoover 2002, Katzner et al. 2012). Die wiederholenden Vorbeiflüge an WEA aufgrund kreisenden und segelnden Flugverhaltens führen zu einem erhöhten Kollisionsrisiko (Katzner et al. 2012, Ledec et al. 2011): Auf Suchflügen und der Jagd sowie im Zuge des Revierverhaltens können Greifvögel von der Präsenz der WEA abgelenkt werden (Abb. 7). So zeigten Untersuchungen, dass Greifvögel bei Suchflügen viel Zeit in der durch den Rotor überstrichenen Fläche verbringen (Smalllwood & Thelander 2004).
Das Kollisionsrisiko mit WEA kann als besonders hoch eingeschätzt werden, wenn Windparks in der Nähe von Kuppen oder Steilhängen errichtet werden. In Folge der Nutzung der orografischen Aufwinde sind Greifvögel zu einer niedrigeren Flughöhe gezwungen und folglich steigt die Wahrscheinlichkeit einer risikobehafteten Näherung an die Rotorblätter einer WEA (Camiña 2011b, Hoover 2002, Katzner et al. 2012). Mit dem Repowering von WEA muss nun auch bedacht werden, dass sich die Betroffenheit artspezifisch verändern kann. Mit den sodann höheren Anlagen, die auch bei Starkwinden länger im Wind bleiben können, sind entsprechende Flughöhen präferierende Arten fortan einem höheren Risiko ausgesetzt, während sich für andere Arten eine geringere Kollisionsgefährdung ergeben kann (Smallwood 2015). So stellten etwa Bergen et al. (2012) für den Rotmilan fest, dass im Zuge eines Repowering-Vorhabens in Nordrhein-Westfalen mit zunehmender Nabenhöhe die Aufenthaltsdauer im Rotorbereich und damit die Kollisionsgefahr sank, allerdings für den in größeren Höhen (über 90m) fliegenden Schwarzmilan (Milvus migrans) und dessen Jungvögel erhöhte Kollisionsrisiken entstanden. Insgesamt gingen Bergen et al. (ebd.) jedoch aufgrund ihrer modellhaften Untersuchungen davon aus, dass ein Repowering i.d.R. zu einer Reduktion der Kollisionsgefahr führt.
Bekannt sind Standortpräferenzen von Greifvögeln, sei es aufgrund der Topografie, welche mit Aufwinden den Segelflug bzw. lineare Flugkorridore begünstigt (Katzner et al. 2012, de Lucas et al. 2012b), oder aufgrund der Landnutzung, die Nahrungsressourcen bereitstellt, wie z.B. offene Agrarlandschaften mit einer geringen Vegetationsbedeckung (Bellebaum et al. 2013, Dürr 2009, Mammen et al. 2011). Vielfach wird davon ausgegangen, dass infolge eines vermehrten Greifvogelaufkommens eine erhöhte Kollisionswahrscheinlichkeit zu erwarten ist, sobald WEA an diesen Standorten errichtet werden (Barrios & Rodríguez 2004, Carrete et al. 2012, Martínez-Abraín et al. 2012, Smallwood et al. 2009). Doch andere Untersuchungen haben eine unmittelbare Korrelation zwischen dem Greifvogelaufkommen und der Sterblichkeit an WEA nicht bestätigen können (Garvin et al. 2011, de Lucas et al. 2008).
Auch die Frage, ob Greifvögel aktiv Standorte mit WEA meiden und damit auch Lebensräume verlieren, ist nicht geklärt (Schuster et al. 2015). Während einige Untersuchungen kein Meideverhalten in einem Windpark im Vergleich zu einer Referenzfläche aufzeigen (Bevanger et al. 2010 sowie Bergen et al. 2012 zu Rot- und Schwarzmilan, Dahl et al. 2013), stellten andere Studien fest, dass Greifvögel WEA visuell erkannten und sie folglich umfangreich mieden (Garvin et al. 2011, Hull & Muir 2013). Wieder andere Ergebnisse unterstreichen die Annahme, dass Greifvögel gar von WEA angezogen werden können (Smallwood & Thelander 2004). Jedoch betonen alle Autoren, dass die Verhaltensweisen immer artenspezifisch (Garvin et al. 2011) und in Abhängigkeit von der Umgebung (z.B. Camiña 2011b, Hoover 2002) um die Anlage sowie vom Anlagendesign und -laufzeiten beurteilt werden müssen (z.B. Hull & Muir 2013, Smallwood & Thelander 2004). Landschaftsveränderungen aufgrund der Anlagen und ihren Zuwegungen (Abb. 8) können ebenso Effekte hervorrufen wie etwa einen Populationsrückgang empfindlicher Arten wie des Schreiadlers (Aquila pomarina), der als eine Flaggschiff-Art für unzerschnittene Landschaften gilt (Langgemach et al. 2011).
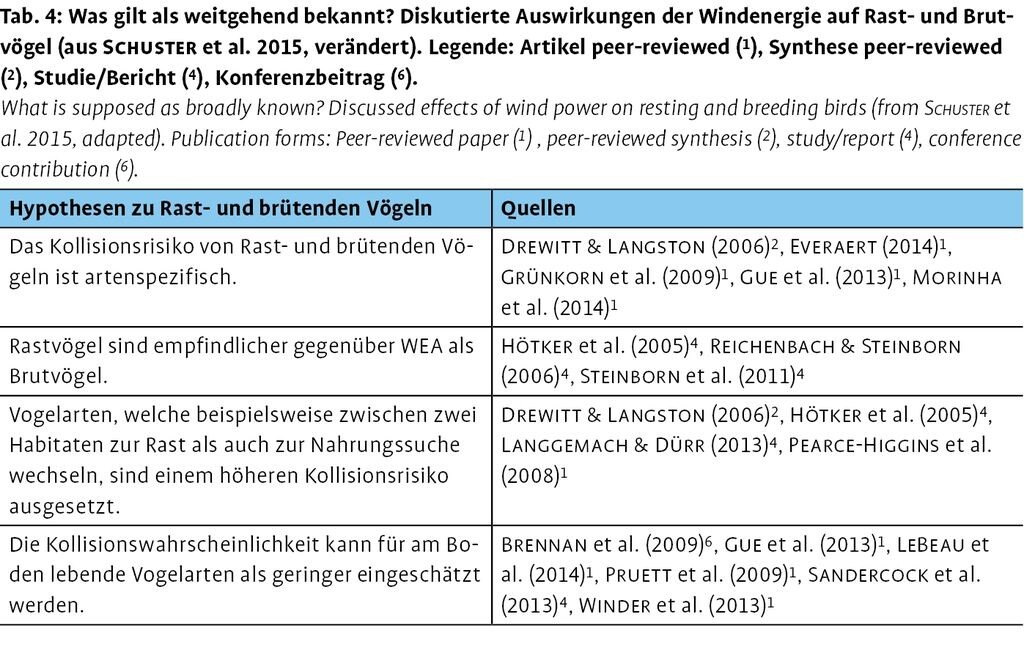
3.3 Auswirkungen auf Rast- und Brutvögel
Die Auswirkungen der Windenergienutzung auf Rast- und brütende Vögel werden im Folgenden beleuchtet (Tab. 4).
Von WEA ausgehende Gefährdungen für Vögel sind überwiegend von artspezifischen Verhaltensweisen und der jeweiligen Nutzung des Habitats abzuleiten, beispielsweise ob Vögel brüten oder rasten (Hötker et al. 2005, Reichenbach & Steinborn 2006). Sofern sich keine Brutstandorte im Einflussbereich von WEA befinden, gelangen Brutvögel weniger in den entsprechenden Risikobereich. Allerdings sind Rastvögel nicht unbedingt einem höheren Risiko ausgesetzt, da sie laut Steinborn et al. (2011) WEA erkennen und durch ein vermeidendes Verhalten alternative Habitate aufsuchen können. Dies hat allerdings zur Folge, dass eine Verdrängung auf weniger geeignete Habitate stattfinden kann (Rees 2012).
Belegt erscheint, dass Vogelarten und -individuen, die täglich zwischen mehreren Teilhabitaten wechseln, wie z.B. zur Nahrungssuche oder zur Rast, einem erhöhten Risiko ausgesetzt sind. Für diese Arten können WEA einen deutlichen Barriereeffekt ausüben oder das Kollisionsrisiko erhöht sich entsprechend (Hötker et al. 2005). Der Schwarzstorch (Ciconia nigra) z.B., welcher bis zu 20km zwischen seinen Nahrungs- und Bruthabitaten zurücklegt, kann durch Flugroutenveränderungen infolge der Barrierewirkung von WEA beeinträchtigt werden (Langgemach & Dürr 2013), auch möglicherweise durch einen höheren Energieverbrauch.
Eine geringe Kollisionswahrscheinlichkeit wiederum ist für am Boden lebende Vogelarten anzunehmen (Brennan et al. 2009, Gue et al. 2013, LeBeau et al. 2014, Pruett et al. 2009, Sandercock et al. 201, Winder et al. 2013). Beispielsweise zeigten Moorhühner eine geringere Kollisionswahrscheinlichkeit, da sie nicht in den Einflussbereich der Rotorblätter gelangten (Brennan et al. 2009, Pruett et al. 2009). Das Flugverhalten spielt somit eine große Rolle in Hinblick auf ein Gefährdungspotenzial; andererseits kann mit der geringeren Flughöhe von Vogelarten die Kollisionswahrscheinlichkeit mit dem Turm einer WEA steigen. Beispielsweise konnte dies für das Schottische Moorschneehuhn (Lagopus lagopus scoticus) in Schweden sowie für das Birkhuhn (Tetrao tetrix) in Österreich gezeigt werden (Falkdalen et al. 2013, Zeiler & Gruenschachner-Berger 2009).
Dennoch bestehen weiterhin vielfältige Unsicherheiten. Während einige Untersuchungen beispielsweise eine Verdrängung von Brachvögeln (Numenius arquata) und ihren Brutstandorten während des Baus eines Windparks hin zu einer Referenzfläche nachwiesen (z.B. Pearce-Higgins et al. 2012), zeigten andere Artengruppen abweichende Verhaltensweisen. So war in der gleichen Untersuchung von Pearce-Higgins et al. (2012) eine erhöhte Dichte von anderen Vogelarten auf dem Standort des neuen Windparks festzustellen. Pearce-Higgins et al. (2012) erläuterten, dass aufgrund der gestörten Vegetationsbedeckung infolge des Baus der WEA verbesserte Habitatbedingungen z.B. für die Feldlerche (Alauda arvensis), das Schwarzkehlchen (Saxicola rubicola) und den Wiesenpieper (Anthis pratensis) geschaffen wurden. Andererseits stellte eine andere Untersuchung ein Meideverhalten des Wiesenpiepers bis 100m Entfernung zu der Baufläche von WEA fest (Steinborn et al. 2011). Nicht nur die Wirkungszusammenhänge zwischen dem Bau von WEA und der Vogelwelt sind somit in den Blick zu nehmen, sondern auch die Auswirkungen von Landnutzungsänderungen im Fokus zu behalten. Ebenfalls die Erschließung zu WEA kann Lebensräume verändern und Barrierewirkungen zwischen Teillebensräumen entfalten (LAG VSW 2014).
Auch beim Betrieb von Windparks stellen einige Untersuchungen ein vermeidendes Verhalten von Vogelarten zu den WEA und den Zugangswegen fest (z.B. Leddy et al. 1999, Pearce-Higgins et al. 2009). Bei einem Vergleich von zwölf Windparks wurde erkenntlich, dass sieben von zwölf Arten, wie der Goldregenpfeifer (Pluvialis apricaria), der Steinschmätzer (Oenanthe oenanthe) und die Bekassine (Gallinago gallinago), mit einem Abstand von 100 bis 800m WEA mieden, ebenso war ein Brutrückgang von 50 % festzustellen (Pearce-Higgins et al. 2009). Demgegenüber konnten Studien von Douglas et al. (2011) und Steinborn & Reichenbach (2012) kein Meideverhalten des Goldregenpfeifers sowie keine Auswirkungen auf die Vogelpopulationen (insbesondere für Sumpf- und Sperlingsvögel) und den Bruterfolg durch die Windenergienutzung feststellen (vgl. de Lucas et al. 2005). Um vorsorglich einem womöglich mit erhöhtem Kollisionsrisiko einhergehenden Gewöhnungsverhalten des Goldregenpfeifers entgegenzuwirken, wird andererseits vorgeschlagen, einen Mindestabstand von 1000m bzw. einen Prüfradius von 6000m bei der Planung von WEA einzuhalten (LAG VSW 2014).
Die Diskrepanzen zwischen den Studienergebnissen machen deutlich, dass hier vielfach Unklarheiten verbleiben, welche auf das jeweilige Studiendesign, die Standortcharakteristika oder die jährlichen Dynamiken und das artspezifische Verhaltens zurückgeführt werden könnten (Schuster et al. 2015). Beispielsweise lassen Untersuchungen von Fijn et al. (2012) erkennen, dass zwar der Zwergschwan ein Meideverhalten gegenüber WEA zeigt, doch während des Jahres mit einer wachsenden Nahrungsressource in Nähe der WEA näher an diese heranrückte.
Inwieweit die jeweilige Landnutzung stärkere Auswirkungen auf die Größe des Brutbestands als der Abstand zu WEA ausüben kann (Steinborn et al. 2011), wird ebenfalls diskutiert. Kollisionen am Anlagenturm wurden vermehrt beobachtet, wenn der Turm eine weiße oder graue Farbe besitzt. Im Vergleich dazu zeigten WEA mit einem grünen Fundament und einem Farbverlauf in hellere Farbtöne mit zunehmender Höhe der Anlage keine Sterblichkeit von Arten (Dürr 2011). Dennoch wird in Untersuchungen eine artenspezifische Reaktion auf Farbe, Typ und Intensität der Befeuerung einer WEA (Blew et al. 2013a, van de Laar 2007) betont und sollte daher weiter untersucht und diskutiert werden (Blackwell & Fernádez-Jurici 2013).
3.4 Auswirkungen auf Zugvögel
Schon eine geringe Anzahl von Schlagopfern bei adulten Seevögeln während der Zugperioden kann nach Stienen et al. (2007) eine Auswirkung auf die Populationen bedeuten. Zugvögel können insbesondere auch durch die Offshore-Windenergie betroffen sein. Diskutiert werden Barriereeffekte, Störungen (z.B. Lärmimmissionen, Reflektionen) oder Habitatverluste (z.B. Hüppop et al. 2006; Tab. 5).
Zugvögel nutzen u.a. bestimmte Landschaftsstrukturen und -elemente zur Orientierung. Wenn Zugvögel über See fliegen, erhöht sich die Migrationsintensität in der Nähe von Küsten, da die meisten Wildvögel diese als Korridore nutzen; wichtige Zugkorridore sind dabei beispielsweise die Nord- und Ostsee (Hüppop et al. 2006, Stienen et al. 2007), die Straße von Gibraltar (Muños Gallego et al. 2011) sowie Flugwege entlang der atlantischen Küstenlinie der USA (Burger et al. 2011). Die Kollisionswahrscheinlichkeit mit WEA erhöht sich dabei mit der Zugvogelkonzentration und infolge des wiederholten Vorbeifliegens an den Anlagen bei der Rückmigration (Hüppop et al. 2006, 2012). Zwar findet Vogelzug ganzjährig statt, doch ist das erhöhte Aufkommen von Zugvögeln im Frühling und Herbst v.a. für das Kollisionsrisiko relevant (Hill et al. 2014, Hüppop et al. 2006). Dies gilt auch insofern, als dass nur wenige Massen-Migrationsperioden von Zugvögeln im Jahr auftreten (Aumüller et al. 2011, Hill et al. 2014, Hüppop et al. 2006). Derart kann die Wahrscheinlichkeit eines erhöhten Kollisionsrisikos auf diese Zeitpunkte fokussiert und bei der Planung von Vermeidungsmaßnahmen berücksichtigt werden.
Viele Untersuchungen stellen heraus, dass schlechte Wetterbedingungen bzw. schnelle Wetterwechsel zu einer niedrigeren Flughöhe und damit zu einer erhöhten Kollisionswahrscheinlichkeit in der durch den Rotor überstrichenen Fläche führen (z.B. Aumüller et al. 2011, Coppack et al. 2011, Hüppop et al. 2006). Folglich können auch schlechte Wetterbedingungen an den Abflugstandorten der Zugvögel zu einem Ausfall der Zugbewegungen führen (Richardson 2000 in Hein et al. 2011a). Auch eine Interaktion zwischen der Wolkenbedeckung, den Windbedingungen, der Lufttemperatur sowie dem Luftdruck, dem Niederschlag und der Zugaktivität von Wildvögeln wird in verschiedenen Studien nachgewiesen (Aschwanden et al. 2011: Onshore, Nilsson et al. 2006; Aumüller et al. 2011, Hüppop et al. 2004, 2006, van der Laar 2007, Hüppop & Winkel 2006, Hein et al. 2011a, Shamoun-Baranes et al. 2006). Gute Wetterbedingungen für einen risikoverminderten Vogelzug würden demnach durch eine gute Sichtbarkeit und Rückenwind charakterisiert werden (Coppack et al. 2011, Hüppop et al. 2006, Plonczkier & Simms 2012).
Neben den Wetterbedingungen übt die Befeuerung von WEA Einfluss auf das Zugverhalten von Wildvögeln aus. Insbesondere nächtlich migrierende Vögel, wie Singvögel oder Sumpfvögel, können durch die Beleuchtung der Anlagen (wie auch von Ölplattformen) angezogen werden (Aumüller et al. 2011, Hill et al. 2014, van der Laar 2007). Vor allem bei geringer Sichtbarkeit wie bei Nebel oder Nieselregen wurde festgestellt, dass die beobachteten Vögel um die Beleuchtungsquellen kreisten, die Anlagen als Ruheplätze aufsuchten und deutlich ein desorientiertes Verhalten anzeigten. Aumüller et al. (2011) verdeutlichen in einer Untersuchung, dass dies nicht allein zu einer erhöhten Kollisionswahrscheinlichkeit der Zugvögel führen kann, sondern auch zu einer gesteigerten Sterblichkeitsrate aufgrund von Erschöpfung oder Mangel an Nahrungsquellen (Blew et al. 2013b, Hüppop et al. 2006, van der Laar 2007).
Weitere Wirkungszusammenhänge zwischen (Offshore-)WEA und Zugvögeln werden noch diskutiert. Beispielsweise weisen einige Studien nach, dass ein Großteil der Zugvögel auf dem Weg von ihrem Sommer- zu ihrem Winterhabitat (und umgekehrt) vorwiegend in der Nacht aufgrund einer großen Zughöhe WEA meiden (Christensen et al. 2004, Hill et al. 2014, Hüppop et al. 2012). Abweichend wurde für die Kurzschnabelgans (Anser brachyrhynchus) überwiegend in der Mittagszeit zwischen 12.00 Uhr und 14.00 Uhr Zugverhalten aufgezeigt und lediglich zu 15 % ein nächtliches zwischen 19.00 Uhr und 07.00 Uhr morgens (Plonczkier & Simms 2012). Hüppop et al. (2012) beobachteten bei Untersuchungen an der Forschungsplattform FINO 1 in der deutschen ausschließlichen Wirtschaftszone (AWZ) von 2004 bis 2007 eine erhöhte nächtliche Flugaktivität kurz vor Sonnenaufgang bei 80 % der Zugvögel. In gleicher Weise betont Hill et al. (2014) ein erhöhtes Zugaufkommen nach Mitternacht insbesondere innerhalb der Migrationsperioden im Herbst und Frühling.
Während Untersuchungen von Reichenbach & Grünkorn (2011) Hinweise auf ein aktives Meideverhalten von Zugvögeln in der Nacht durch eine angepasste Flughöhe erbrachten (ebenso Singvögel bei Hill et al. 2014 bzw. Pettersson & Fågelvind 2011), zeigen andere Studien dieses nicht. Ein präziseres Vermeidungsverhalten wurde dabei auch am Tage festgestellt; Christensen et al. (2004) untersuchten den Offshore-Windpark Horns Rev in Dänemark, um artenspezifische Verhaltens- und Bewegungsmuster von vorbeiziehenden Vögeln an dem zu diesem Zeitpunkt kürzlich errichteten Windpark zu bestimmen. Am Tage konnten die WEA von Zugvögeln besser erkannt und umflogen werden.
3.5 Auswirkungen auf Meeressäuger
Meeressäugetiere wie Seehunde und Wale besitzen eine hohe akustische Sensibilität, da ihre Kommunikation, Navigation und Lokalisierung von Nahrungsressourcen i.d.R. auf akustischen Signalen zur Echoortung beruht (Kastelein et al. 2013, Lucke et al. 2009; Abb. 9). Insbesondere die sogenannten inshore-Arten, wie der Schweinswal (Phocoena phocoena) und der Große Tümmler (Tursiops truncatus), können durch die Windenergienutzung im Bereich von geringeren Wassertiefen betroffen sein (Gordon et al. 2007). Die Auswirkungen lassen sich nach Stand des Wissens jedoch überwiegend auf den Bau der Anlagen zurückführen: So üben die betriebsbedingten Lärmimmissionen laut z.B. Gordon et al. (2007), Madsen et al. (2006), Teilmann et al. (2012), Tougaard et al. (2006a) und Tougaard (2013) vernachlässigbare Effekte auf die marine Meereswelt aus. Infolge eines ggf. forcierten Offshore-Ausbaus dürfen allerdings künftig betriebsbedingte Lärmimmissionen von WEA nicht von vornherein vernachlässigt werden, so bei der Betrachtung kumulierender Lärmimmissionen, wie etwa aufgrund der Schifffahrt bzw. Tätigkeiten zur Unterhaltung der Offshore-Windparks (wie z.B. Wartung) (Madsen et al. 2006). Infolgedessen können überwiegend die in Tab. 6 zusammengefassten Erkenntnisse hinsichtlich der Auswirkungen der Offshore-Windenergie als plausibel eingestuft werden:
Bei dem Bau von Offshore-WEA können neben dem bislang vorherrschenden direkten Rammen (Pile-Driving) des Anlagenturms in den Meeresgrund (Abb. 10) Baggerarbeiten und Bohrungen zum Einsatz kommen (Schuster et al. 2015). Dies ist von der Anlagenkonstruktion bzw. dem Fundament und den jeweiligen Standortbedingungen, der Wassertiefe und Wirtschaftlichkeit abhängig, z.B. ob Einpfahl- (Monopiles) oder auch Mehrpfahlstrukturen bzw. Bucket-Fundamente oder andere Bauweisen zum Einsatz kommen. Die meisten Auswirkungen sind von der Impulsrammung von Monopiles bekannt (z.B. Bailey et al. 2010, Danish Energy Agency 2013, Gerdes & Ludwig 2014, Gordon et al. 2007, Gordon 2012). Die jeweilige Ausbreitung der Lärmimmissionen ist dabei von verschiedenen Faktoren wie dem Salzgehalt, der Wassertiefe und -temperatur abhängig und wird desgleichen durch die Wetterbedingungen beeinflusst (Gerdes & Ludwig 2014). Eindeutig belegt ist, dass durch die Impulsrammung marine Säuger wie der Schweinswal von den Anlagenstandorten verdrängt werden (z.B. Brandt et al. 2011, Dähne et al. 2013, Lucke et al. 2012, Tougaard et al. 2012).
Gleichwohl lassen Untersuchungen vermuten, dass diese Auswirkungen lediglich zeitlich begrenzt und damit reversibel sein können (z.B. BMUB 2013, Danish Energy Agency 2013). Dennoch zeigen Beispiele, dass die Wiederansiedlung um die WEA bis zu zwei Jahre andauern kann (Tougaard 2006a, b), während in einer Studie von Diedrichs et al. (2008) erst vier Jahre nach dem Bau eines Offshore-Windparks keine Unterschiede der Schweinswalpopulationen im Vergleich zu der Populationsstärke vor dem Bau auftraten. Die jeweilige Signifikanz der Verdrängung sollte dabei immer vor dem Hintergrund von weiteren Faktoren beurteilt werden, wie z.B. der umliegenden Habitatqualität, Beuteverfügbarkeit, Schifffahrtsaktivität sowie Konkurrenzverhalten (Scheidat 2012).
Die genannten Risiken wie ein direkter Hörverlust infolge der Impulsrammung entstehen i.d.R. in der direkten Umgebung der jeweiligen Bauarbeiten (Brandt et al. 2009; Tougaard et al. 2009, 2012). Dabei gilt zu beachten, dass der emittierte Lärm nicht nur in den Wasserkörper, sondern auch in den Meeresgrund abgegeben wird und so in einer Distanz von 20 bis 50m je nach Sediment wieder an den Wasserkörper weitergegeben werden kann (Lippert et al. 2014). Beispielsweise deckten Untersuchungen von Bailey et al. (2010) auf, dass Populationen des Großen Tümmlers (Tursiops truncatus) in einem Umkreis von 100m um die Rammarbeiten einer WEA in Gefahr sind, starke Verletzungen des Hörsystems zu erleiden. Verhaltensänderungen sind bereits in einem Umkreis von 50m um den Anlagenbau zu beobachten (Bailey et al. 2010). Verletzungen der Hörsysteme, wie eine permanente Schwerhörigkeit (permanente Hörschwellenverschiebung) der Meeressäugetiere können laut Nedwell et al. (2007) bereits ab einer Exposition von 130dBht für wenige Sekunden auftreten.
Auch das mögliche Unvermögen, wichtige Signale im Hintergrundschall zu erkennen, die sogenannte Maskierung innerartlicher Kommunikation durch die Schallereignisse beim Rammen, kann zu erheblichen Konsequenzen führen; Meeressäugetiere sind auf den Empfang von akustischen Signalen für Nahrungssuche, Orientierung, Kommunikation, Paarung usw. angewiesen (Kastelein 2013). Folglich sind langfristige, v.a. auch kumulative Auswirkungen auf die Meeresumwelt nicht auszuschließen, allerdings auch noch schwierig zu beurteilen (Dähne et al. 2014).
Die Sorge um sich kumulierende Lärmimmissionen aufgrund von sich ein- bis zweimal pro Sekunde wiederholenden Rammstößen sowie bezüglich simultaner und dauerhafter Bauaktivitäten an verschiedenen Standorten oder innerhalb eines Windparks wird daher vielfach diskutiert (z.B. Gordon et al. 2009 in Tougaard et al. 2012, Lucke et al. 2009). Beispielsweise sind für die Installation eines Tripod- oder Jacket-Fundamentes einer Offshore-WEA im Mittel ca. 15000 Rammstöße nötig, welche einen jeweiligen Wert von 230dB re 1μPA bei 1m erreichen (Matuschek & Betke 2009). Eine langandauernde Belastung durch Lärmimmissionen auch mit einem geringen Wert von 90dB würde zwar einen permanenten Hörverlust von Meeressäugetieren ausschließen, aber aufgrund der Belastungsdauer (z.B. bei einer Arbeitszeit von acht Stunden pro Tag) das Hörvermögen dennoch beeinträchtigen (Nedwell et al. 2007). Dies gilt insbesondere, wenn die Intervalle der Rammarbeiten kürzer als die nötige Erholungszeit für einen temporären Hörverlust ausfallen, so dass sich die auditiven Effekte kumulieren (Lucke et al. 2009).
Betriebsbedingte Auswirkungen von Offshore-WEA bedeuten auch neuartige Unterwasserstrukturen für die marine Fauna. Zwar sind Offshore-WEA oft auf Sandbänken lokalisiert, welche ein wichtiges Habitat während des Kalbens und Säugens für Meeressäugetiere darstellen (Koschinski 2002 in Carstensen et al. 2006), doch wird aufgrund des Einbringens von harten Substraten durch den Turmfuß sowie dessen Kolkschutz die strukturelle Heterogenität des Meeresgrundes vergrößert. In dieser Weise werden künstliche Riffe ausgebildet, welche zu einer neuen Habitatqualität nicht nur für Meeressäugetiere, sondern auch für marine Beutetiere führen können (Gordon 2012, Lindeboom 2012, Scheidat et al. 2011, Thompson et al. 2013, Winter et al. 2012). Beispielsweise zeigen Untersuchungen von Russel et al. (2014), dass Seehunde zwei Offshore-Windparks in Deutschland und England (alpha ventus und Sheringham Shoal) offenbar zur Nahrungssuche gezielt aufsuchten (nach vier bzw. zwei Jahren des Baus der WEA).
In welchem Maße letztlich die betriebsbedingten Effekte auf die Meeresumwelt insgesamt einzuschätzen sind, wird nicht einheitlich diskutiert. Neben der Annahme, dass die betriebsbedingten Lärmimmissionen unter Wasser vernachlässigbar seien (z.B. Gordon et al. 2007, Madsen et al. 2006, Teilmann et al. 2012), betonen andere Untersuchungen die Auswirkungen von kumulativen Effekten infolge des forcierten Offshore-Ausbaus (Koschinski et al. 2003, Tougaard et al. 2009). Eine Maskierung (Schallüberdeckung) während des Betriebs der WEA, welche eine eingeschränkte Kommunikation von Schweinswalen aufgrund von anderen Lärmimmissionen zur Folge haben kann, werden bei dem Betrieb von Windparks teilweise als nicht signifikant eingestuft; lediglich eine geringe spektrale Überlappung mit den akustischen Signalen der Echoortung von Schweinswalen sei zu beobachten (Gordon 2012, Tougaard et al. 2009). Untersuchungen von Lucke et al. (2008) betonen dagegen, dass eine auditive Maskierung bei kleineren Anlagen möglich ist, allerdings nur in geringen Abständen zu der WEA sowie in Abhängigkeit von den Wetterbedingungen. So würden starke Winde hemmende Auswirkungen auf die Kommunikation der Meeressäugetiere wahrscheinlicher machen und auch Ødegaard & Daneskiold-Samsøe A/S 2000 (in Simmonds & Brown 2010) machen deutlich, dass die betriebsbedingten Lärmimmissionen nicht nur von der Art des Fundaments der WEA abhängig sind, sondern ferner von der Schallausbreitung im Meer.
Eine weitere Untersuchung hält die betriebsbedingten Auswirkungen der auditiven Maskierung für Seehunde (Phoca vitulina) für wahrscheinlicher als für den Schweinswal (Koschinski et al. 2003). Beide Arten können die von Offshore-WEA emittierten niedrigen Frequenzen nachweisbar wahrnehmen, allerdings besitzen Seehunde ein besseres Hörvermögen von Tieftonfrequenzen (100 bis 1000Hz) und nutzen dies für die intraspezifische Kommunikation (Richardson et al. 1995 in Koschinski et al. 2003), im Vergleich zu Schweinswalen (Koschinski et al. 2003).
4 Zwischenfazit und Ausblick
Zahlreiche Untersuchungen in den letzten Jahren belegen einen intensiven internationalen Austausch und eine umfassende Analyse der Wirkungszusammenhänge zwischen der Windenergienutzung und den relevanten Tierartengruppen. Zwar konnten einige Wirkungszusammenhänge bereits gut nachvollzogen und damit für die Windenergieplanung hilfreich genutzt werden (z.B. zur Entwicklung von Abschaltalgorithmen in Zeiten hoher Fledermausaktivitäten), doch verbleiben gleichwohl Unsicherheiten.
Die Gegenüberstellung der Hypothesen zu eher bekannten Effekten und den mit größeren Unsicherheiten gekennzeichneten Ergebnissen macht deutlich, dass bei allen Artengruppen die artenspezifischen Verhaltensmuster sowie die äußeren Umweltbedingungen (wie z.B. die jeweilige Landnutzung) divergierende Interaktionen mit WEA hervorrufen können. Dennoch können für bestimmte Faktoren offensichtliche Verhaltensmuster aufgezeigt und z.B. Zeiten höherer Kollisionsrisiken prognostiziert werden, wie etwa die Korrelation der Kollisionsgefährdung von Fledermäusen mit den vorherrschenden Windgeschwindigkeiten. Allerdings umfasst die Auswahl der untersuchten Wildtierarten oftmals besonders planungs- und genehmigungsrelevante Arten, wie auch diese synoptische Literaturauswertung zeigt (vgl. Boldt & Hummel 2013), so dass mögliche Kenntnislücken zu Auswirkungen auf zunächst als weniger relevant eingeschätzte Arten auftreten können.
Weiterhin wurde deutlich, dass einige Effekte in Studien bereits zahlreich untersucht wurden, beispielsweise der Einfluss von Standortfaktoren auf die Raumnutzung von Greifvögeln (z.B. der Einfluss von Hängen und Kämmen auf den Segelflug und entsprechende Kollisionsrisiken), während einige Themen nur von wenigen Studien behandelt werden, etwa die offenen Fragen der Gewöhnung von Zugvögeln an Windparks (vgl. Plonczkier & Simms 2012). Inwiefern sich Arten womöglich nicht nur an Vermeidungsmaßnahmen (Gartman et al. 2016), sondern eventuell auch an Windparks und mit welchen möglichen Folgen gewöhnen könnten, bleibt eine ebenso offene Frage. Um ggf. Erkenntnisse über die Art und Weise einer Lebensraumanpassung zu erlangen, sind methodische Langzeitüberwachungen nötig und auch die Analyse von Verdrängungsstudien kann als hilfreich erachtet werden, um langfristige Veränderungen in der Folge der Errichtung von Windparks zu erfassen (vgl. May 2015).
Vorher-Nachher-Untersuchungen (sowohl vor als auch nach dem Bau von WEA) sollten in diesem Zuge vermehrt Erkenntnisse bringen (May et al., angen.). Funktionsraumanalysen können zum Einsatz kommen, um von Arten aktiv genutzte Flächen für Planungen frei zu halten (wie z.B. für den Schreiadler, Langgemach et al. 2013). Gleichzeitig können nicht hinreichende Funktionsraumanalysen zu verfälschten Ergebnissen und Schlussfolgerungen für Standorte von WEA führen (ebd.).
Außerdem erfordern Genehmigungsverfahren ein größeres Wissen darüber, ab wann sich zusätzliche Mortalitäten aufgrund der weiteren Entwicklung der Windenergie als populationsrelevant erweisen können (Bellebaum et al. 2013, Busch & Garthe 2016, Diffendorfer et al. 2015, Lonergan 2011). So können kumulative Effekte im Gesamtlebensraum einer Art zu einer Verschlechterung des Erhaltungszustands einer Population führen (LAG VSW 2014). Auch sind bislang die Auswirkungen der Windenergie auf wandernde Arten häufig nicht hinreichend beschrieben. Weitere Populationsmodellierungen von kollisionsgefährdeten Vogel- und Fledermausarten sind sicherlich hilfreich (vgl. Bellebaum et al. 2013, Busch & Garte 2016). Gleichzeitig ist festzustellen, dass auch bei Kollisionsrisikomodellen (Masden & Cook 2016) noch ein vielleicht weiterer Weg vor uns liegt als bereits angenommen, wie sich etwa in jüngerer Zeit bei der Plausibilisierung von Eingangs-Annahmen in das schottische Band-Modell zeigte (Reichenbach 2015).
Neben einem weiteren intensiven internationalen Wissensaustausch bedarf es auch einer besseren Datenverfügbarkeit, um einen nachhaltigen Ausbau der Windenergie zu ermöglichen. Folglich ist auch an die Praxis zu appellieren, ihren Beitrag für einen verbesserten Umgang mit Unsicherheiten zu leisten und etwa die zahlreichen Monitoring-Daten auf geeignete Art und Weise für mögliche Meta-Analysen zur Verfügung zu stellen (vgl. Voigt et al. 2015). Abschließend gilt festzuhalten, dass Unsicherheiten keineswegs jemals weitgehend ausgeräumt werden können (Köppel et al. 2014b).
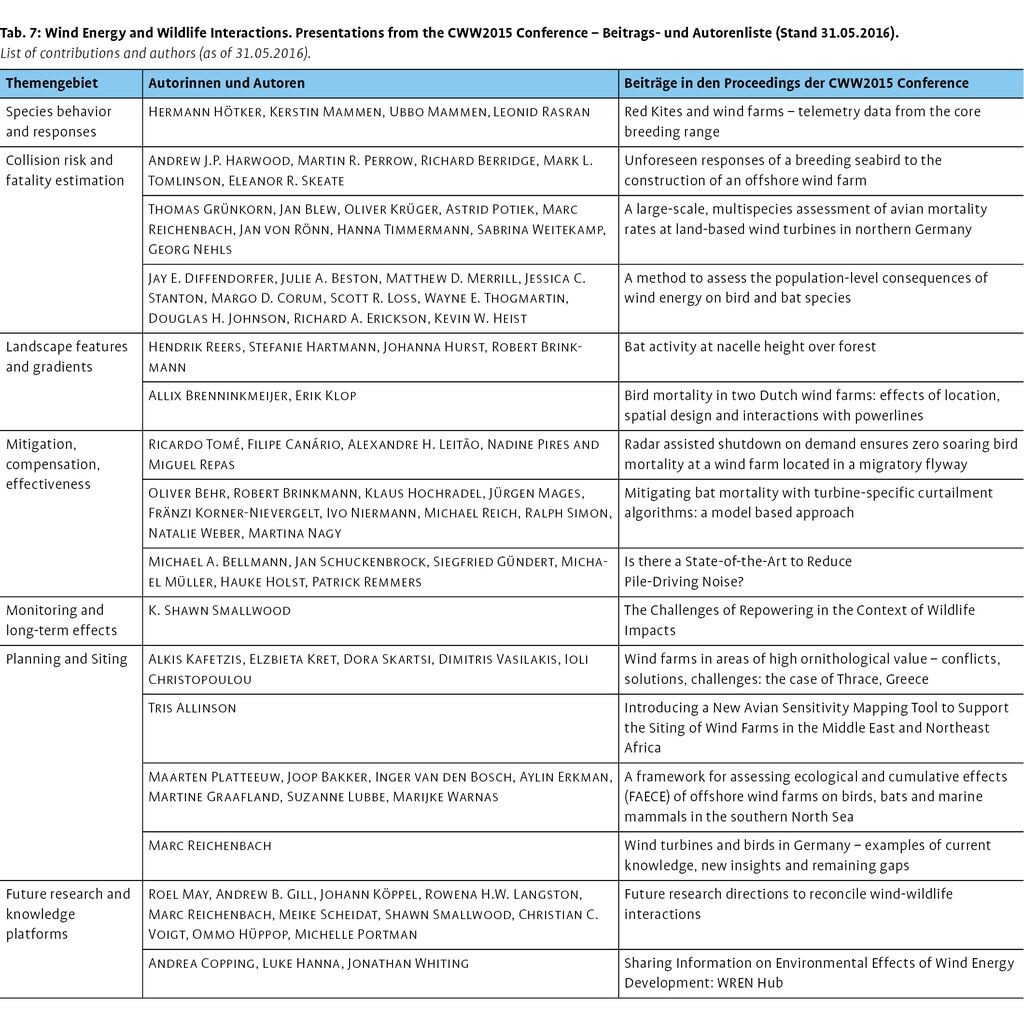
Inwiefern den hier aufgezeigten Wirkungsbeziehungen zwischen der Windenergie und der Tierwelt mit Hilfe von Vermeidungsmaßnahmen begegnet werden kann und wie weit der Stand der Forschung hierzu ist, wird in dem Beitrag von Biehl et al. (2017, in diesem Heft) näher beleuchtet. Eine aktuelle Buchpublikation zu der internationalen Conference on Wind energy and Wildlife impacts (CWW2015) in Berlin mit ausgewählten Konferenzbeiträgen befindet sich in Vorbereitung, in welcher auch eine Reihe deutscher Autorenteams Beiträge aus Forschung und Praxis vorstellen (Tab. 7).
Literatur
siehe beitragsübergreifende Bibliographie unter http://www.nul-online.de, Webcode 2231
-
Abb. 1: Kein seltenes Bild in Norddeutschland ziehende Kraniche (Grus grus) vor Windkraftanlagen.
Not a rare picture in Northern Germany migrating cranes (Grus grus) in front of wind power plantsARSU GmbH -
Abb. 2: Totfund einer Heringsmöwe (Larus fuscus).
Lesser black-backed gull (Larus fuscus) found dead.ARSU GmbH -
Abb. 3: Totfund eines Sommergoldhähnchens (Regulus ignicapilla).
Common firecrest (Regulus ignicapilla) found dead.ARSU GmbH -
Abb. 4: Abendsegler (Nyctalus noctula) zählen zu den windkraftsensitiven Fledermäusen
Common noctule bats (Nyctalus noctula) are particularly sensitive to wind power plants. Freiburger Institut für angewandte Wildtierökologie GmbH -
Abb. 5: Nächtliche Lichtspur beim Netzfang von Fledermäusen.
Nocturnal track of light during mist netting of bats.Freiburger Institut für angewandte Wildtierökologie GmbH -
Abb. 6: Netzfänge von Fledermäusen ergänzen die Erfassung mit Bat-Detektoren und erlauben eine artspezifische Ansprache.
Mist netting of bats complements the recording with bat detectors and allows the secure identification of the species.Ed Arnett -
Abb. 7: Die Erfassung von Kollisionsopfern (hier Mäusebussard, Buteo buteo) erfordert ein hohes Maß an Standardisierung und bedarf statistischer Verfahren zur Abschätzung nicht erfasster Totfunde.
The recording of collision casualties (here common buzzard Buteo buteo) requires a large amount of standardisation and involves statistical procedures to estimate the amount of dead individuals which are not found.ARSU GmbH -
Abb. 8: Die Auswirkungen der Erschließung der Windparks werden ebenfalls diskutiert.
The effects of the infrastructure of wind farms are also discussed.Ed Arnett -
Abb. 9: Ausbringung von Hydrophonen zur Erfassung von Schweinswalen
The application of hydrophones to record harbour porpoises.
Abb. 10: Rammarbeiten zur Gründung von Offshore WEA.
Pile driving for the foundation of offshore wind farms.Thilo Liesenjohann; Andreas Buchheim -
Auswirkungen der Windenergie auf Tierarten -
Auswirkungen der Windenergie auf Tierarten -
Auswirkungen der Windenergie auf Tierarten -
Auswirkungen der Windenergie auf Tierarten -
Auswirkungen der Windenergie auf Tierarten -
Auswirkungen der Windenergie auf Tierarten -
Auswirkungen der Windenergie auf Tierarten -
Auswirkungen der Windenergie auf Tierarten













Barrierefreiheits-Menü
Schriftgröße
Kontrast
Menü sichtbar
Einstellungen
Zu diesem Artikel liegen noch keine Kommentare vor.
Artikel kommentierenSchreiben Sie den ersten Kommentar.